Human Exonuclease 1 (EXO1) Regulatory Functions in DNA Replication with Putative Roles in Cancer

 , and
, and
Abstract
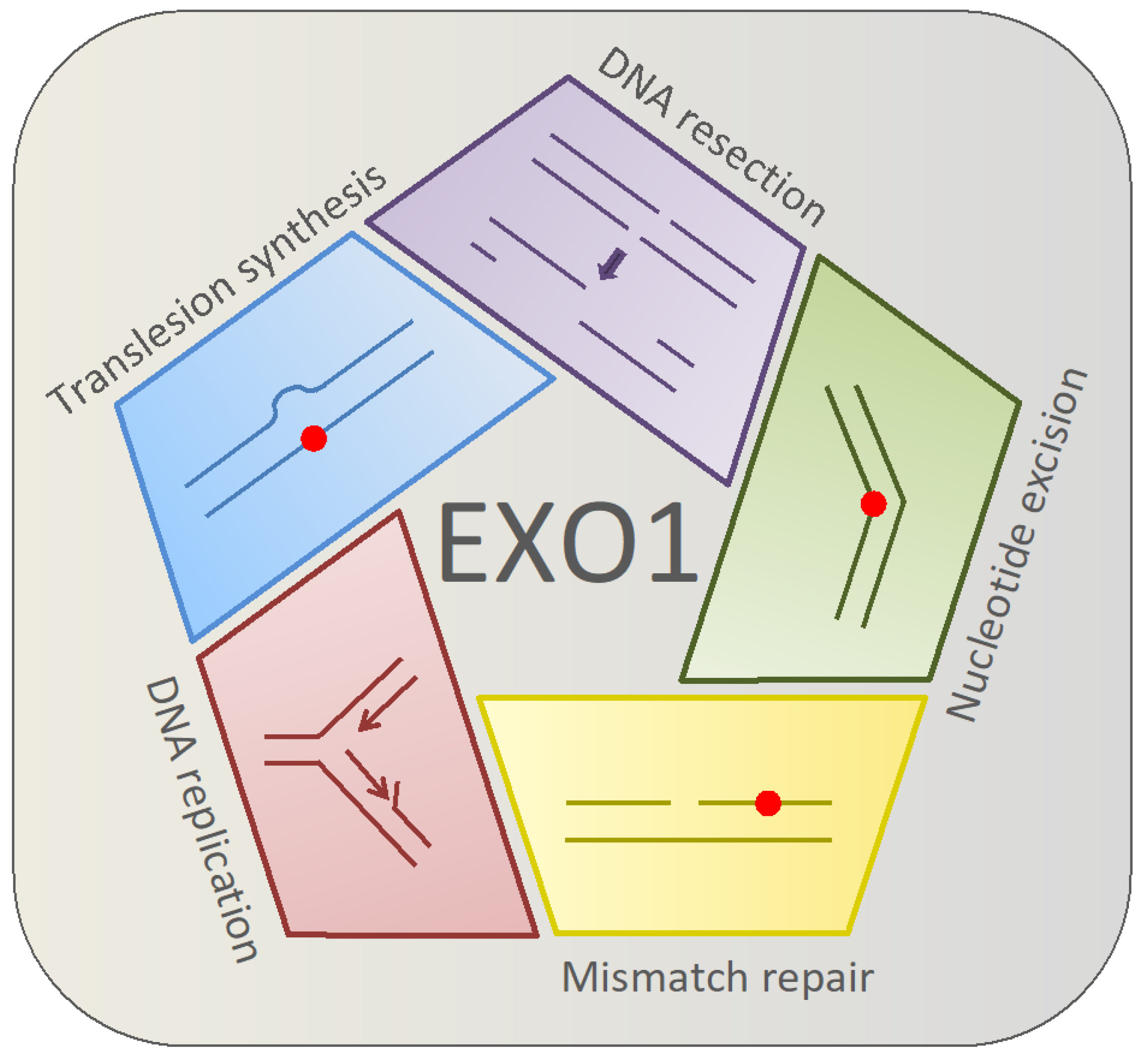
:1. Introduction
2. DNA Replication
3. Mismatch Repair
4. Translesion DNA Synthesis
5. Nucleotide Excision Repair
6. Homologous Recombination and DNA End Resection
7. Cell Cycle Regulation
8. Link to Cancer
9. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Cotta-Ramusino, C.; Fachinetti, D.; Lucca, C.; Doksani, Y.; Lopes, M.; Sogo, J.; Foiani, M. Exo1 processes stalled replication forks and counteracts fork reversal in checkpoint-defective cells. Mol. Cell 2005, 17, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Szankasi, P.; Smith, G.R. A role for exonuclease I. from S. pombe in mutation avoidance and mismatch correction. Science 1995, 267, 1166–1169. [Google Scholar] [CrossRef] [PubMed]
- Sertic, S.; Mollica, A.; Campus, I.; Roma, S.; Tumini, E.; Aguilera, A.; Muzi-Falconi, M. Coordinated Activity of Y Family TLS Polymerases and EXO1 Protects Non-S Phase Cells from UV-Induced Cytotoxic Lesions. Mol. Cell 2018, 70, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Guan, M.X.; Bailis, A.M.; Shen, B. Saccharomyces cerevisiae exonuclease-1 plays a role in UV resistance that is distinct from nucleotide excision repair. Nucleic Acids Res. 1998, 26, 3077–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolderson, E.; Tomimatsu, N.; Richard, D.J.; Boucher, D.; Kumar, R.; Pandita, T.K.; Burma, S.; Khanna, K.K. Phosphorylation of Exo1 modulates homologous recombination repair of DNA double-strand breaks. Nucleic Acids Res. 2010, 38, 1821–1831. [Google Scholar] [CrossRef] [Green Version]
- Keijzers, G.; Liu, D.; Rasmussen, L.J. Exonuclease 1 and its versatile roles in DNA repair. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 440–451. [Google Scholar] [CrossRef]
- Wilson, D.M., 3rd; Carney, J.P.; Coleman, M.A.; Adamson, A.W.; Christensen, M.; Lamerdin, J.E. Hex1: A new human Rad2 nuclease family member with homology to yeast exonuclease 1. Nucleic Acids Res. 1998, 26, 3762–3768. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Qian, Y.; Chen, V.; Guan, M.X.; Shen, B. Human exonuclease 1 functionally complements its yeast homologues in DNA recombination, RNA primer removal, and mutation avoidance. J. Biol. Chem. 1999, 274, 17893–17900. [Google Scholar] [CrossRef]
- Lee, B.I.; Wilson, D.M., 3rd. The RAD2 domain of human exonuclease 1 exhibits 5’ to 3’ exonuclease and flap structure-specific endonuclease activities. J. Biol. Chem. 1999, 274, 37763–37769. [Google Scholar] [CrossRef]
- Genschel, J.; Modrich, P. Mechanism of 5’-directed excision in human mismatch repair. Mol. Cell 2003, 12, 1077–1086. [Google Scholar] [CrossRef]
- Keijzers, G.; Bohr, V.A.; Rasmussen, L.J. Human exonuclease 1 (EXO1) activity characterization and its function on flap structures. Biosci. Rep. 2015, 35, e00206. [Google Scholar] [CrossRef]
- Zakharyevich, K.; Ma, Y.; Tang, S.; Hwang, P.Y.; Boiteux, S.; Hunter, N. Temporally and biochemically distinct activities of Exo1 during meiosis: Double-strand break resection and resolution of double Holliday junctions. Mol. Cell 2010, 40, 1001–1015. [Google Scholar] [CrossRef]
- Segurado, M.; Diffley, J.F. Separate roles for the DNA damage checkpoint protein kinases in stabilizing DNA replication forks. Genes Dev. 2008, 22, 1816–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Nickoloff, J.A.; Wu, Y.; Williamson, E.A.; Sidhu, G.S.; Reinert, B.L.; Jaiswal, A.S.; Srinivasan, G.; Patel, B.; Kong, K.; et al. Endonuclease EEPD1 Is a Gatekeeper for Repair of Stressed Replication Forks. J. Biol. Chem. 2017, 292, 2795–2804. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.L.; Bambara, R.A. Reconstituted Okazaki fragment processing indicates two pathways of primer removal. J. Biol. Chem. 2006, 281, 26051–26061. [Google Scholar] [CrossRef]
- Moreau, S.; Morgan, E.A.; Symington, L.S. Overlapping functions of the Saccharomyces cerevisiae Mre11, Exo1 and Rad27 nucleases in DNA metabolism. Genetics 2001, 159, 1423–1433. [Google Scholar] [PubMed]
- Sparks, J.L.; Chon, H.; Cerritelli, S.M.; Kunkel, T.A.; Johansson, E.; Crouch, R.J.; Burgers, P.M. RNase H2-initiated ribonucleotide excision repair. Mol. Cell 2012, 47, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, P.; Huang, K.N.; Tishkoff, D.X.; Kolodner, R.D.; Symington, L.S. Exonuclease I of Saccharomyces cerevisiae functions in mitotic recombination in vivo and in vitro. Mol. Cell. Biol. 1997, 17, 2764–2773. [Google Scholar] [CrossRef] [PubMed]
