The Formation and Therapeutic Update of Tumor-Associated Macrophages in Cervical Cancer


1
Department of Obstetrics and Gynecology, University Hospital, LMU Munich, 80337 Munich, Germany
2
Klinik für Innere Medizin I, Technische Universität München, 80333 Munich, Germany
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(13), 3310; https://doi.org/10.3390/ijms20133310
Submission received: 7 May 2019
/
Revised: 1 July 2019
/
Accepted: 3 July 2019
/
Published: 5 July 2019
(This article belongs to the Special Issue Molecular Research on Cervical Cancer)
Abstract
:Both clinicopathological and experimental studies have suggested that tumor-associated macrophages (TAMs) play a key role in cervical cancer progression and are associated with poor prognosis in the respects of tumor cell proliferation, invasion, angiogenesis, and immunosuppression. Therefore, having a clear understanding of TAMs is essential in treating this disease. In this review, we will discuss the origins and categories of macrophages, the molecules responsible for forming and reeducating TAMs in cervical cancer (CC), the biomarkers of macrophages and the therapy development targeting TAMs in CC research.
1. Introduction
Although vaccinations, radical operations, and radiochemotherapy have been widely used for preventing and treating cervical cancer (CC), there are still over half a million new cases worldwide annually. CC is the fourth most common cause of death from cancer among women [1]. Persistent infection of human papillomavirus (mainly HPV16 and HPV18) in the cervix has been established as the primary cause of cervical cancer [2]. E6 and E7, as HPV oncogenes, contribute to neoplastic progression when combined with the host cervical cell genome [3]. Drug resistance, recurrence, and metastasis are the primary culprits for increased mortality, which are also associated with the tumor microenvironment (TME). The TME is mainly compromised of tumor cells, bone-marrow-derived cells and host stromal cells. Their components interact to offer an immunosuppressive environment, thus facilitating tumor progression [4]. Among these components, the role of tumor-associated macrophages (TAMs) has been receiving increasing consideration. TAMs belong to a subunit of macrophages that are recruited to tumor tissue and affect tumor growth [5]. Dr. Günther pioneered the macrophage electrophoresis mobility test (MEM test) in the diagnosis of malignant gynecological diseases [6]. As a result, the function of TAMs in CC has attracted more and more attention in recent years [7]. In this review, we will discuss the origins and categories of macrophages, the molecules involved in forming and reeducating TAMs in CC, the biomarkers of macrophages and the therapy development targeting TAMs in CC research.
2. The Origins and Categories of Macrophages
There are two kinds of macrophages involved in the pathophysiology of cervical cancer: tissue-resident macrophages and infiltrating macrophages (Table 1).
2.1. Tissue-Resident Macrophages
During embryonic organogenesis, macrophages derived from yolk sac and fetal liver precursors are seeded in or on the epithelia throughout normal, non-injured or inflamed tissues. These macrophages persist into adulthood as resident, self-maintaining populations [8]. They generally show specialized properties and functions that are related to their respective tissue locations, such as Kupffer cells in the liver, alveolar macrophages in the lungs, red pulp macrophages in the spleen and microglia in the brain [12,13]. After birth, bone marrow or spleen-derived monocytes can replenish tissue resident macrophages following injury, infection or inflammation. These macrophages reside in interstitial locations within the tissue. For example, the replacement of alveolar macrophages by postnatal monocytes yielded new macrophages that had an expression profile that was almost identical to the original embryonically derived alveolar macrophages. This suggests an overarching role of the tissue environment in dictating overall macrophage function [9]. Tissue-resident macrophage maintenance and homeostasis depend on its origin in the steady state, which can be disturbed by pathological or physiological inflammation [8,14]. For example, small doses of human seminal plasma inhibited lymphocyte transformation, which could possibly be mediated through accessory macrophages. Investigators also revealed that seminal plasma interferes with the attachment, spreading and phagocytic activity of C. parvum elicited murine peritoneal macrophages [15]. Moreover, treatment with Sizofiran (SPG) and recombinant interferon γ (IFN-γ) could make peritoneal macrophages obtained from CC acquire new potent tumoricidal activity along with the increasing secretion of TNF, IFN-γ, and IL-1 and inhibiting the increased secretion of PGE2 [16].
