Quantum Modeling: A Bridge between the Pumping and Signaling Functions of Na/K-ATPase

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
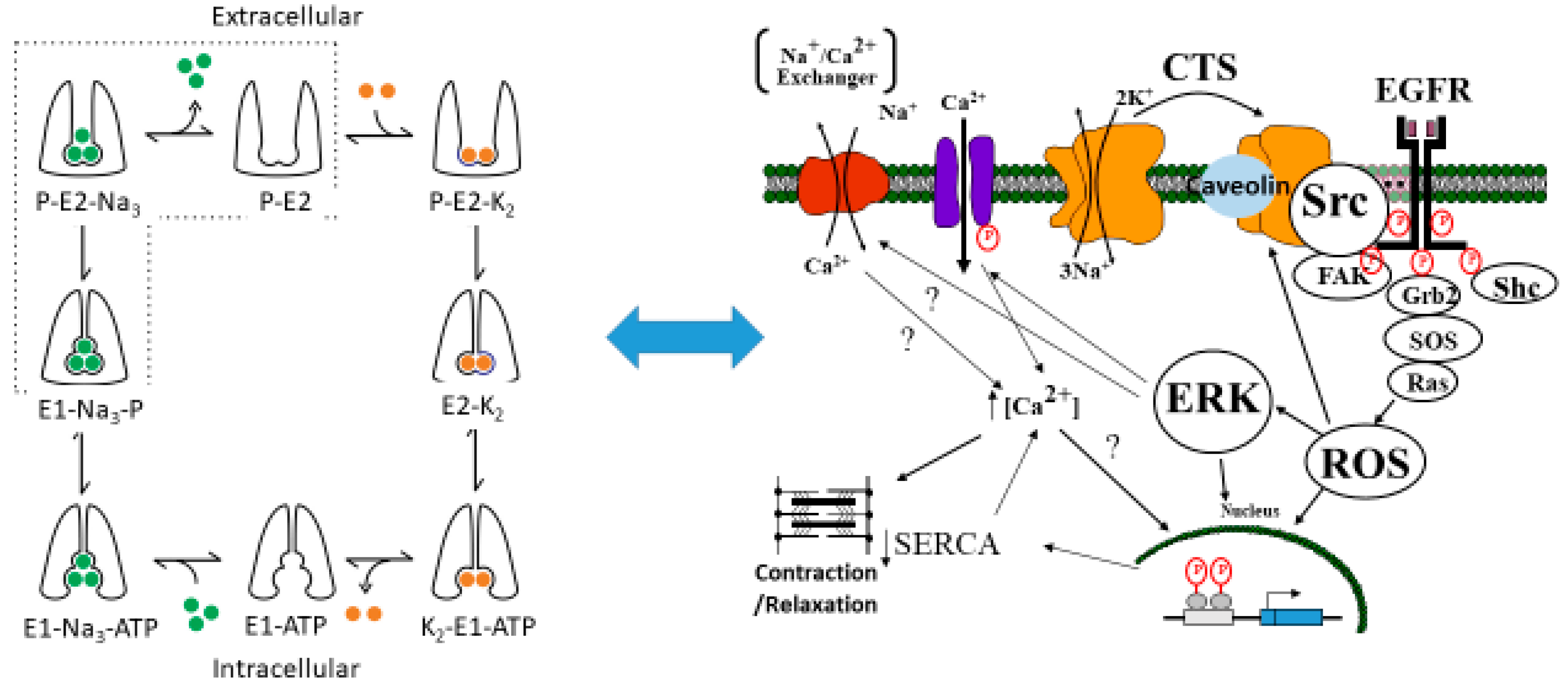
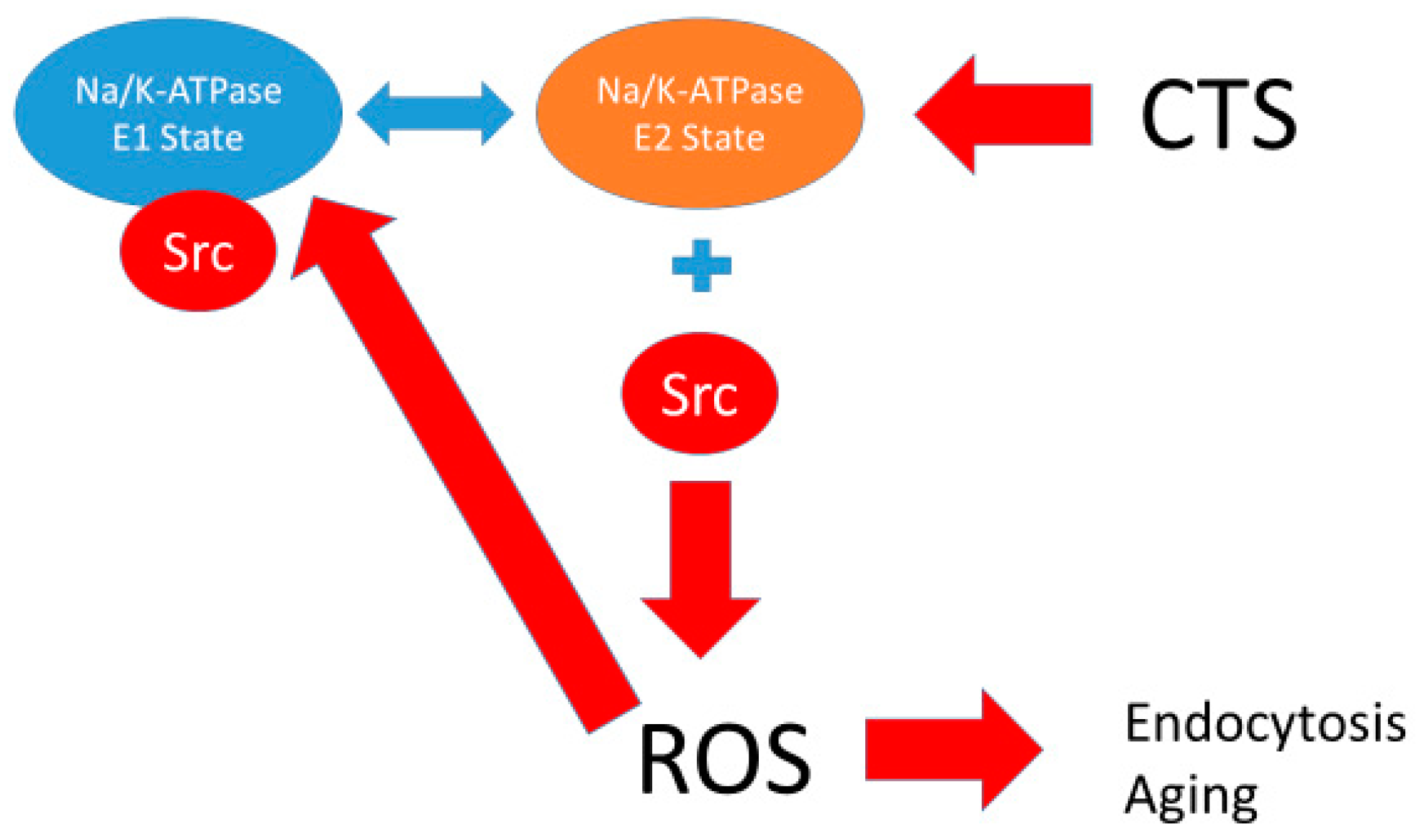
- The Na/K-ATPase α1 subunit inhibits associated Src kinase while in the E1 state. This has been suggested by modeling studies that demonstrate a number of poses by which a peptide in the N domain of the α1 subunit of the Na/K-ATPase interacts with the kinase domain of Src while in the E1 state, but an absence of such poses for the E2 state [18].
- Activation of Src kinase leads to transactivation of the epithelial growth factor receptor (EGFR) and downstream generation of reactive oxygen species.
- 3.
- ROS both result from the activation of the Na/K-ATPase-Src-EGFR signal cascade and can initiate the previously discussed activation by oxidation of the Na/K-ATPase which, in turn, decreases the tonic inhibition of associated Src Kinase. We further assume (candidly without much proof at present) that the oxidation of the Na/K-ATPase results in an increase in time than the Na/K-ATPase spends in the E2 state. The data for ouabain or other CTS increasing time in the E2 state are excellent. What is also clear is that either the binding of CTS or exposure to exogenous oxidants leads to the identical oxidation of amino acids in the A domain detailed in [18]. Alterations in the lipid environment of the Na/K-ATPase induced by oxidants is another possibility that must be studied further.
- 4.
- The level of oxidant stress determines the rate of endocytosis of the Na/K-ATPase complex from the plasma membrane which results in a net disinhibition of membrane associated Src. This axiom is supported by extensive studies examining endocytosis in porcine kidney cell line (LLC-PK1) cells and rat proximal tubule cells which both respond to either oxidants or CTS with endocytosis of the basolateral Na/K-ATPase and redistribution of the apical sodium proton antiporter isoform 3 (NHE3) in an oxidant-dependent manner [21,23,24,25].