- Llorente, B.; Symington, L.S. The Mre11 nuclease is not required for 5’ to 3’ resection at multiple HO-induced double-strand breaks. Mol. Cell. Biol. 2004, 24, 9682–9694. [Google Scholar] [CrossRef]
- Stith, C.M.; Sterling, J.; Resnick, M.A.; Gordenin, D.A.; Burgers, P.M. Flexibility of eukaryotic Okazaki fragment maturation through regulated strand displacement synthesis. J. Biol. Chem. 2008, 283, 34129–34140. [Google Scholar] [CrossRef]
- Tishkoff, D.X.; Filosi, N.; Gaida, G.M.; Kolodner, R.D. A novel mutation avoidance mechanism dependent on S. cerevisiae RAD27 is distinct from DNA mismatch repair. Cell 1997, 88, 253–263. [Google Scholar] [CrossRef]
- Sharma, S.; Sommers, J.A.; Driscoll, H.C.; Uzdilla, L.; Wilson, T.M.; Brosh, R.M., Jr. The exonucleolytic and endonucleolytic cleavage activities of human exonuclease 1 are stimulated by an interaction with the carboxyl-terminal region of the Werner syndrome protein. J. Biol. Chem. 2003, 278, 23487–23496. [Google Scholar] [CrossRef] [PubMed]
- Doherty, K.M.; Sharma, S.; Uzdilla, L.A.; Wilson, T.M.; Cui, S.; Vindigni, A.; Brosh, R.M., Jr. RECQ1 helicase interacts with human mismatch repair factors that regulate genetic recombination. J. Biol. Chem. 2005, 280, 28085–28094. [Google Scholar] [CrossRef] [PubMed]
- Pike, J.E.; Henry, R.A.; Burgers, P.M.; Campbell, J.L.; Bambara, R.A. An alternative pathway for Okazaki fragment processing: Resolution of fold-back flaps by Pif1 helicase. J. Biol. Chem. 2010, 285, 41712–41723. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.L.; Pike, J.E.; Wang, W.; Burgers, P.M.; Campbell, J.L.; Bambara, R.A. Pif1 helicase directs eukaryotic Okazaki fragments toward the two-nuclease cleavage pathway for primer removal. J. Biol. Chem. 2008, 283, 27483–27493. [Google Scholar] [CrossRef]
- Gloor, J.W.; Balakrishnan, L.; Campbell, J.L.; Bambara, R.A. Biochemical analyses indicate that binding and cleavage specificities define the ordered processing of human Okazaki fragments by Dna2 and FEN1. Nucleic Acids Res. 2012, 40, 6774–6786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munashingha, P.R.; Lee, C.H.; Kang, Y.H.; Shin, Y.K.; Nguyen, T.A.; Seo, Y.S. The trans-autostimulatory activity of Rad27 suppresses dna2 defects in Okazaki fragment processing. J. Biol. Chem. 2012, 287, 8675–8687. [Google Scholar] [CrossRef]
- Zaher, M.S.; Rashid, F.; Song, B.; Joudeh, L.I.; Sobhy, M.A.; Tehseen, M.; Hingorani, M.M.; Hamdan, S.M. Missed cleavage opportunities by FEN1 lead to Okazaki fragment maturation via the long-flap pathway. Nucleic Acids Res. 2018, 46, 2956–2974. [Google Scholar] [CrossRef] [Green Version]
- Duxin, J.P.; Moore, H.R.; Sidorova, J.; Karanja, K.; Honaker, Y.; Dao, B.; Piwnica-Worms, H.; Campbell, J.L.; Monnat, R.J., Jr.; Stewart, S.A. Okazaki fragment processing-independent role for human Dna2 enzyme during DNA replication. J. Biol. Chem. 2012, 287, 21980–21991. [Google Scholar] [CrossRef]
- Levikova, M.; Cejka, P. The Saccharomyces cerevisiae Dna2 can function as a sole nuclease in the processing of Okazaki fragments in DNA replication. Nucleic Acids Res. 2015, 43, 7888–7897. [Google Scholar] [CrossRef]
- Hsieh, P.; Yamane, K. DNA mismatch repair: Molecular mechanism, cancer, and ageing. Mech. Ageing Dev. 2008, 129, 391–407. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Keijzers, G.; Rasmussen, L.J. DNA mismatch repair and its many roles in eukaryotic cells. Mutat. Res. 2017, 773, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, T.A.; Erie, D.A. DNA mismatch repair. Annu. Rev. Biochem. 2005, 74, 681–710. [Google Scholar] [CrossRef] [PubMed]
- Tishkoff, D.X.; Boerger, A.L.; Bertrand, P.; Filosi, N.; Gaida, G.M.; Kane, M.F.; Kolodner, R.D. Identification and characterization of Saccharomyces cerevisiae EXO1, a gene encoding an exonuclease that interacts with MSH2. Proc. Natl. Acad. Sci. USA 1997, 94, 7487–7492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.T.; Gordenin, D.A.; Resnick, M.A. The 3’-->5’ exonucleases of DNA polymerases delta and epsilon and the 5’-->3’ exonuclease Exo1 have major roles in postreplication mutation avoidance in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 2000–2007. [Google Scholar] [CrossRef] [PubMed]
