Thermococcus Eurythermalis Endonuclease IV Can Cleave Various Apurinic/Apyrimidinic Site Analogues in ssDNA and dsDNA


Abstract
:1. Introduction
2. Results
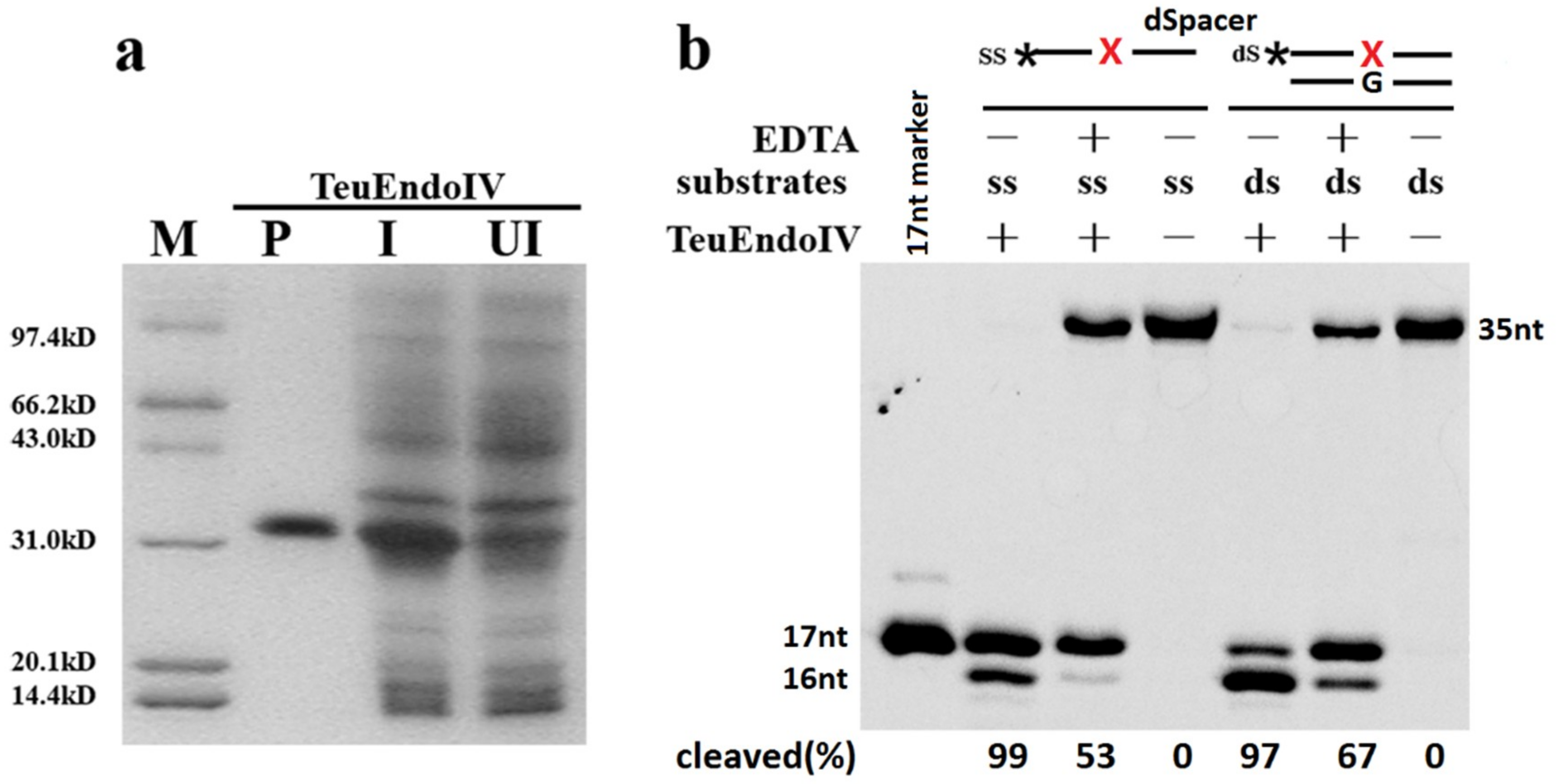
2.1. TeuendoIV Possesses AP (Apurinic/Apyrimidinic) Endonuclease Activity
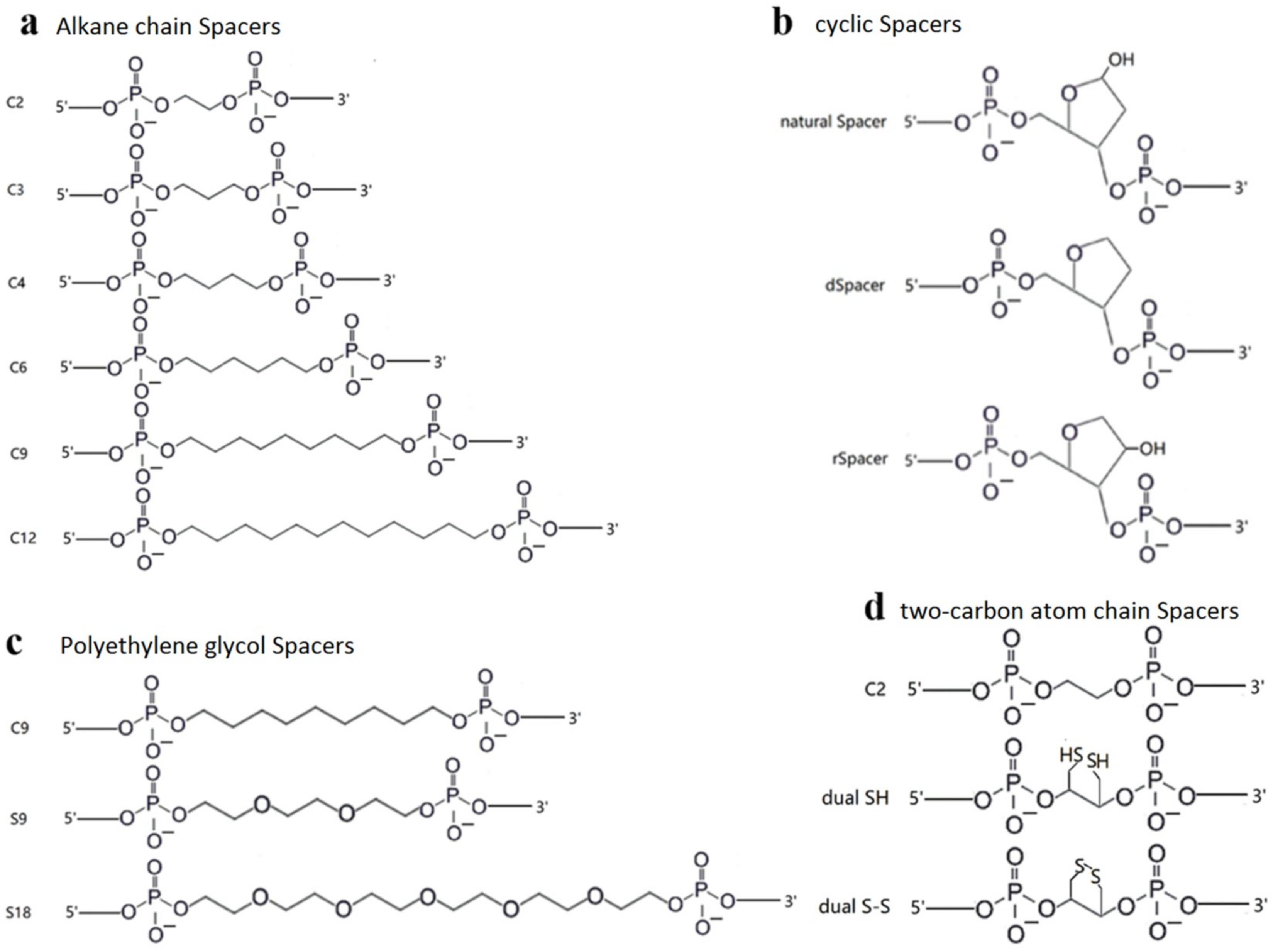
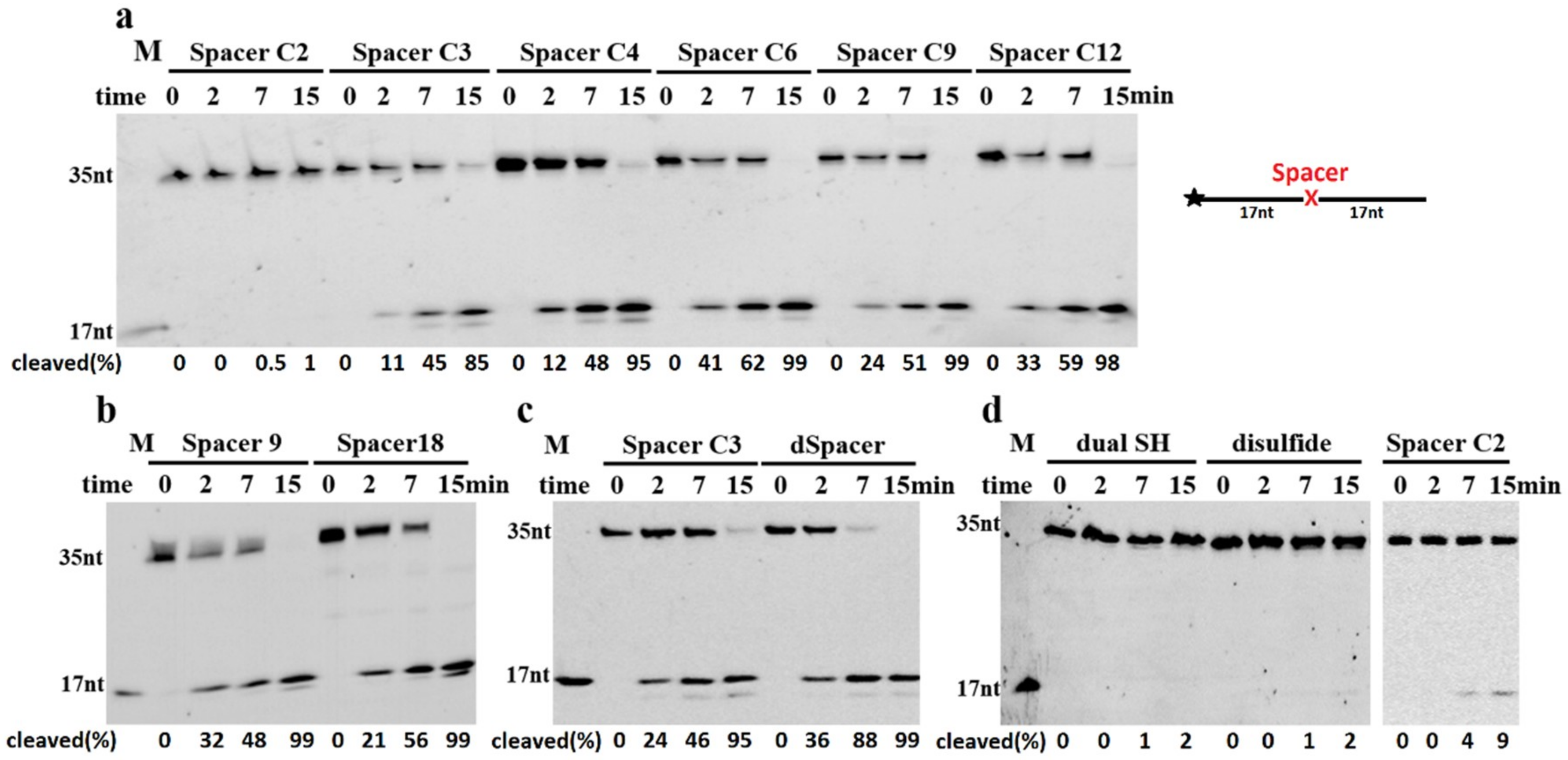
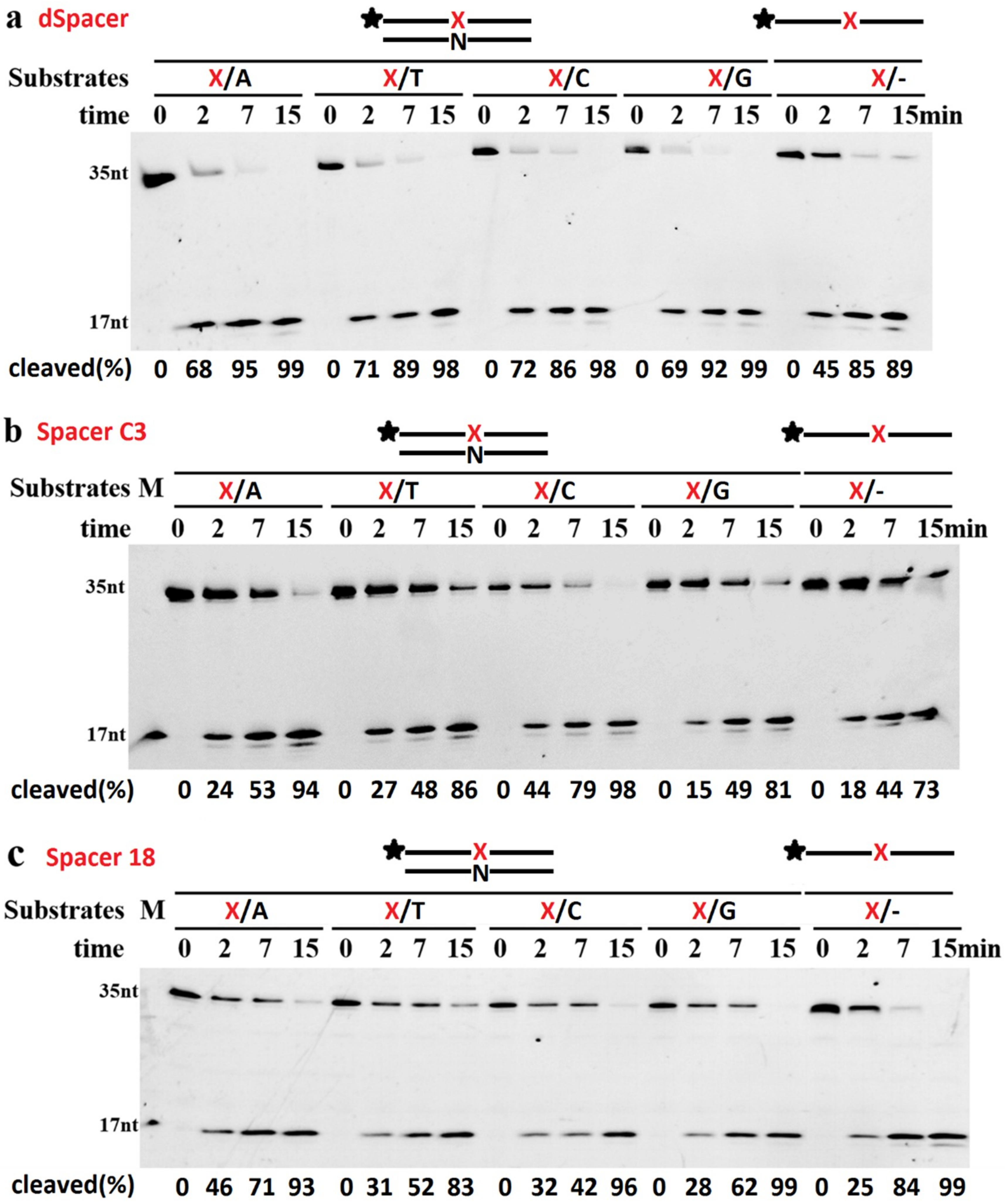
2.2. TeuendoIV Shows AP Endonuclease on Various AP Site Analogues
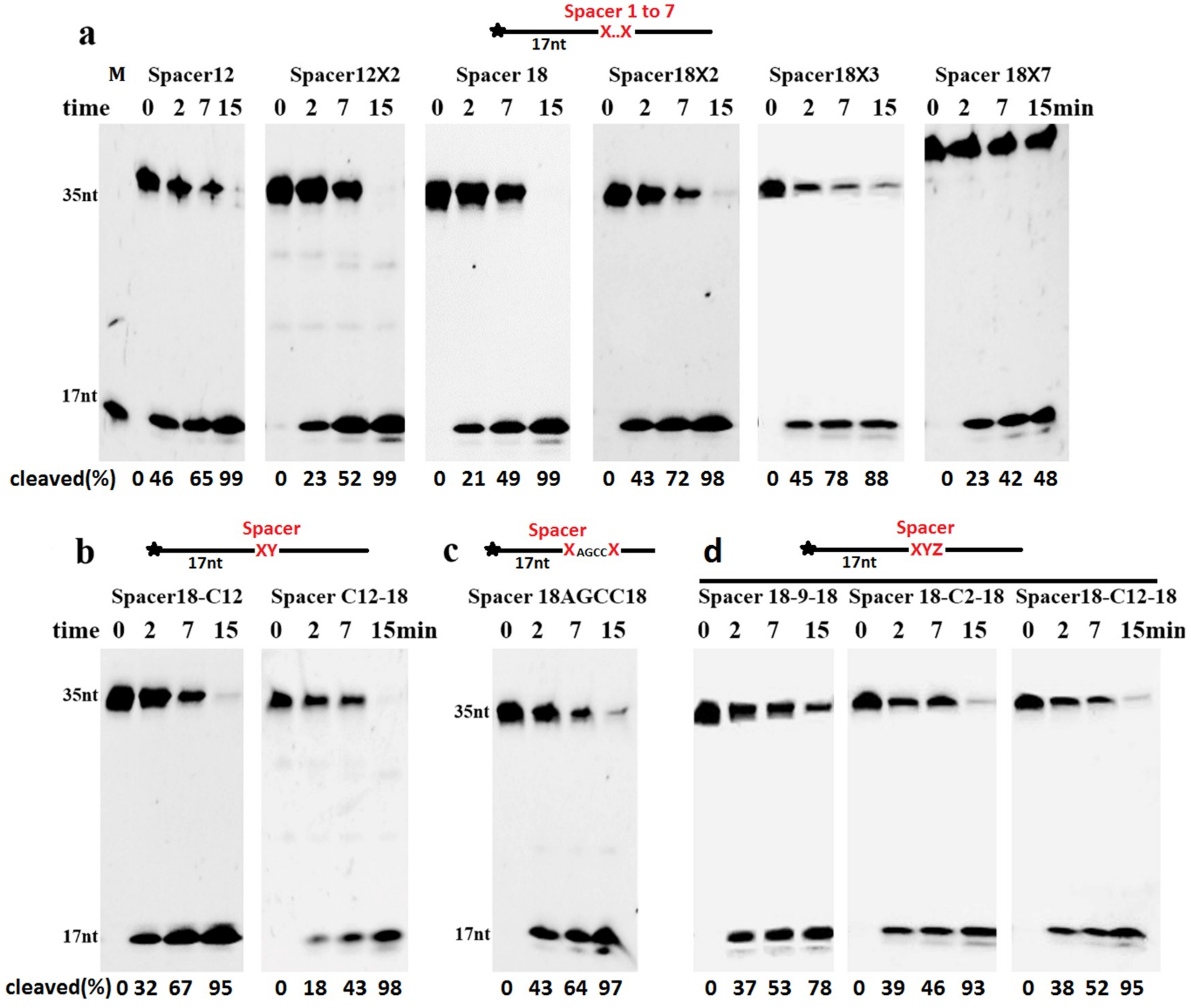
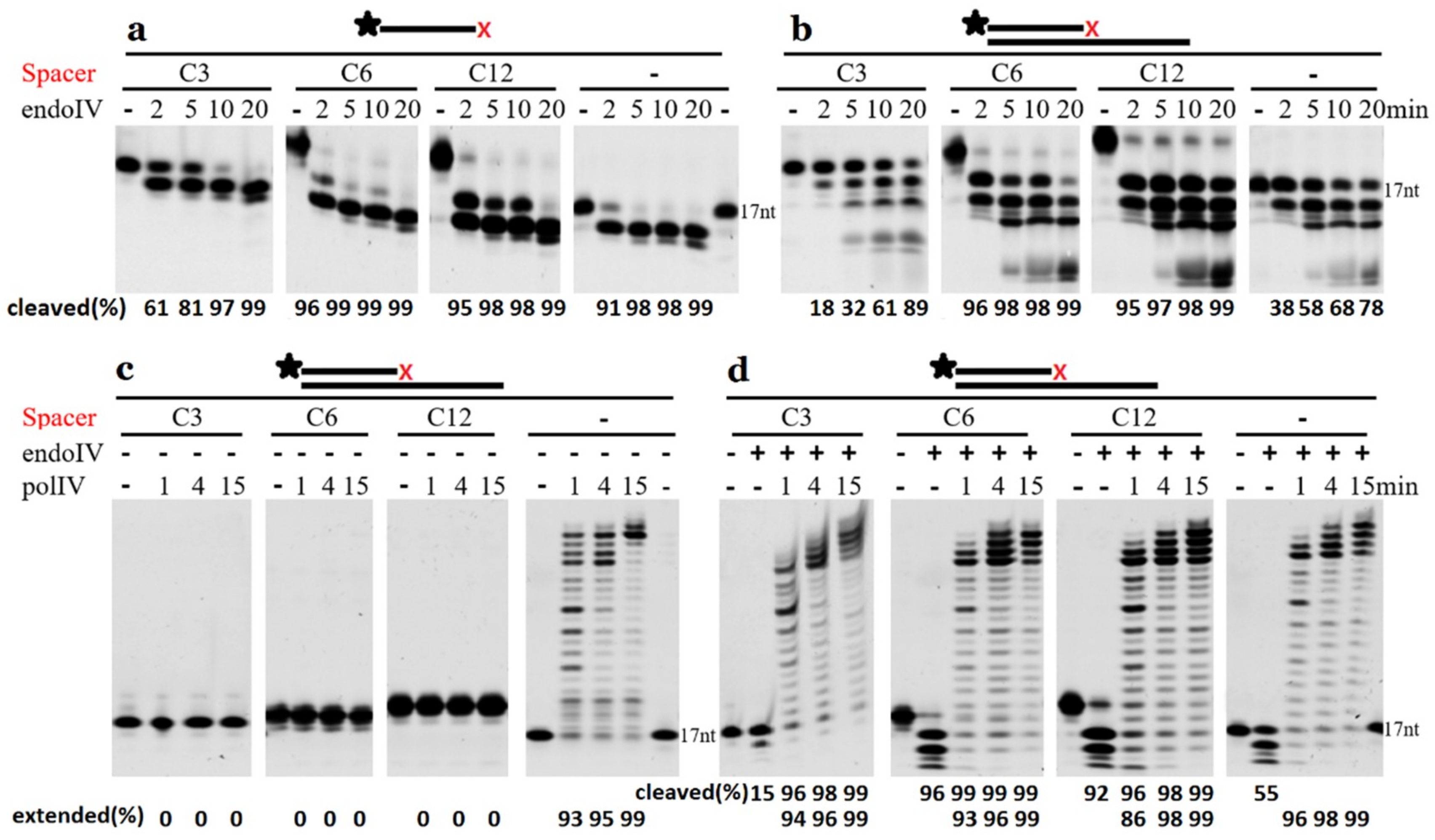
2.3. Cleavage of DNA Containing Clustered AP Site Analogues
2.4. The Base Opposite the AP Site Has Little Effect on dsDNA Cleavage
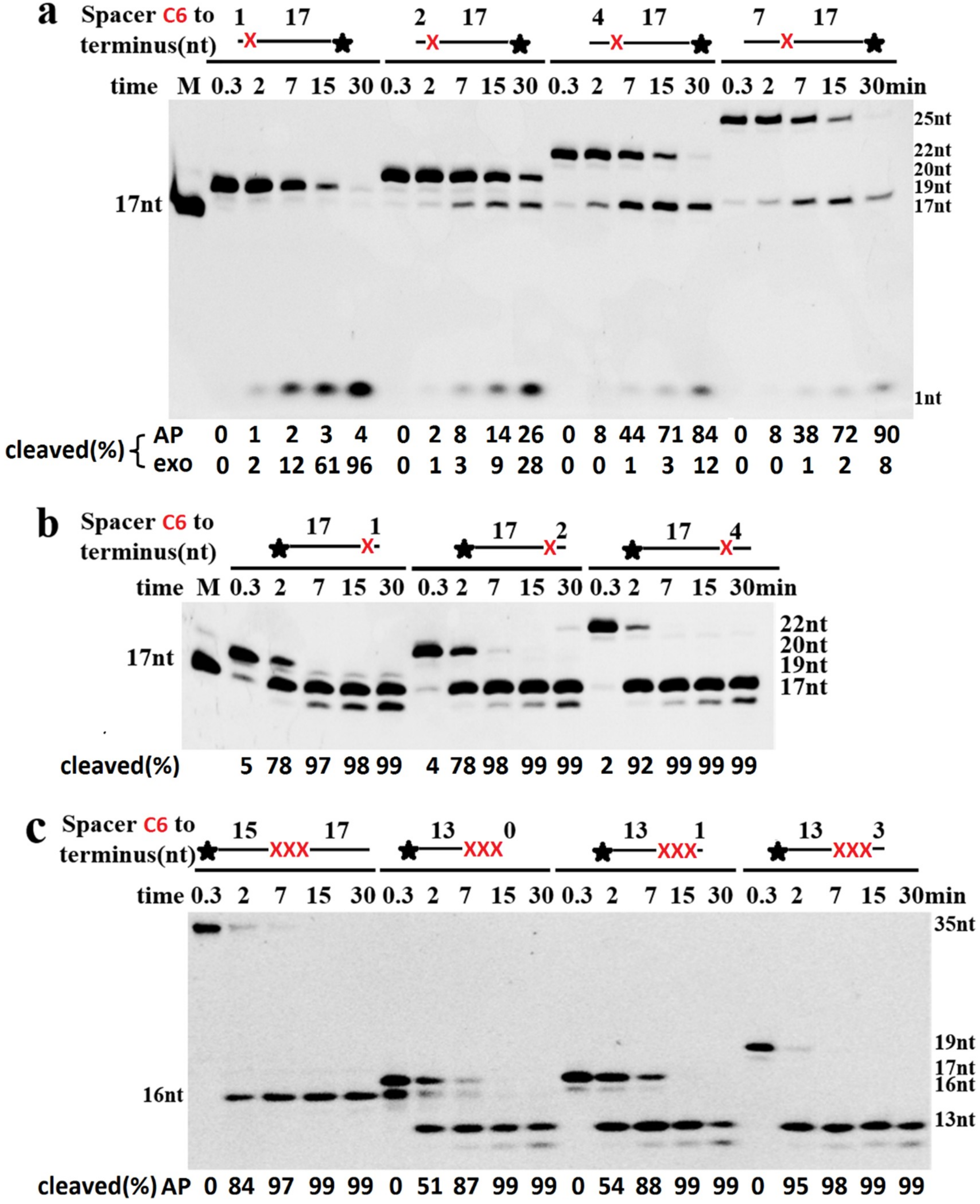
2.5. Cleavage of DNA Containing AP Sites Adjacent to Termini
2.6. Extension by DNA Polymerase Requires the Removal of 3′-Terminal AP Site Analogues
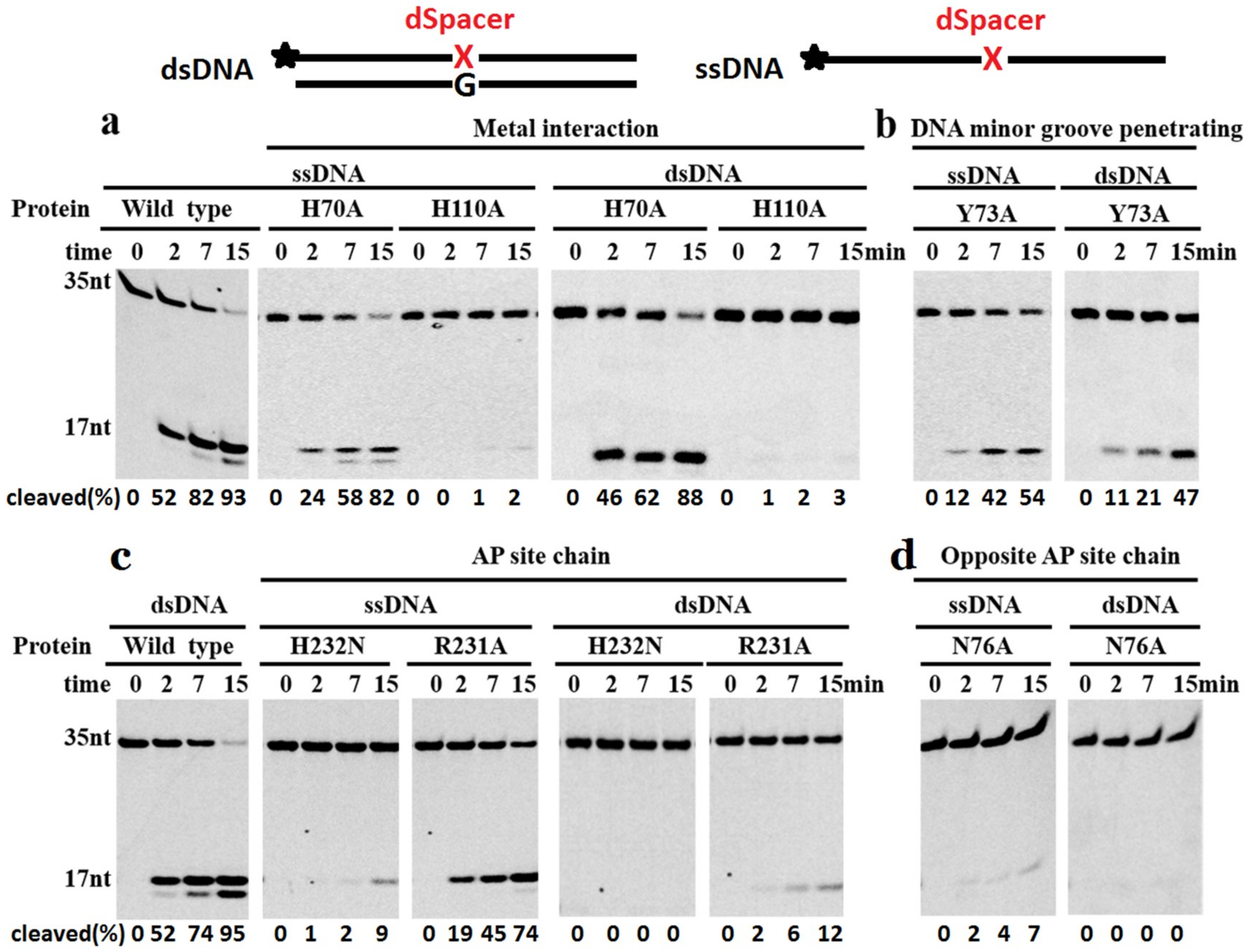
2.7. Key Residues for Recognition and Cleavage of Phosphodiester Bonds
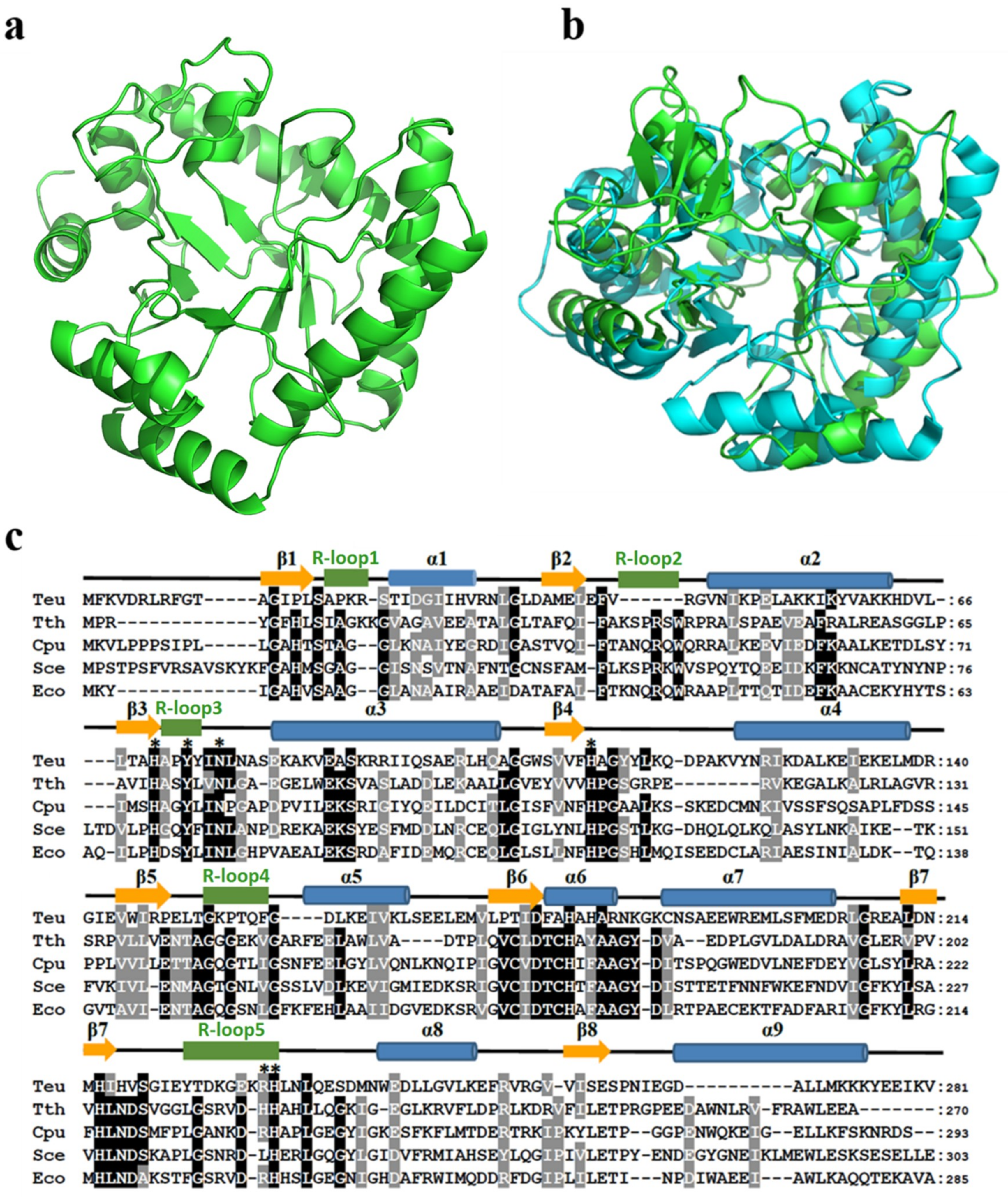
2.8. Structure Comparison of TeuendoIV and EcoendoIV
3. Materials and Methods
3.1. Materials
3.2. Construction of Expression Plasmids
3.3. Expression and Purification of TeuendoIV
3.4. Activity Assay of TeuendoIV
3.5. Multiple Sequence Alignment and Constructing Phylogenetic Tree