2.2. Infiltrating Macrophages
Many pro-inflammatory mediators recruit circulating monocytes to tumor, inflammatory or infected tissue and acquire the properties of infiltrating macrophages. Along with the inflammatory stage, inflammatory macrophages present mainly two types of properties, ‘pro-inflammatory’ versus ‘pro-resolving’, ‘classically activated’ versus ‘alternatively activated’ or ‘M1-like’ versus ‘M2-like’. These properties are under the influence of growth factors, metabolic requirements, local oxygen tension, the tissue cells and the tissue matrix [10]. In fact, monocyte-derived macrophages in the tissue are often highly heterogeneous as they go through the various influencing factors. For example, four subsets of M2 were identified, including M2a, M2b, M2c, and M2d [17].
M1-like polarized macrophages are induced by Th1 cytokines IFN γ, IL-2, IL-3, IL-12, TNF-α, bacterial constituent such as lipopolysaccharide (LPS), and Toll-like receptor (TLR) agonists. M1-like macrophages secret pro-inflammatory factors such as IL-1 beta, IL-6, IL-12, IL-23, iNOS and tumor necrosis factor-α (TNF-α), chemokine ligands 9 (CXCL-9), CXCL-10 and express the major histocompatibility complex class I (MHC I) and class II (MHC II) molecules. Thus, the M1-like macrophages act in the inflammatory response and antitumor immunity. Oppositely, the M2-like macrophages are induced by Th2 cytokines such as IL-4, IL-10, IL-13, TNF-α, TGF-β, GM-CSF, immune complexes and TLRs. M2-like macrophages secret a wide array of anti-inflammatory molecules, such as IL-10, TGF-β, and arginase 1 and exert anti-inflammatory and pro-tumorigenic activities [11,18]. TAMs could promote the proliferation, invasion, and metastasis of cervical cancer cells. In addition, TAMs could participate in tissue remodeling, fibrosis and angiogenesis in cervical cancer. Recent studies show TAMs exhibit functions similar to those of M2-like macrophages [19].
3. The Molecules Involved in Forming TAMs
In cervical cancer, the mechanism involved in forming TAMs mainly relate to tumor-derived molecules, T cell-derived molecules, seminal plasma-derived molecules, the anaerobic microenvironment and molecules from other sources.
3.1. Tumor-Derived Molecules
Multiple malignant tumor cell-derived molecules could promote TAM formation. For example, the conditioned medium of pancreatic cells, which contained secreted REG4, could induce the polarization of macrophages to an M2-like phenotype [20]. Prostate cancer cells secreted CCN3, which could recruit macrophages and skew their differentiation to an M2-like phenotype [21]. Meanwhile, the supernatant of colorectal cancer cells induced the M2-like polarization of macrophages [22]. Finally, both senescent thyrocytes and thyroid tumor cell lines triggered M2-like macrophage polarization that was related to PGE2 secretion [23].
As to the formation of TAMs in cervical cancer, in 1982 investigators noted that monocyte functions of chemotaxis, phagocytosis and helper function on T-cell mitogen response were significantly inhibited when monocytes were pre-incubated in cervical cancer sera. It was speculated that some components in cancer sera played an immuno-suppressive role through the inhibition of monocyte functions in cancer-bearing state [24]. It was further discovered that when treated with the supernatant of CC cell lines, M1-like macrophages developed an M2-like phenotype with increased CD163, TLR-3, -7, -9 and IL-10 [25,26]. Similarly, co-culture cervical cancer cells decrease the macrophage M1-like polarization partly through necroptosis downregulation [27]. However, these studies did not clarify which component of the supernatant was responsible for the M2-like induction. With the development of research, investigators have found that CC cell-derived molecules act in the polarization and activation of monocytes (Table 2).