- Schmutte, C.; Sadoff, M.M.; Shim, K.S.; Acharya, S.; Fishel, R. The interaction of DNA mismatch repair proteins with human exonuclease I. J. Biol. Chem. 2001, 276, 33011–33018. [Google Scholar] [CrossRef] [PubMed]
- Schmutte, C.; Marinescu, R.C.; Sadoff, M.M.; Guerrette, S.; Overhauser, J.; Fishel, R. Human exonuclease I interacts with the mismatch repair protein hMSH2. Cancer Res. 1998, 58, 4537–4542. [Google Scholar]
- Jäger, A.C.; Rasmussen, M.; Bisgaard, H.C.; Singh, K.K.; Nielsen, F.C.; Rasmussen, L.J. HNPCC mutations in the human DNA mismatch repair gene hMLH1 influence assembly of hMutLalpha and hMLH1-hEXO1 complexes. Oncogene 2001, 20, 3590–3595. [Google Scholar] [CrossRef]
- Amin, N.S.; Nguyen, M.N.; Oh, S.; Kolodner, R.D. exo1-Dependent mutator mutations: Model system for studying functional interactions in mismatch repair. Mol. Cell. Biol. 2001, 21, 5142–5155. [Google Scholar] [CrossRef]
- Nielsen, F.C.; Jäger, A.C.; Lützen, A.; Bundgaard, J.R.; Rasmussen, L.J. Characterization of human exonuclease 1 in complex with mismatch repair proteins, subcellular localization and association with PCNA. Oncogene 2004, 23, 1457–1468. [Google Scholar] [CrossRef]
- Liberti, S.E.; Andersen, S.D.; Wang, J.; May, A.; Miron, S.; Perderiset, M.; Keijzers, G.; Nielsen, F.C.; Charbonnier, J.B.; Bohr, V.A.; et al. Bi-directional routing of DNA mismatch repair protein human exonuclease 1 to replication foci and DNA double strand breaks. DNA Repair 2011, 10, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Goellner, E.M.; Putnam, C.D.; Graham, W.J., 5th; Rahal, C.M.; Li, B.Z.; Kolodner, R.D. Identification of Exo1-Msh2 interaction motifs in DNA mismatch repair and new Msh2-binding partners. Nat. Struct. Mol. Biol. 2018, 25, 650–659. [Google Scholar] [CrossRef]
- Chen, H.; Lisby, M.; Symington, L.S. RPA coordinates DNA end resection and prevents formation of DNA hairpins. Mol. Cell 2013, 50, 589–600. [Google Scholar] [CrossRef]
- Li, G.M. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008, 18, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Pedroni, M.; Tamassia, M.G.; Percesepe, A.; Roncucci, L.; Benatti, P.; Lanza, G., Jr.; Gafà, R.; Di Gregorio, C.; Fante, R.; Losi, L.; et al. Microsatellite instability in multiple colorectal tumors. Int. J. Cancer 1999, 81, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Keijzers, G.; Bakula, D.; Scheibye-Knudsen, M. Monogenic Diseases of DNA Repair. N. Engl. J. Med. 2017, 377, 1868–1876. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, N.C.; Littman, S.J.; Modrich, P.; Kinzler, K.W.; Vogelstein, B. A naturally occurring hPMS2 mutation can confer a dominant negative mutator phenotype. Mol. Cell. Biol. 1998, 18, 1635–1641. [Google Scholar] [CrossRef]
- Kadyrov, F.A.; Genschel, J.; Fang, Y.; Penland, E.; Edelmann, W.; Modrich, P. A possible mechanism for exonuclease 1-independent eukaryotic mismatch repair. Proc. Natl. Acad. Sci. USA 2009, 106, 8495–8500. [Google Scholar] [CrossRef] [Green Version]
- Goellner, E.M.; Smith, C.E.; Campbell, C.S.; Hombauer, H.; Desai, A.; Putnam, C.D.; Kolodner, R.D. PCNA and Msh2-Msh6 activate an Mlh1-Pms1 endonuclease pathway required for Exo1-independent mismatch repair. Mol. Cell 2014, 55, 291–304. [Google Scholar] [CrossRef]
- Saydam, N.; Kanagaraj, R.; Dietschy, T.; Garcia, P.L.; Peña-Diaz, J.; Shevelev, I.; Stagljar, I.; Janscak, P. Physical and functional interactions between Werner syndrome helicase and mismatch-repair initiation factors. Nucleic Acids Res. 2007, 35, 5706–57016. [Google Scholar] [CrossRef]
- Machwe, A.; Lozada, E.; Wold, M.S.; Li, G.M.; Orren, D.K. Molecular cooperation between the Werner syndrome protein and replication protein A in relation to replication fork blockage. J. Biol. Chem. 2011, 286, 3497–3508. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Ohnishi, M.; Suemoto, Y.; Kirkner, G.J.; Liu, Z.; Yamamoto, H.; Loda, M.; Fuchs, C.S.; Ogino, S. WRN promoter methylation possibly connects mucinous differentiation, microsatellite instability and CpG island methylator phenotype in colorectal cancer. Mod. Pathol. 2008, 21, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.D.; Shen, J.C.; Kamath-Loeb, A.S.; Blank, A.; Sopher, B.L.; Martin, G.M.; Oshima, J.; Loeb, L.A. The Werner syndrome protein is a DNA helicase. Nat. Genet. 