3.6. Structure Modeling of EndoIV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lengauer, C.; Kinzler, K.W.; Vogelstein, B. DNA methylation and genetic instability in colorectal cancer cells. Proc. Natl. Acad. Sci. USA 1997, 94, 2545–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Ho, E.; Ames, B.N. Low intracellular zinc induces oxidative DNA damage, disrupts p53, NF kappa B, and AP1 DNA binding, and affects DNA repair in a rat glioma cell line. Proc. Natl. Acad. Sci. USA 2002, 99, 16770–16775. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, R.P. DNA glycosylases. Mutat. Res. 1997, 383, 189–196. [Google Scholar] [CrossRef]
- Lindahl, T.; Nyberg, B. Rate of depurination of native deoxyribonucleic acid. Biochemistry 1972, 11, 3610–3618. [Google Scholar] [CrossRef] [PubMed]
- Cuniasse, P.; Fazakerley, G.V.; Guschlbauer, W.; Kaplan, B.E.; Sowers, L.C. The abasic site as a challenge to DNA polymerase. A nuclear magnetic resonance study of G, C and T opposite a model abasic site. J. Mol. Biol. 1990, 213, 303–314. [Google Scholar] [CrossRef]
- Piersen, C.E.; McCullough, A.K.; Lloyd, R.S. AP lyases and dRPases: Commonality of mechanism. Mutat. Res. 2000, 459, 43–53. [Google Scholar] [CrossRef]
- Erzberger, J.P.; Barsky, D.; Scharer, O.D.; Colvin, M.E.; Wilson, D.M. Elements in abasic site recognition by the major human and Escherichia coli apurinic/apyrimidinic endonucleases. Nucleic Acids Res. 1998, 26, 2771–2778. [Google Scholar] [CrossRef]