3.1.1. Molecules Promoting the Differentiation of Monocytes to TAMs
PGE2 and IL-6 produced by CC cells could cause M2-like phenotype differentiation. Furthermore, these M2-like macrophages displayed a lower expression of costimulatory molecules, an altered balance in IL-12p70 and IL-10 production, and a poor capacity to stimulate T-cell proliferation and IFN-γ production after having been activated by TLR-agonists. This creates a tolerogenic tumor microenvironment [28].
Karyopherin β1 is a nuclear import protein involved in the transport of proteins containing a nuclear localization sequence. Transcriptional factors such as NFκB and AP-1 initiate the expression of multiple factors associated with inflammation and cancer cell biology. Inhibition of Karyopherin β1 in HeLa cells led to declined transcriptional activity of NFκB and AP-1 and reduced migration and invasion of cervical cancer cells [29]. In addition, a study showed that E6 stimulated the expression of multiple genes known to be inducible by NFkB and AP-1 [38], such as COX-2 [39]. The onocogene E6 enhanced expression of functional components of the NFκB signal pathway, including p50, NIK, and TRAF-interacting protein, and increased the binding of NFκB and AP-1 to DNA consensus binding sites [38]. Also, Th2 cell- secreting cytokines such as interleukin-6, interleukin-1β, TNFα, and GM-CSF are also the targeted genes of NFκB and AP-1 [29]. It was further found that Th2 cytokines induce M2-like macrophages. [18]. These results indicated that Karyopherin β1 may participate in inducing M2-like macrophage differentiation, but the existing studies have lacked direct evidence to show the effect of Karyopherin β1 on macrophage polarization.
The chemokine CCL2 (MCP-1) combines with the receptors on monocytes and is important for monocyte recruitment and modulation of macrophages towards the M2-like phenotype. CCL2 was found to determine the extent of macrophage polarization because CCL2 enhanced the LPS-induced production of IL-10, whereas the blockade of CCL2 led to enhanced expression of M1-like polarization-associated genes and cytokines, and diminished expression of M2-like-associated markers in human macrophages [28,29,30,38]. The CCL2 mRNA expression level in CC cells was positively associated with the number of TAMs. Lack of CCL2 mRNA was associated with increased cumulative relapse-free survival, cumulative overall survival, less post-operative surgery, reduced local and distant recurrence, reduced vascular invasion and smaller tumor size [40].
IL-10 is produced by multiple cells, including HPV transformed CC cells [41]. IL-10 causes contradictory effects on CC. It has been found that high IL-10 levels may prevent cervical neoplasia by assisting HPV elimination [42]. Conversely, low IL-10 levels are associated with a higher risk for cervical cancer [43]. It could be that IL-10 induces monocytes to the M2c subtype [44], while also inhibiting MHC I and MHC II expression and thus impairing their ability to present antigens. [45,46,47]. Other research has shown that IL-10 inhibited the classic activation of macrophages by JAK1/Tyk2/STAT3 pathway, which was the most commonly activated pathway by IL-10 [31].
A variant of prolactin (PRL) is produced by cervical cancer cells. It reduced apoptosis in HeLa, SiHa and C-33A cell lines and induced IL-1β and TNF-α production by human peripheral blood mononuclear cell line THP-1 macrophages [32]. Higher doses of PRL (1000 ng/mL) induces macrophages to produce anti-inflammatory cytokine IL-10, which leads to a significant decrease in production of proinflammatory cytokines [33]. These results indicated that PRL might participate in the polarization of monocytes towards the M2-2 phenotype.
3.1.2. Molecules Promoting the Activation of TAMs
α-l-Fucosidase (FUCA-1) activity was found to be increased in CC tissue. FUCA-1 is a glycosidase, which splits terminal α-l-fucose from glycoproteins [48]. FUCA-1 was discovered to be expressed in thyroid carcinoma cell lines. However, few research papers have reported its expression in CC cells. After incubation of human monocyte-derived macrophages with FUCA-1, the macrophages were no longer able to respond to lipopolysaccharide (LPS). In addition, after tumor cells were treated with FUCA-1 and then co-cultivated with macrophages, macrophage-mediated cytotoxicity had no effect on tumor cells [34]. These studies indicate that FUCA-1 may be over-expressed in CC cells and might promote the activation of TAMs by inhibiting antigen presentation function and phagocytosis.