1997, 17, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Sommers, J.A.; Sharma, S.; Doherty, K.M.; Karmakar, P.; Yang, Q.; Kenny, M.K.; Harris, C.C.; Brosh, R.M., Jr. p53 modulates RPA-dependent and RPA-independent WRN helicase activity. Cancer Res. 2005, 65, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Constantinou, A.; Tarsounas, M.; Karow, J.K.; Brosh, R.M.; Bohr, V.A.; Hickson, I.D.; West, S.C. Werner’s syndrome protein (WRN) migrates Holliday junctions and co-localizes with RPA upon replication arrest. EMBO Rep. 2000, 1, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Cunniff, C.; Bassetti, J.A.; Ellis, N.A. Bloom’s Syndrome: Clinical Spectrum, Molecular Pathogenesis, and Cancer Predisposition. Mol. Syndromol. 2017, 8, 4–23. [Google Scholar] [CrossRef] [PubMed]
- Pedrazzi, G.; Perrera, C.; Blaser, H.; Kuster, P.; Marra, G.; Davies, S.L.; Ryu, G.H.; Freire, R.; Hickson, I.D.; Jiricny, J.; et al. Direct association of Bloom’s syndrome gene product with the human mismatch repair protein MLH1. Nucleic Acids Res. 2001, 29, 4378–4386. [Google Scholar] [CrossRef] [PubMed]
- Pedrazzi, G.; Bachrati, C.Z.; Selak, N.; Studer, I.; Petkovic, M.; Hickson, I.D.; Jiricny, J.; Stagljar, I. The Bloom’s syndrome helicase interacts directly with the human DNA mismatch repair protein hMSH6. Biol. Chem. 2003, 384, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Sommers, J.A.; Banerjee, T.; Hinds, T.; Wan, B.; Wold, M.S.; Lei, M.; Brosh, R.M., Jr. Novel function of the Fanconi anemia group J or RECQ1 helicase to disrupt protein-DNA complexes in a replication protein A-stimulated manner. J. Biol. Chem. 2014, 289, 19928–19941. [Google Scholar] [CrossRef]
- Banerjee, T.; Sommers, J.A.; Huang, J.; Seidman, M.M.; Brosh, R.M., Jr. Catalytic strand separation by RECQ1 is required for RPA-mediated response to replication stress. Curr. Biol. 2015, 25, 2830–2838. [Google Scholar] [CrossRef]
- Doherty, K.M.; Sommers, J.A.; Gray, M.D.; Lee, J.W.; von Kobbe, C.; Thoma, N.H.; Kureekattil, R.P.; Kenny, M.K.; Brosh, R.M., Jr. Physical and functional mapping of the replication protein a interaction domain of the werner and bloom syndrome helicases. J. Biol. Chem. 2005, 280, 29494–29505. [Google Scholar] [CrossRef] [PubMed]
- Langland, G.; Kordich, J.; Creaney, J.; Goss, K.H.; Lillard-Wetherell, K.; Bebenek, K.; Kunkel, T.A.; Groden, J. The Bloom’s syndrome protein (BLM) interacts with MLH1 but is not required for DNA mismatch repair. J. Biol. Chem. 2001, 276, 30031–30035. [Google Scholar] [CrossRef] [PubMed]
- Bennett, S.E.; Umar, A.; Oshima, J.; Monnat, R.J., Jr.; Kunkel, T.A. Mismatch repair in extracts of Werner syndrome cell lines. Cancer Res. 1997, 57, 2956–2960. [Google Scholar] [PubMed]
- Desai, A.; Gerson, S. Exo1 independent DNA mismatch repair involves multiple compensatory nucleases. DNA Repair 2014, 21, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Frederiksen, J.H.; Liberti, S.E.; Lützen, A.; Keijzers, G.; Pena-Diaz, J.; Rasmussen, L.J. Human DNA polymerase delta double-mutant D316A; E318A interferes with DNA mismatch repair in vitro. Nucleic Acids Res. 2017, 45, 9427–9440. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.E.; Mendillo, M.L.; Bowen, N.; Hombauer, H.; Campbell, C.S.; Desai, A.; Putnam, C.D.; Kolodner, R.D. Dominant mutations in S. cerevisiae PMS1 identify the Mlh1- Pms1 endonuclease active site and an exonuclease 1-independent mismatch repair pathway. PLoS Genet. 2013, 9, e1003869. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.T.; Fey, J.P.; Erdeniz, N.; Gellon, L.; Boiteux, S.; Liskay, R.M. A mutation in EXO1 defines separable roles in DNA mismatch repair and post-replication repair. DNA Repair 2007, 6, 1572–1583. [Google Scholar] [CrossRef] [Green Version]
- Karras, G.I.; Fumasoni, M.; Sienski, G.; Vanoli, F.; Branzei, D.; Jentsch, S. Noncanonical role of the 9-1-1 clamp in the error-free DNA damage tolerance pathway. Mol. Cell 2013, 49, 536–546. [Google Scholar] [CrossRef]
- Friedberg, E.C. How nucleotide excision repair protects against cancer. Nat. Rev. Cancer 2001, 1, 22–33. [Google Scholar] [CrossRef]
- Giannattasio, M.; Follonier, C.; Tourrière, H.; Puddu, F.; Lazzaro, F.; Pasero, P.; Lopes, M.; Plevani, P.; Muzi-Falconi, M. Exo1 competes with repair synthesis, converts NER intermediates to long ssDNA gaps, and promotes checkpoint activation. Mol. Cell 2010, 40, 50–62. [Google Scholar] [CrossRef]