- Greenberg, M.M.; Weledji, Y.N.; Kim, J.; Bales, B.C. Repair of oxidized abasic sites by exonuclease III, endonuclease IV, and endonuclease III. Biochemistry 2004, 43, 8178–8183. [Google Scholar] [CrossRef]
- Shida, T.; Noda, M.; Sekiguchi, J. Cleavage of single- and double-stranded DNAs containing an abasic residue by Escherichia coli exonuclease III (AP endonuclease VI). Nucleic Acids Res. 1996, 24, 4572–4576. [Google Scholar] [CrossRef]
- Shida, T.; Ogawa, T.; Ogasawara, N.; Sekiguchi, J. Characterization of Bacillus subtilis ExoA protein: A multifunctional DNA-repair enzyme similar to the Escherichia coli exonuclease III. Biosci. Biotechnol. Biochem. 1999, 63, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Sandigursky, M.; Tainer, J.A.; Franklin, W.A.; Cunningham, R.P. Purification and characterization of Thermotoga maritima endonuclease IV, a thermostable apurinic/apyrimidinic endonuclease and 3′-repair diesterase. J. Bacteriol. 1999, 181, 2834–2839. [Google Scholar] [PubMed]
- Kow, Y.W.; Wallace, S.S. Exonuclease III recognizes urea residues in oxidized DNA. Proc. Natl. Acad. Sci. USA 1985, 82, 8354–8358. [Google Scholar] [CrossRef] [PubMed]
- Hoheisel, J.D. On the activities of Escherichia coli exonuclease III. Anal. Biochem. 1993, 209, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Barnes, T.; Kim, W.C.; Mantha, A.K.; Kim, S.E.; Izumi, T.; Mitra, S.; Lee, C.H. Identification of Apurinic/apyrimidinic endonuclease 1 (APE1) as the endoribonuclease that cleaves c-myc mRNA. Nucleic Acids Res. 2009, 37, 3946–3958. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.C.; Berquist, B.R.; Chohan, M.; Uy, C.; Wilson, D.M., 3rd; Lee, C.H. Characterization of the endoribonuclease active site of human apurinic/apyrimidinic endonuclease 1. J. Mol. Biol. 2011, 411, 960–971. [Google Scholar] [CrossRef] [PubMed]
- Chohan, M.; Mackedenski, S.; Li, W.M.; Lee, C.H. Human apurinic/apyrimidinic endonuclease 1 (APE1) has 3′ RNA phosphatase and 3′ exoribonuclease activities. J. Mol. Biol. 2015, 427, 298–311. [Google Scholar] [CrossRef]
- SKerins, M.; Collins, R.; McCarthy, T.V. Characterization of an endonuclease IV 3′–5′ exonuclease activity. J. Biol. Chem. 2003, 278, 3048–3054. [Google Scholar] [CrossRef]
- Xie, J.J.; Liu, X.P.; Han, Z.; Yuan, H.; Wang, Y.; Hou, J.L.; Liu, J.H. Chlamydophila pneumoniae endonuclease IV prefers to remove mismatched 3′ ribonucleotides: Implication in proofreading mismatched 3′-terminal nucleotides in short-patch repair synthesis. DNA Repair 2013, 12, 140–147. [Google Scholar] [CrossRef]
- Gros, L.; Ishchenko, A.A.; Ide, H.; Elder, R.H.; Saparbaev, M.K. The major human AP endonuclease (Ape1) is involved in the nucleotide incision repair pathway. Nucleic Acids Res. 2004, 32, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Ischenko, A.A.; Saparbaev, M.K. Alternative nucleotide incision repair pathway