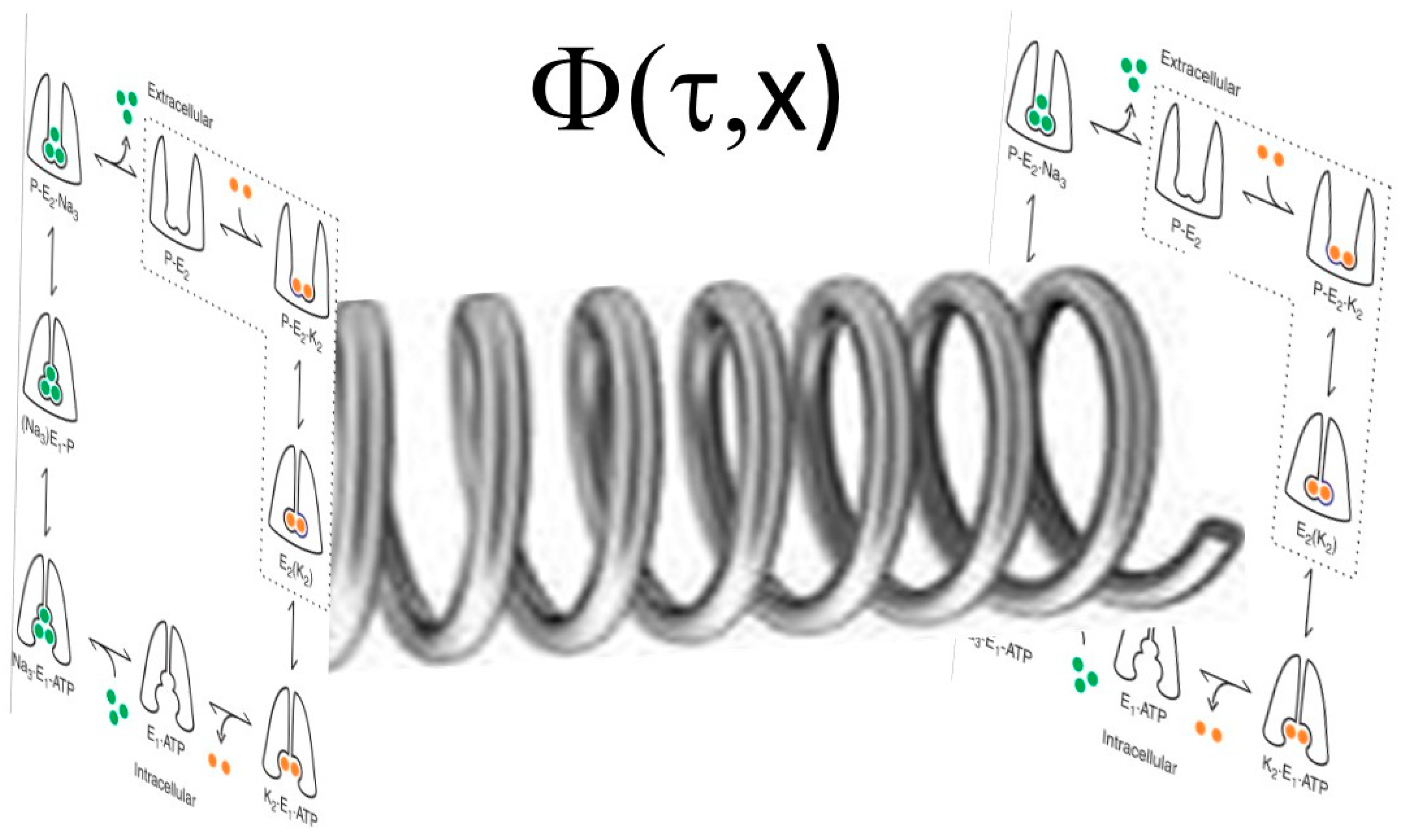
2. The Markov Chain Model
3. Superposition of States
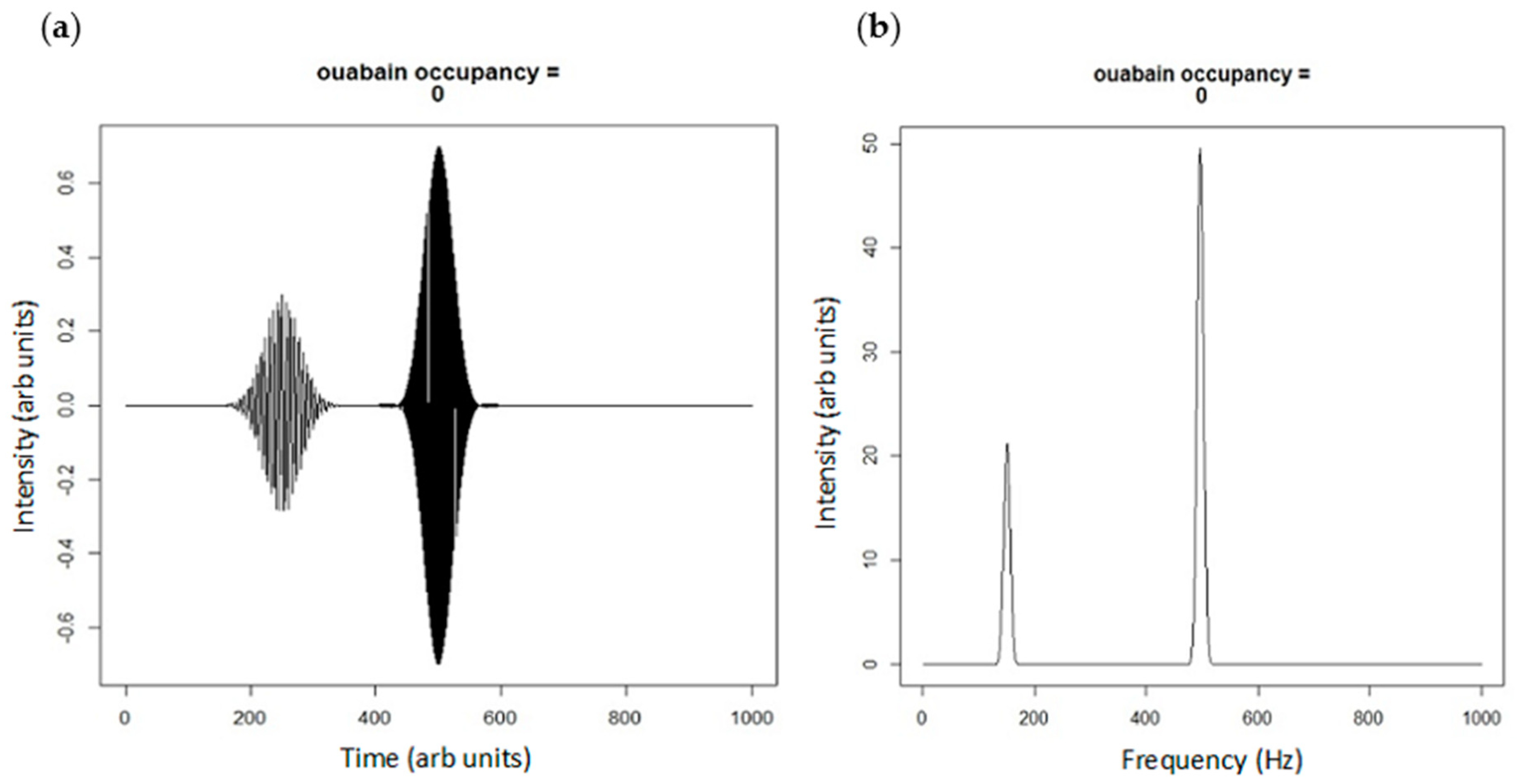
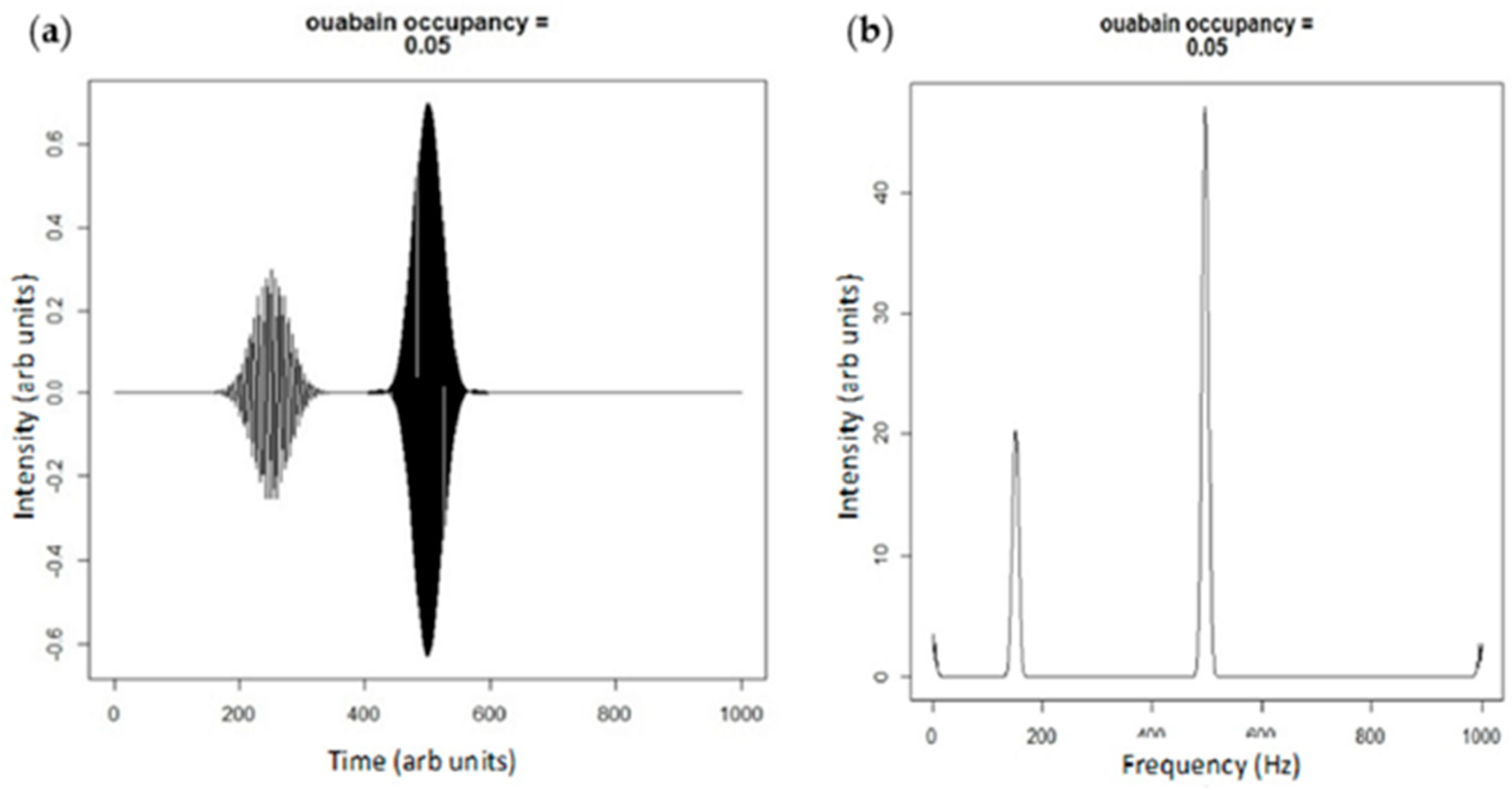
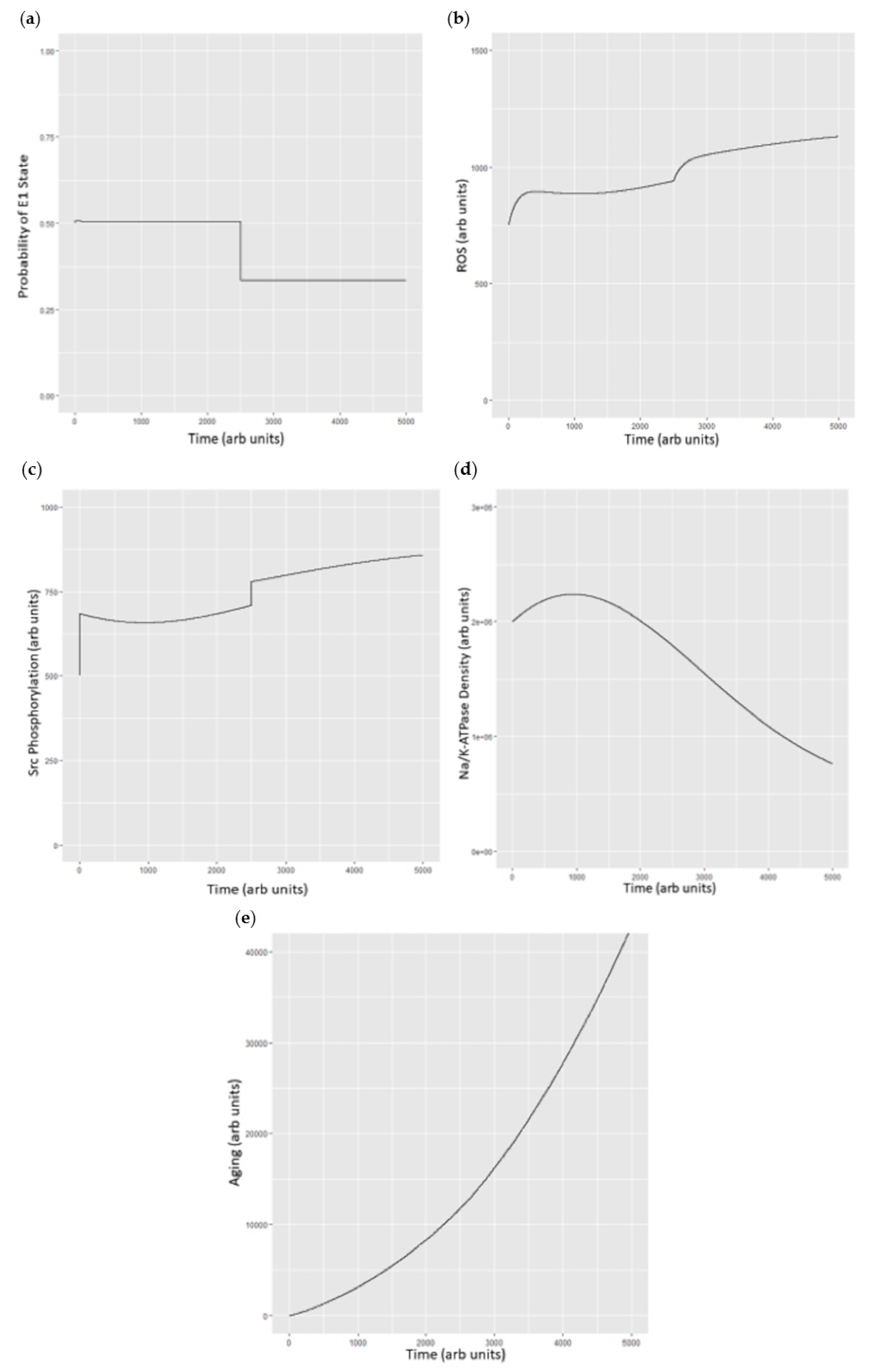
4. Extension of the Markov Chain to Define the Oxidant Amplification Loop
- The Na/K-ATPase has a basal synthesis rate.
- ROS shift the Na/K-ATPase into the E2 state (as do CTS).
- There is a basal rate of ROS production from other sources.
- There is a link between Src phosphorylation and ROS production (oversimplifying the cascade, which we believe involves transactivation of the EGFR and other steps).
- There is a detoxification rate of ROS which is proportional to ROS concentration (an assumption that is at least partially true, based on the kinetics of superoxide dismutase and catalase).
- There is a relationship coupling ROS concentration to rates of endocytosis of the Na/K-ATPase [30].
- There is a decay rate of phosphorylated Src which is proportional to the amount of phosphorylated Src.
- The Na/K-ATPase can be viewed as existing in a superposition of states which relate to each other as a Markov chain.
- The Na/K-ATPase interacts with Src and its other signaling partners using principles also seen with Markov chains. This allows modeling to be developed using a system of ordinary differential equations.
- As seen in biological systems, oxidant stress results from the shift in the Na/K-ATPase to the E2 state, which then becomes ineffective in inhibiting Src [18]. This leads to a cascade generating oxidants which further stimulates the Na/K-ATPase oxidant amplification loop (feed forward amplification) as well as oxidant injury consistent with aging.