- Schaetzlein, S.; Chahwan, R.; Avdievich, E.; Roa, S.; Wei, K.; Eoff, R.L.; Sellers, R.S.; Clark, A.B.; Kunkel, T.A.; Scharff, M.D.; et al. Mammalian Exo1 encodes both structural and catalytic functions that play distinct roles in essential biological processes. Proc. Natl. Acad. Sci. USA 2013, 110, E2470–E2479. [Google Scholar] [CrossRef] [PubMed]
- Modrich, P.; Lahue, R. Mismatch repair in replication fidelity, genetic recombination, and cancer biology. Annu. Rev. Biochem. 1996, 65, 101–133. [Google Scholar] [CrossRef] [PubMed]
- El-Shemerly, M.; Hess, D.; Pyakurel, A.K.; Moselhy, S.; Ferrari, S. ATR-dependent pathways control hEXO1 stability in response to stalled forks. Nucleic Acids Res. 2008, 6, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Mimitou, E.P.; Symington, L.S. Sae2, Exo1 and Sgs1 collaborate in DNA double-strand break processing. Nature 2008, 455, 770–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheruiyot, A.; Paudyal, S.C.; Kim, I.K.; Sparks, M.; Ellenberger, T.; Piwnica-Worms, H.; You, Z. Poly(ADP-ribose)-binding promotes Exo1 damage recruitment and suppresses its nuclease activities. DNA Repair 2015, 35, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Shi, J.; Chen, S.H.; Bian, C.; Yu, X. The PIN domain of EXO1 recognizes poly(ADP-ribose) in DNA damage response. Nucleic Acids Res. 2015, 43, 10782–10794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Kadyrov, F.A.; Modrich, P. PARP-1 enhances the mismatch-dependence of 5’-directed excision in human mismatch repair in vitro. DNA Repair 2011, 10, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Farah, J.A.; Cromie, G.A.; Smith, G.R. Ctp1 and Exonuclease 1, alternative nucleases regulated by the MRN complex, are required for efficient meiotic recombination. Proc. Natl. Acad. Sci. USA 2009, 106, 9356–9361. [Google Scholar] [CrossRef] [Green Version]
- Nimonkar, A.V.; Genschel, J.; Kinoshita, E.; Polaczek, P.; Campbell, J.L.; Wyman, C.; Modrich, P.; Kowalczykowski, S.C. BLM-DNA2-RPA-MRN and EXO1-BLM-RPA-MRN constitute two DNA end resection machineries for human DNA break repair. Genes Dev. 2011, 25, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.H.; Zhou, R.; Campbell, J.; Chen, J.; Ha, T.; Paull, T.T. The SOSS1 single-stranded DNA binding complex promotes DNA end resection in concert with Exo1. EMBO J. 2013, 32, 126–139. [Google Scholar] [CrossRef]
- Williams, B.R.; Mirzoeva, O.K.; Morgan, W.F.; Lin, J.; Dunnick, W.; Petrini, J.H. A murine model of Nijmegen breakage syndrome. Curr. Biol. 2002, 12, 648–653. [Google Scholar] [CrossRef]
- Rein, K.; Yanez, D.A.; Terré, B.; Palenzuela, L.; Aivio, S.; Wei, K.; Edelmann, W.; Stark, J.M.; Stracker, T.H. EXO1 is critical for embryogenesis and the DNA damage response in mice with a hypomorphic Nbs1 allele. Nucleic Acids Res. 2015, 43, 7371–7387. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Putnam, C.D.; Kane, M.F.; Zhang, W.; Edelmann, L.; Russell, R.; Carrión, D.V.; Chin, L.; Kucherlapati, R.; Kolodner, R.D.; et al. Mutation in Rpa1 results in defective DNA double-strand break repair, chromosomal instability and cancer in mice. Nat. Genet. 2005, 37, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S. DNA end resection by CtIP and exonuclease 1 prevents genomic instability. EMBO Rep. 2010, 11, 962–968. [Google Scholar] [Green Version]
- Shim, E.Y.; Chung, W.H.; Nicolette, M.L.; Zhang, Y.; Davis, M.; Zhu, Z.; Paull, T.T.; Ira, G.; Lee, S.E. Saccharomyces cerevisiae Mre11/Rad50/Xrs2 and Ku proteins regulate association of Exo1 and Dna2 with DNA breaks. EMBO J. 2010, 29, 3370–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Daley, J.M.; Kwon, Y.; Krasner, D.S.; Sung, P. Plasticity of the Mre11-Rad50-Xrs2-Sae2 nuclease ensemble in the processing of DNA-bound obstacles. Genes Dev. 2017, 31, 2331–2336. [Google Scholar] [CrossRef] [PubMed]
- Rulten, S.L.; Grundy, G.J. Non-homologous end joining: Common interaction sites and exchange of multiple factors in the DNA repair process. Bioessays 2017, 39, 1600209. [Google Scholar] [CrossRef]
- Kragelund, B.B.; Weterings, E.; Hartmann-Petersen, R.; Keijzers, G. The Ku70/80 ring in Non-Homologous End-Joining: Easy to slip on, hard to remove. Front. Biosci. 2016, 21, 514–527. [Google Scholar]
- Shamanna, R.A.; Lu, H.; de Freitas, J.K.; Tian, J.; Croteau, D.L.; Bohr, V.A. WRN regulates pathway choice between classical and alternative non-homologous end joining. Nat. Commun. 2016, 7, 13785. [Google Scholar] [CrossRef] [Green Version]