for oxidative DNA damage. Nature 2002, 415, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Golan, G.; Ishchenko, A.A.; Khassenov, B.; Shoham, G.; Saparbaev, M.K. Coupling of the nucleotide incision and 3′->5′ exonuclease activities in Escherichia coli endonuclease IV: Structural and genetic evidences. Mutat. Res. 2010, 685, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.; Weiss, B. Endonuclease IV of Escherichia coli is induced by paraquat. Proc. Natl. Acad. Sci. USA 1987, 84, 3189–3193. [Google Scholar] [CrossRef] [PubMed]
- Demple, B.; Herman, T.; Chen, D.S. Cloning and expression of APE, the cDNA encoding the major human apurinic endonuclease: Definition of a family of DNA repair enzymes. Proc. Natl. Acad. Sci. USA 1991, 88, 11450–11454. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.W.; Demple, B. Yeast DNA 3′-repair diesterase is the major cellular apurinic/apyrimidinic endonuclease: Substrate specificity and kinetics. J. Biol. Chem. 1988, 263, 18017–18022. [Google Scholar] [PubMed]
- Hosfield, D.J.; Guan, Y.; Haas, B.J.; Cunningham, R.P.; Tainer, J.A. Structure of the DNA repair enzyme endonuclease IV and its DNA complex: Double-nucleotide flipping at abasic sites and three-metal-ion catalysis. Cell 1999, 98, 397–408. [Google Scholar] [CrossRef]
- Garcin, E.D.; Hosfield, D.J.; Desai, S.A.; Haas, B.J.; Björas, M.; Cunningham, R.P.; Tainer, J.A. DNA apurinicapyrimidinic site binding and excision by endonuclease IV. Nat. Struct. Mol. Biol. 2008, 15, 515–522. [Google Scholar] [CrossRef]