Author Contributions
Funding
Conflicts of Interest
Appendix A. R code for Oxidant Stress/Aging Simulation
References
- Skou, J.C. Nobel lecture: The identification of the sodium-potassium pump. Angew. Chem. 1998, 37, 2320–2328. [Google Scholar] [CrossRef]
- Wagg, J.; Chapman, J.B.; Wood, S.A. A flux ratio analysis of the post-albers mechanism of the sodium pump. J. Theor. Biol. 1995, 174, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Sachs, J.R. Successes and failures of the albers-post model in predicting ion flux kinetics. Soc. Gen. Physiol. Ser. 1991, 46, 249–266. [Google Scholar] [PubMed]
- Xie, J.X.; Li, X.; Xie, Z. Regulation of renal function and structure by the signaling Na+/K+-ATPase. IUBMB Life 2013, 65, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tian, J.; Haas, M.; Shapiro, J.I.; Askari, A.; Xie, Z. Ouabain interaction with cardiac Na+/K+-ATPase initiates signal cascades independent of changes in intracellular Na+ and Ca2+ concentrations. J. Biol. Chem. 2000, 275, 27838–27844. [Google Scholar] [PubMed]
- Xie, Z.; Kometiani, P.; Liu, J.; Li, J.; Shapiro, J.I.; Askari, A. Intracellular reactive oxygen species mediate the linkage of Na+/K+-ATPase to hypertrophy and its marker genes in cardiac myocytes. J. Biol. Chem. 1999, 274, 19323–19328. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Cai, T.; Yuan, Z.; Wang, H.; Liu, L.; Haas, M.; Maksimova, E.; Huang, X.Y.; Xie, Z.J. Binding of src to Na+/K+-ATPase forms a functional signaling complex. Mol. Biol. Cell 2006, 17, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Haas, M.; Liang, M.; Cai, T.; Tian, J.; Li, S.; Xie, Z. Ouabain assembles signaling cascades through the caveolar Na+/K+-ATPase. J. Biol. Chem. 2004, 279, 17250–17259. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Wang, H.; Tian, J.; Xie, Z. Src-mediated inter-receptor cross-talk between the Na+/K+-ATPase and the epidermal growth factor receptor relays the signal from ouabain to mitogen-activated protein kinases. J. Biol. Chem. 2002, 277, 18694–18702. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yan, Y.; Nie, Y.; Shapiro, J.I. Na+/K+-ATPase signaling and salt sensitivity: The role of oxidative stress. Antioxidants 2017, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.I.; Tian, J. Signaling through the Na+/K+-ATPase: Implications for cardiac fibrosis. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H29–H30. [Google Scholar] [CrossRef] [PubMed]
- Bagrov, A.Y.; Shapiro, J.I.; Fedorova, O.V. Endogenous cardiotonic steroids: Physiology, pharmacology, and novel therapeutic targets. Pharmacol. Rev. 2009, 61, 9–38. [Google Scholar] [CrossRef] [PubMed]
- Yosef, E.; Katz, A.; Peleg, Y.; Mehlman, T.; Karlish, S.J. Do src kinase and caveolin interact directly with Na+/K+-ATPase? J. Biol. Chem. 2016, 291, 11736–11750. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.; Srikanthan, K.; Goguet-Rubio, P.; Nichols, A.; Mallick, A.; Nawab, A.; Martin, R.; Shah, P.T.; Chaudhry, M.; Sigdel, S.; et al. Pnaktide attenuates steatohepatitis and atherosclerosis by blocking Na+/K+-ATPase /ROS amplification in c57bl6 and apoe knockout mice fed a western diet. Sci. Rep. 2017, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tian, J.; Chaudhry, M.; Maxwell, K.; Yan, Y.; Wang, X.; Shah, P.T.; Khawaja, A.A.; Martin, R.; Robinette, T.J.; et al. Attenuation of Na+/K+-ATPase mediated oxidant amplification with pnaktide ameliorates experimental uremic cardiomyopathy. Sci. Rep. 2016, 6, 34592. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.; Maxwell, K.; Yan, Y.; Liu, J.; Chaudhry, M.A.; Getty, M.; Xie, Z.; Abraham, N.G.; Shapiro, J.I. Pnaktide inhibits Na+/K+-ATPase reactive oxygen species amplification and attenuates adipogenesis. Sci. Adv. 2015, 1, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Z.; Xie, J.X.; Li, X.; Tian, J.; Cai, T.; Cui, H.; Ding, H.; Shapiro, J.I.; Xie, Z. Na+/K+-ATPase mimetic pnaktide inhibits the growth of human cancer cells. J. Biol. Chem. 2011, 286, 32394–32403. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Shapiro, A.P.; Mopidevi, B.R.; Chaudhry, M.A.; Maxwell, K.; Haller, S.T.; Drummond, C.A.; Kennedy, D.J.; Tian, J.; Malhotra, D.; et al. Protein carbonylation of an amino acid residue of the Na+/K+-ATPase α1 subunit determines Na+/K+-ATPase signaling and sodium transport in renal proximal tubular cells. J. Am. Heart Assoc. 2016, 5, e003675. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Shapiro, A.P.; Haller, S.; Katragadda, V.; Liu, L.; Tian, J.; Basrur, V.; Malhotra, D.; Xie, Z.J.; Abraham, N.G.; et al. Involvement of reactive oxygen species in a feed-forward mechanism of Na+/K+-ATPase-mediated signaling transduction. J. Biol. Chem. 2013, 288, 34249–34258. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Haller, S.; Shapiro, A.; Malhotra, N.; Tian, J.; Xie, Z.; Malhotra, D.; Shapiro, J.I.; Liu, J. Ouabain-stimulated trafficking regulation of the Na+/K+-ATPase and nhe3 in renal proximal tubule cells. Mol. Cell. Biochem. 2012, 367, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kesiry, R.; Periyasamy, S.M.; Malhotra, D.; Xie, Z.; Shapiro, J.I. Ouabain induces endocytosis of plasmalemmal Na+/K+-ATPase in llc-pk1 cells by a clathrin-dependent mechanism. Kidney Int. 2004, 66, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Liu, J.; Garlid, K.D.; Shapiro, J.I.; Xie, Z. Involvement of mitogen-activated protein kinases and reactive oxygen species in the inotropic action of ouabain on cardiac myocytes. A potential role for mitochondrial k(atp) channels. Mol. Cell. Biochem. 2003, 242, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Periyasamy, S.M.; Gunning, W.; Fedorova, O.V.; Bagrov, A.Y.; Malhotra, D.; Xie, Z.; Shapiro, J.I. Effects of cardiac glycosides on sodium pump expression and function in llc-pk1 and mdck cells. Kidney Int. 2002, 62, 2118–2125. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liang, M.; Liu, L.; Malhotra, D.; Xie, Z.; Shapiro, J.I. Ouabain-induced endocytosis of the plasmalemmal Na+/K+-ATPase in llc-pk1 cells requires caveolin-1. Kidney Int. 2005, 67, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Periyasamy, S.M.; Liu, J.; Tanta, F.; Kabak, B.; Wakefield, B.; Malhotra, D.; Kennedy, D.J.; Nadoor, A.; Fedorova, O.V.; Gunning, W.; et al. Salt loading induces redistribution of the plasmalemmal Na+/K+-ATPase in proximal tubule cells. Kidney Int. 2005, 67, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Gonzalezflecha, B. Hydrogen peroxide increases Na+/K+-ATPase function in alveolar typeII cells. Biochim. Biophy. Acta 1996, 1290, 46–52. [Google Scholar] [CrossRef]
- Spahn, P.N.; Hansen, A.H.; Kol, S.; Voldborg, B.G.; Lewis, N.E. Predictive glycoengineering of biosimilars using a markov chain glycosylation model. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Kou, S.C. Statistics and related topics in single-molecule biophysics. Annu. Rev. Stat. Appl. 2014, 1, 465–492. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Tian, J.; Liu, L.; Pierre, S.; Liu, J.; Shapiro, J.; Xie, Z.J. Identification of a pool of non-pumping Na+/K+-ATPase. J. Biol. Chem. 2007, 282, 10585–10593. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kennedy, D.J.; Yan, Y.; Shapiro, J.I. Reactive oxygen species modulation of Na+/K+-ATPase regulates fibrosis and renal proximal tubular sodium handling. Int. J. Nephrol. 2012, 2012, 381320. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Wang, Y.; Liu, G.; Zolotarjova, N.; Periyasamy, S.M.; Askari, A. Similarities and differences between the properties of native and recombinant Na+/K+-ATPase. Arch. Biochem. Biophys. 1996, 330, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.; Nichols, A.; Mallick, A.; Klug, R.L.; Liu, J.; Wang, X.; Srikanthan, K.; Goguet-Rubio, P.; Nawab, A.; Pratt, R.; et al. The Na+/K+-ATPase oxidant amplification loop regulates aging. Sci. Rep. 2018, 8, 9721. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Shapiro, J.I. Quantum Modeling: A Bridge between the Pumping and Signaling Functions of Na/K-ATPase. Int. J. Mol. Sci. 2018, 19, 2347. https://doi.org/10.3390/ijms19082347
Wang W, Shapiro JI. Quantum Modeling: A Bridge between the Pumping and Signaling Functions of Na/K-ATPase. International Journal of Molecular Sciences. 2018; 19(8):2347. https://doi.org/10.3390/ijms19082347
Chicago/Turabian StyleWang, Weiguang, and Joseph I. Shapiro. 2018. "Quantum Modeling: A Bridge between the Pumping and Signaling Functions of Na/K-ATPase" International Journal of Molecular Sciences 19, no. 8: 2347. https://doi.org/10.3390/ijms19082347
APA StyleWang, W., & Shapiro, J. I. (2018). Quantum Modeling: A Bridge between the Pumping and Signaling Functions of Na/K-ATPase. International Journal of Molecular Sciences, 19(8), 2347. https://doi.org/10.3390/ijms19082347