- Yu, A.M.; McVey, M. Synthesis-dependent microhomology-mediated end joining accounts for multiple types of repair junctions. Nucleic Acids Res. 2010, 38, 5706–5717. [Google Scholar] [CrossRef] [Green Version]
- Keijzers, G.; Maynard, S.; Shamanna, R.A.; Rasmussen, L.J.; Croteau, D.L.; Bohr, V.A. The role of RecQ helicases in non-homologous end-joining. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, M.; Sommers, J.A.; Morris, C.; Brosh, R.M., Jr. Delineation of WRN helicase function with EXO1 in the replicational stress response. DNA Repair 2010, 9, 765–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannascoli, C.; Palermo, V.; Murfuni, I.; Franchitto, A.; Pichierri, P. The WRN exonuclease domain protects nascent strands from pathological MRE11/EXO1-dependent degradation. Nucleic Acids Res. 2015, 43, 9788–9803. [Google Scholar] [CrossRef] [PubMed]
- Genschel, J.; Kadyrova, L.Y.; Iyer, R.R.; Dahal, B.K.; Kadyrov, F.A.; Modrich, P. Interaction of proliferating cell nuclear antigen with PMS2 is required for MutLα activation and function in mismatch repair. Proc. Natl. Acad. Sci. USA 2017, 114, 4930–4935. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Paudyal, S.C.; Chin, R.I.; You, Z. PCNA promotes processive DNA end resection by Exo1. Nucleic Acids Res. 2013, 41, 9325–9338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engels, K.; Giannattasio, M.; Muzi-Falconi, M.; Lopes, M.; Ferrari, S. 14-3-3 Proteins regulate exonuclease 1-dependent processing of stalled replication forks. PLoS Genet. 2011, 7, e1001367. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.D.; Keijzers, G.; Rampakakis, E.; Engels, K.; Luhn, P.; El-Shemerly, M.; Nielsen, F.C.; Du, Y.; May, A.; Bohr, V.A.; et al. 14-3-3 checkpoint regulatory proteins interact specifically with DNA repair protein human exonuclease 1 (hEXO1) via a semi-conserved motif. DNA Repair 2012, 11, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Kim, I.K.; Honaker, Y.; Paudyal, S.C.; Koh, W.K.; Sparks, M.; Li, S.; Piwnica-Worms, H.; Ellenberger, T.; You, Z. 14-3-3 proteins restrain the Exo1 nuclease to prevent overresection. J. Biol. Chem. 2015, 290, 12300–12312. [Google Scholar] [CrossRef]
- Ngo, G.H.; Balakrishnan, L.; Dubarry, M.; Campbell, J.L.; Lydall, D. The 9-1-1 checkpoint clamp stimulates DNA resection by Dna2-Sgs1 and Exo1. Nucleic Acids Res. 2014, 42, 10516–10528. [Google Scholar] [CrossRef] [Green Version]
- Ngo, G.H.; Lydall, D. The 9-1-1 checkpoint clamp coordinates resection at DNA double strand breaks. Nucleic Acids Res. 2015, 43, 5017–5032. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zheng, L.; Shen, B. Functional alterations of human exonuclease 1 mutants identified in atypical hereditary nonpolyposis colorectal cancer syndrome. Cancer Res. 2002, 62, 6026–6030. [Google Scholar] [PubMed]
- Hansen, M.F.; Johansen, J.; Bjørnevoll, I.; Sylvander, A.E.; Steinsbekk, K.S.; Sætrom, P.; Sandvik, A.K.; Drabløs, F.; Sjursen, W. A novel POLE mutation associated with cancers of colon, pancreas, ovaries and small intestine. Fam. Cancer 2015, 14, 437–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolenko, A.P.; Preobrazhenskaya, E.V.; Aleksakhina, S.N.; Iyevleva, A.G.; Mitiushkina, N.V.; Zaitseva, O.A.; Yatsuk, O.S.; Tiurin, V.I.; Strelkova, T.N.; Togo, A.V.; et al. Candidate gene analysis of BRCA1/2 mutation-negative high-risk Russian breast cancer patients. Cancer Lett. 2015, 359, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Li, Y.; Hess, K.R.; Abbruzzese, J.L.; Li, D. DNA mismatch repair gene polymorphisms affect survival in pancreatic cancer. Oncologist 2011, 16, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Hanafusa, H.; Ouchida, M.; Yano, M.; Suzuki, H.; Murakami, M.; Aoe, M.; Shimizu, N.; Nakachi, K.; Shimizu, K. Single nucleotide polymorphisms in the EXO1 gene and risk of colorectal cancer in a Japanese population. Carcinogenesis 2005, 26, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Bayram, S.; Akkız, H.; Bekar, A.; Akgöllü, E.; Yıldırım, S. The significance of Exonuclease 1 K589E polymorphism on hepatocellular carcinoma susceptibility in the Turkish population: A case-control study. Mol. Biol. Rep. 2012, 39, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.H.; Tseng, H.C.; Liu, C.S.; Chang, C.L.; Tsai, C.W.; Tsou, Y.A.; Wang, R.F.; Lin, C.C.; Wang, H.C.; Chiu, C.F.; et al. Interaction of Exo1 genotypes and smoking habit in oral cancer in Taiwan. Oral Oncol. 2009, 45, e90–e94. [Google Scholar] [CrossRef]