- Ivanov, I.; Tainer, J.A.; McCammon, J.A. Unraveling the three-metal-ion catalytic mechanism of the DNA repair enzyme endonuclease IV. Proc. Natl. Acad. Sci. USA 2007, 104, 1465–1470. [Google Scholar] [CrossRef]
- Asano, R.; Ishikawa, H.; Nakane, S.; Nakagawa, N.; Kuramitsu, S.; Masui, R. An additional C-terminal loop in endonuclease IV, an apurinic/apyrimidinic endonuclease, controls binding affinity to DNA. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 149–155. [Google Scholar] [CrossRef]
- Mol, C.D.; Izumi, T.; Mitra, S.; Tainer, J.A. DNA-bound structures and mutants reveal abasic DNA binding by APE1 and DNA repair coordination. Nature 2000, 403, 451–456. [Google Scholar] [CrossRef]
- Freudenthal, B.D.; Beard, W.A.; Cuneo, M.J.; Dyrkheeva, N.S.; Wilson, S.H. Capturing snapshots of APE1 processing DNA damage. Nat. Struct. Mol. Biol. 2015, 22, 924–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mol, C.D.; Kuo, C.F.; Thayer, M.M.; Cunningham, R.P.; Tainer, J.A. Structure and function of the multifunctional DNA-repair enzyme exonuclease III. Nature 1995, 374, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Schmiedel, R.; Kuettner, E.B.; Keim, A.; Strater, N.; Greiner-Stoffele, T. Structure and function of the abasic site specificity pocket of an AP endonuclease