Cervical cancer progression is associated with increased serum levels of granulocyte macrophage colony-stimulating factor (GM-CSF) [49]. The expression levels of GM-CSF produced by CC cells were positively correlated with the number of TAMs [50]. CC cells release GM-CSF, which in turn activates mononuclear phagocytes and induces them to release HB-EGF. HB-EGF then triggers anti-apoptotic and proliferative signals in CC cells, which leads to the release of more GM-CSF [35]. These studies indicate that GM-CSF could activate TAMs to release pro-tumor factors.
Macrophage migration inhibitory factor (MIF) expression was significantly increased in cervical cancer samples. SiHa and CaSki cervical cancer cells were discovered to secret soluble MIF into cell culture supernatants [51]. CD74, a receptor for MIF, has been known to be expressed on HLA Class II-positive normal cells including macrophages [52]. The genetic polymorphism MIF-173 is associated with cervical cancer in humans. Patients with the CC genotype exhibited higher MIF serum concentration, which could increase the risk of early stage cervical cancer and lymphatic metastasis [53]. Deep stromal infiltration correlated with the overexpression of MIF in cervical intraepithelial neoplasia (CIN) and squamous cell carcinoma (SCC). Both protein levels of MIF and CD74 were also associated with microvessel density [54]. What is more, long-term loss of MIF significantly inhibited growth and proliferation of HeLa cells while increasing HeLa cell adhesion and therefore impairing their migratory capacity [55]. In melanomas, MIF deficiency attenuated the alternative activation of tumor-polarized macrophages, immunosuppression, and neoangiogenesis [36]. These studies indicate that MIF may participate in the activation of M2-like macrophages in CC.
IL-10 is produced by multiple cells, including HPV transformed CC cells [41] and TAMs [37]. In tuberculosis, macrophage-derived IL-10 could promote M2-like activation by elevating the expression of IL-4Rα as well as IL-4R-dependent arginase 1 [37]. Moreover, IL-10 could promote the expression of arginase with LPS so that the macrophages could exert the ability of immunosuppression [56,57]. These studies implicate that IL-10 from CC cells may participate in the alternative activation of macrophages.
3.1.3. The role of Oncoproteins on TAMs Formation in CC
The HPV genome contains eight open reading frames (ORF) divided into two regions, the early (E) and the late (L) regions. The E-region encodes for six genes (E1, E2, E4, E5, E6, and E7) [58]. Generally, E1, E2, and E4 proteins are responsible for viral amplification and release [59,60,61]. E5, E6 and E7 proteins show tumor-promoting activities, with E6 and E7 corresponding to the primary transformation of viral proteins [61,62,63]. Oncoproteins could promote the production of molecules which participate in TAM formation in CC (Figure 1).
Accumulated data has shown that after HPV infection, IL-10 levels increase because HPV E2, E6, and E7 proteins act in IL10 gene transcription, while IL-10 stimulates HPV E6 and E7 expression [65]. Moreover, IL-10 participates in the differentiation of cells into M2-like macrophages. M2 macrophages were further characterized by expressing high levels of IL-10 as stated above [37,45,46,47,69]. Therefore, the interaction between HPV oncoproteins and IL-10 creates a vicious cycle that could favor an immunosuppressive microenvironment in the cervix.
The HPV-16 E6 protein stimulated the expression of multiple genes known to be inducible by NF-κB and AP-1. What is more, E6 enhanced expression of functional components within the NF-κB signal pathway [38]. The E6-dependent c-fos oncogenic protein expression contributes as well to AP-1 complex formation under oxidative stress in SiHa cells [70]. This leads to the idea that downstream targeted genes of NF-κB and AP-1 might participate in the differentiation towards the M2-like phenotype.