- Wang, H.C.; Chiu, C.F.; Tsai, R.Y.; Kuo, Y.S.; Chen, H.S.; Wang, R.F.; Tsai, C.W.; Chang, C.H.; Lin, C.C.; Bau, D.T. Association of genetic polymorphisms of EXO1 gene with risk of breast cancer in Taiwan. Anticancer Res. 2009, 29, 3897–3901. [Google Scholar]
- Hsu, N.Y.; Wang, H.C.; Wang, C.H.; Chiu, C.F.; Tseng, H.C.; Liang, S.Y.; Tsai, C.W.; Lin, C.C.; Bau, D.T. Lung cancer susceptibility and genetic polymorphisms of Exo1 gene in Taiwan. Anticancer Res. 2009, 29, 725–730. [Google Scholar]
- Jin, G.; Wang, H.; Hu, Z.; Liu, H.; Sun, W.; Ma, H.; Chen, D.; Miao, R.; Tian, T.; Jin, L.; et al. Potentially functional polymorphisms of EXO1 and risk of lung cancer in a Chinese population: A case-control analysis. Lung Cancer 2008, 60, 340–346. [Google Scholar] [CrossRef]
- Bau, D.T.; Wang, H.C.; Liu, C.S.; Chang, C.L.; Chiang, S.Y.; Wang, R.F.; Tsai, C.W.; Lo, Y.L.; Hsiung, C.A.; Lin, C.C.; et al. Single-nucleotide polymorphism of the Exo1 gene: Association with gastric cancer susceptibility and interaction with smoking in Taiwan. Chin. J. Physiol. 2009, 52, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, D.; Yan, C.; Zhang, L.; Liang, C. Associations between Nine Polymorphisms in EXO1 and Cancer Susceptibility: A Systematic Review and Meta-Analysis of 39 Case-control Studies. Sci. Rep. 2016, 6, 29270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarrola-Villava, M.; Peña-Chilet, M.; Fernandez, L.P.; Aviles, J.A.; Mayor, M.; Martin-Gonzalez, M.; Gomez-Fernandez, C.; Casado, B.; Lazaro, P.; Lluch, A.; et al. Genetic polymorphisms in DNA repair and oxidative stress pathways associated with malignant melanoma susceptibility. Eur. J. Cancer 2011, 47, 2618–2625. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, M.M.; Taleghani, M.Y.; Mohebbi, S.R.; Vahedi, M.; Fatemi, S.R.; Zali, N.; Shemirani, A.I.; Zali, M.R. Impact of EXO1 polymorphism in susceptibility to colorectal cancer. Genet. Test. Mol. Biomarkers 2010, 14, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Alimirzaie, S.; Mohamadkhani, A.; Masoudi, S.; Sellars, E.; Boffetta, P.; Malekzadeh, R.; Akbari, M.R.; Pourshams, A. Mutations in Known and Novel cancer Susceptibility Genes in Young Patients with Pancreatic Cancer. Arch. Iran Med. 2018, 21, 228–233. [Google Scholar] [PubMed]
- Michailidou, K.; Beesley, J.; Lindstrom, S.; Canisius, S.; Dennis, J.; Lush, M.J.; Maranian, M.J.; Bolla, M.K.; Wang, Q.; Shah, M.; et al. Genome-wide association analysis of more than 120,000 individuals identifies 15 new susceptibility loci for breast cancer. Nat. Genet. 2015, 47, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Jiang, R.; Wang, P.; Xu, Y.; Yin, S.; Cheng, X.; Zang, R. Significant association of the EXO1 rs851797 polymorphism with clinical outcome of ovarian cancer. Onco. Targets Ther. 2017, 10, 4841–4851. [Google Scholar] [CrossRef] [PubMed]
- Peltomäki, P.; Vasen, H.F. Mutations predisposing to hereditary nonpolyposis colorectal cancer: Database and results of a collaborative study. The International Collaborative Group on Hereditary Nonpolyposis Colorectal Cancer. Gastroenterology 1997, 113, 1146–1158. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Baitinger, C.; Soderblom, E.J.; Burdett, V.; Modrich, P. Hydrolytic function of Exo1 in mammalian mismatch repair. Nucleic Acids Res. 2014, 42, 7104–7112. [Google Scholar] [CrossRef] [Green Version]
- Bregenhorn, S.; Jiricny, J. Biochemical characterization of a cancer-associated E109K missense variant of human exonuclease 1. Nucleic Acids Res. 2014, 42, 7096–7103. [Google Scholar] [CrossRef] [Green Version]
- Axelsen, J.B.; Lotem, J.; Sachs, L.; Domany, E. Genes overexpressed in different human solid cancers exhibit different tissue-specific expression profiles. Proc. Natl. Acad. Sci. USA 2007, 104, 13122–13127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Tang, Z.; Yang, Z.; Zhang, L.; Deng, Q.; Zhang, X.; Yu, Y.; Liu, X.; Zhu, J. EXO1 overexpression is associated with poor prognosis of hepatocellular carcinoma patients. Cell Cycle 2018, 17, 2386–2397. [Google Scholar] [CrossRef] [PubMed]