from Archaeoglobus fulgidus. DNA Repair 2009, 8, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Nyberg, B. Heat-induced deamination of cytosine residues in deoxyribonucleic acid. Biochemistry 1974, 13, 3405–3410. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Xiao, X. Complete genome sequence of Thermococcus eurythermalis A501, a conditional piezophilic hyperthermophilic archaeon with a wide temperature range, isolated from an oil-immersed deep-sea hydrothermal chimney on Guaymas Basin. J. Biotechnol. 2015, 193, 14–15. [Google Scholar] [CrossRef] [PubMed]
- Kiyonari, S.; Tahara, S.; Shirai, T.; Iwai, S.; Ishino, S.; Ishino, Y. Biochemical properties and base excision repair complex formation of apurinic/apyrimidinic endonuclease from Pyrococcus furiosus. Nucleic Acids Res. 2009, 37, 6439–6453. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.P.; Liu, J.H. Chlamydia pneumoniae AP endonuclease IV could cleave AP sites of double- and single-stranded DNA. Biochim. Biophys. Acta 2005, 1753, 217–225. [Google Scholar] [CrossRef]
- Marenstein, D.R.; Wilson, D.M., 3rd; Teebor, G.W. Human AP endonuclease (APE1) demonstrates endonucleolytic activity against AP sites in single-stranded DNA. DNA Repair 2004, 3, 527–533. [Google Scholar] [CrossRef]
- Wang, W.W.; Xie, J.J.; Zhou, H.; Liu, X.P. The function of N-terminal peptide for endoIV during cleavage of AP site. Gene. in preparation.
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, 252–258. [Google Scholar] [CrossRef]
- Georg, J.; Schomacher, L.; Chong, J.P.; Majerník, A.I.; Raabe, M.; Urlaub, H.; Müller, S.; Ciirdaeva, E.; Kramer, W.; Fritz, H.J. The Methanothermobacter thermautotrophicus ExoIII homologue Mth212 is a DNA uridine endonuclease. Nucleic Acids Res. 2006, 34, 5325–5336. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.P.; Corbett, A.; Thomson, H.; Adacha, J.; Jensen, K.; Bergeron, J.; Kasampalidis, I.; Exley, R.; Winterbotham, M.; Tang, C.; et al. AP endonuclease paralogues with distinct activities in DNA repair and bacterial pathogenesis. EMBO J. 2007, 26, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.P.; Zhang, Y.; Liang, R.B.; Hou, J.L.; Liu, J.H. Characterization of the 3′ exonuclease of Chlamydophila pneumoniae endonuclease IV on double-stranded DNA and the RNA strand of RNA/DNA hybrid. Biochem. Biophys. Res. Commun. 2007, 361, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Ishchenko, A.A.; Ide, H.; Ramotar, D.; Nevinsky, G.; Saparbaev, M. α-Anomeric deoxynucleotides, anoxic products of ionizing radiation, are substrates for the endonuclease IV-type AP endonucleases. Biochemistry 2004, 43, 15210–15216. [Google Scholar] [CrossRef] [PubMed]
- Ide, H.; Tedzuka, K.; Shimzu, H.; Kimura, Y.; Purmal, A.A.; Wallace, S.S.; Kow, Y.W. α-Deoxyadenosine, a major anoxic radiolysis product of adenine in DNA, is a substrate for Escherichia coli endonuclease IV. Biochemistry 1994, 33, 7842–7847. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, M.C.; Balachander, S.; Antoniali, G.; Koh, K.D.; Saint-Pierre, C.; Gasparutto, D.; Chon, H.; Crouch, R.J.; Storici, F.; Tell, G. Abasic and oxidized ribonucleotides embedded in DNA are processed by human APE1 and not by RNase H2. Nucleic Acids Res. 2017, 45, 11193–11212. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrates | Km (μM) | kcat (min−1) | kcat/Km (min−1·μM−1) |
|---|---|---|---|
| dSpacer | 0.35 ± 0.03 | 0.59 ± 0.05 | 1.68 ± 0.15 |
| Spacer C2 | 0.16 ± 0.02 | 0.0021 ± 0.0002 | 0.013 ± 0.002 |
| Spacer C3 | 0.37 ± 0.03 | 0.23 ± 0.02 | 0.62 ± 0.06 |
| Spacer C4 | 0.41 ± 0.04 | 0.33 ± 0.03 | 0.80 ± 0.08 |
| Spacer C6 | 0.36 ± 0.03 | 0.37 ± 0.04 | 1.03 ± 0.10 |
| Spacer C12 | 0.41 ± 0.04 | 0.35 ± 0.03 | 0.85 ± 0.08 |
| Spacer 9 | 0.34 ± 0.03 | 0.36 ± 0.03 | 1.06 ± 0.10 |
| Spacer 18 | 0.37 ± 0.04 | 0.42 ± 0.04 | 1.14 ± 0.11 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.-W.; Zhou, H.; Xie, J.-J.; Yi, G.-S.; He, J.-H.; Wang, F.-P.; Xiao, X.; Liu, X.-P. Thermococcus Eurythermalis Endonuclease IV Can Cleave Various Apurinic/Apyrimidinic Site Analogues in ssDNA and dsDNA. Int. J. Mol. Sci. 2019, 20, 69. https://doi.org/10.3390/ijms20010069
Wang W-W, Zhou H, Xie J-J, Yi G-S, He J-H, Wang F-P, Xiao X, Liu X-P. Thermococcus Eurythermalis Endonuclease IV Can Cleave Various Apurinic/Apyrimidinic Site Analogues in ssDNA and dsDNA. International Journal of Molecular Sciences. 2019; 20(1):69. https://doi.org/10.3390/ijms20010069
Chicago/Turabian StyleWang, Wei-Wei, Huan Zhou, Juan-Juan Xie, Gang-Shun Yi, Jian-Hua He, Feng-Ping Wang, Xiang Xiao, and Xi-Peng Liu. 2019. "Thermococcus Eurythermalis Endonuclease IV Can Cleave Various Apurinic/Apyrimidinic Site Analogues in ssDNA and dsDNA" International Journal of Molecular Sciences 20, no. 1: 69. https://doi.org/10.3390/ijms20010069