- Muthuswami, M.; Ramesh, V.; Banerjee, S.; Viveka Thangaraj, S.; Periasamy, J.; Bhaskar Rao, D.; Barnabas, G.D.; Raghavan, S.; Ganesan, K. Breast tumors with elevated expression of 1q candidate genes confer poor clinical outcome and sensitivity to Ras/PI3K inhibition. PLoS ONE 2013, 8, e77553. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, J.F.; Torrieri, R.; Serafim, R.B.; Di Cristofaro, L.F.; Escanfella, F.D.; Ribeiro, R.; Zanette, D.L.; Paçó-Larson, M.L.; da Silva, W.A., Jr.; Tirapelli, D.P.; et al. Expression of signautes of DNA repair genes correlate with survival prognosis of astrocytomapatients. Tumour. Biol. 2017, 39, 010428317694552. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.S.; Hou, M.F.; Hsieh, Y.C.; Huang, C.Y.; Lee, Y.C.; Chen, Y.J.; Lo, S. Role of Mre11 in cell proliferation, tumor invasion and DNA repair in breast cancer. J. Natl. Cancer Inst. 2012, 104, 1485–1502. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Keymeulen, S.; Nelson, R.; Tong, T.R.; Yuan, Y.C.; Yun, X.; Liu, Z.; Lopez, J.; Raz, D.J.; Kim, J.Y. Overexpression of Flap Endonuclease 1 Correlates with Enhanced Proliferation and Poor Prognosis of Non-Small-Cell Lung Cancer. Am. J. Pathol. 2018, 188, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fatah, T.M.; Russell, R.; Albarakati, N.; Maloney, D.J.; Dorjsuren, D.; Rueda, O.M.; Moseley, P.; Mohan, V.; Sun, H.; Abbotts, R.; et al. Genomic and protein expression analysis reveals flap endonuclease 1 (FEN1) as a key biomarker in breast and ovarian cancer. Mol. Oncol. 2014, 7, 1326–1338. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Repair Process | EXO1 Interaction Proteins in Human | Reference | EXO1 Interaction Proteins in Yeast | Reference |
|---|---|---|---|---|
| Mismatch repair | MSH2 MSH3 MLH1 PCNA | [36,38] [32,33] [38,41] [40] | MSH2 MSH3 MLH1 | [2,34] [72] [72] |
| Homologous recombination /DNA replication/DNA end resection | PARP1 BLM WRN RECQ1 CTIP | [75,76] [57,79] [22] [23] [85] | SGS1 SAE2 | [74] [74] |
| Cell cycle regulation | PCNA 14-3-3η 14-3-3σ | [40,41,95] [97,98] [97,98] | 9-1-1 14-3-3 | [99,100] [96] |
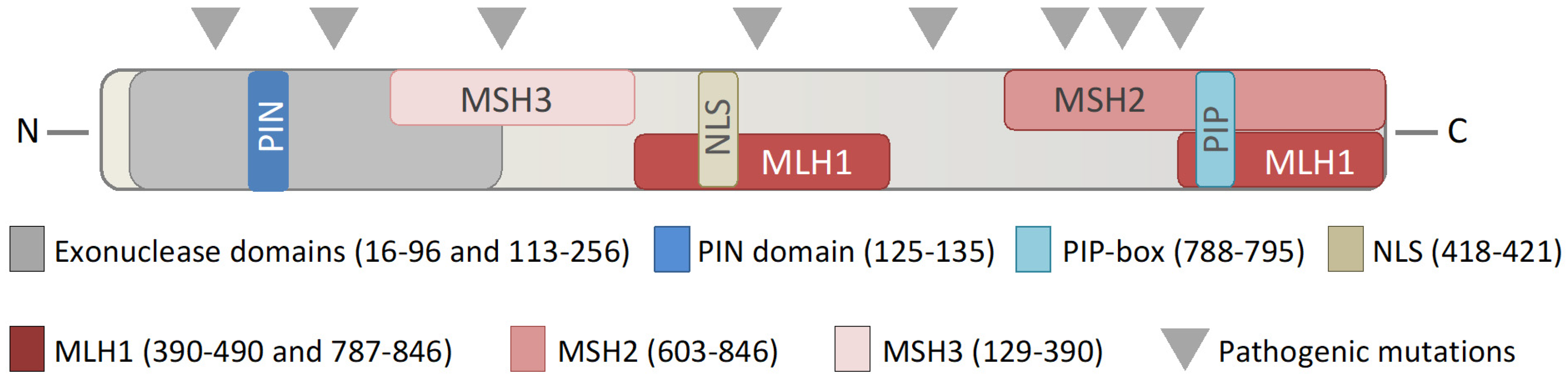
| Mutations in EXO1 Region | Corresponding DNA Sequence Mutation | Reported SNP | Coding and Non-Coding Region | Type of Cancer/Remark | Reference |
|---|---|---|---|---|---|
| p.E109K | c.326A>G | rs756251971 | exon | CRC | [101] |
| p.A153V | c.458C>G | rs143955774 | exon | CRC, IC Combined with polε c.1373A>T, p.Y458F | [102] |
| p.N279S | c.836A>G | rs4149909 | exon | BC, PC | [103,104] |
| p.T439M | c.1317G>A | rs4149963 | exon | CRC | [105] |
| p.E589K | c.1765G>A | rs1047840 | exon | GC, LC, HCC, Melanoma, Glioblastoma | [106,107,108,109,110,111,112,113] |
| p.E670G | c.2009A>G | rs1776148 | exon | GC, BC, OC, LC, Melanoma, Glioblastoma | [106,107,108,109,111,112,113] |
| p.R723G/p.R723S | c.2167C>A/c.2167C>T | rs1635498 | exon | GC, BC, OC, LC | [107,108,109,111,112] |
| p.P757L | c.2270C>T | rs9350 | exon | CRC, PC, GC, OC, LC, BC, Melanoma | [105,107,108,109,111,112,113,114] |
| Non coding region | c.2212-1G>C | rs4150000 | Intron, splicing variant | PC | [115] |
| rs72755295 | Intron, splicing variant | [116] | |||
| rs1776177 | UTR region | GC, BC, OC, LC | [107,108,109,111] | ||
| rs1635517 | UTR region | GC, BC, OC, LC | [107,108,109,111] | ||
| rs3754093 | UTR region | GC, BC, OC, LC | [107,108,109,111] | ||
| rs851797 | UTR region | GC, BC, OC, LC | [107,108,109,111,112,117] | ||
| c.C-908G | rs10802996 | UTR region | CC, GC, BC, OC, LC | [107,108,109,111,112] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keijzers, G.; Bakula, D.; Petr, M.A.; Madsen, N.G.K.; Teklu, A.; Mkrtchyan, G.; Osborne, B.; Scheibye-Knudsen, M. Human Exonuclease 1 (EXO1) Regulatory Functions in DNA Replication with Putative Roles in Cancer. Int. J. Mol. Sci. 2019, 20, 74. https://doi.org/10.3390/ijms20010074
Keijzers G, Bakula D, Petr MA, Madsen NGK, Teklu A, Mkrtchyan G, Osborne B, Scheibye-Knudsen M. Human Exonuclease 1 (EXO1) Regulatory Functions in DNA Replication with Putative Roles in Cancer. International Journal of Molecular Sciences. 2019; 20(1):74. https://doi.org/10.3390/ijms20010074
Chicago/Turabian StyleKeijzers, Guido, Daniela Bakula, Michael Angelo Petr, Nils Gedsig Kirkelund Madsen, Amanuel Teklu, Garik Mkrtchyan, Brenna Osborne, and Morten Scheibye-Knudsen. 2019. "Human Exonuclease 1 (EXO1) Regulatory Functions in DNA Replication with Putative Roles in Cancer" International Journal of Molecular Sciences 20, no. 1: 74. https://doi.org/10.3390/ijms20010074