While several studies have shown that CCL2 participates in the recruitment of monocytes and promotes the polarization of monocytes into the M2-like phenotype, it should also be noted that the expression of CCL2 in CC cells was individualized [28,29,38,39]. Meanwhile, other studies have proven that the HPV oncogenes E6 or E7 inversely correlate with the expression of the MCP-1 gene [69], which means E6 or E7 could inhibit the CCL2s ability to recruitment monocytes. There may, however, be other molecules in CC cells that positively regulate CCL2 expression.
E5, E6 or E7 could induce cyclooxygenase-2 (COX-2) expression which leads to increased PGE2 secretion. Among them, HPV-16 E6 and E7 oncoproteins induce COX-2 transcription by inducing the release of EGFR ligand amphiregulin and then activating the epidermal growth factor receptor (EGFR)-Ras-mitogen protein kinase pathway [67,68]. As earlier asserted, PGE2 has a predominant impact on the phenotype of the M2-like macrophage [28].
3.2. T Cell-Derived Molecules
T cells, such as CD4+ and T-regs, infiltrate the tissue in cervical cancer [71]. By secreting specific molecules, it could be that T cells affect the differentiation of monocytes to M2-like macrophages (Table 3).
IL-17 is a cytokine produced by an activated human memory CD4 T-cell Two cervical cell lines transfected with a cDNA encoding IL-17 exhibited a significant increase in tumor size. IL-17 increased IL-6 and IL-8 secretion in cervical carcinoma cell lines. This enhanced tumor growth elicited by IL-17 was associated with increased expression of IL-6 and also macrophage recruitment at the tumor site [72]. Meanwhile, IL-6 has a profound impact on the phenotype of M2-like macrophages [28]. The results implicate that IL-17 may be involved in the differentiation of infiltrating macrophages.
In CC, tumor infiltrating T-effector-cells constitutively expressed IL-4 [79]. IL-4 has been verified to promote bone marrow-derived macrophages (BMDMs) to polarize into the M2a-like subset by inducing a transcriptional factor Kruppel-like factor 4 (KLF4) which then acts in macrophage polarization [73]. Therefore, T-cell derived IL-4 may exert the same role in CC. In addition, IL-10 could induce monocytes to the M2c-like subtype [44]. IL-10 is produced by multiple cells, including T-regs [80]. Low IL-10 levels are associated with increased risk for cervical cancer [43]. So, T-cell derived IL-10 may activate the alternative activation of macrophages in CC. Recently a study has proven that the combination of IL-4 and IL-10 simulate macrophages to the M2a macrophage subtype [74].
3.3. Seminal Plasma-Derived Molecules
Seminal plasma (SP) could activate cyclooxygenase-2 and prostaglandin E2 receptor expression and promote the expression of tumorigenic and angiogenic genes via the E-series prostanoid 4 receptor in cervical adenocarcinoma cells [75,76]. In vitro stimulation of cervical cells with normal seminal plasma resulted in significantly elevated concentrations of secreted IL-6, IL-8 and GM-CSF [77]. Seminal plasma interfered with the attachment, spreading and phagocytic activity of C. parvum elicited murine peritoneal macrophages and the release of reactive oxygen species from zymosan triggered human peripheral blood monocytes [15]. Moreover, prostaglandin E2 and IL-6 acted in M2-like macrophage polarization [28]. These results indicate that SP may affect the differentiation of monocytes via prostaglandin in CC (Table 3).
3.4. Anaerobic Microenvironment
The hypoxic cervical TME stimulates the recruited macrophages to transform into the M2-like phenotype. Overexpressed Nrp-1 in hypoxia-primed cervical cancer cells was necessary for hypoxic cervical TME to recruit and polarize macrophages towards the M2-like phenotype. Nrp-1 and M2-like TAMs have been shown to be related to the malignant properties of cervical cancer, such as the FIGO stage and lymph node metastasis. The results indicate that hypoxic TME play a critical role in activation and pro-tumoral growth by Nrp-1 in cervical cancer [78] (Table 3).
4. Biomarkers of TAMs
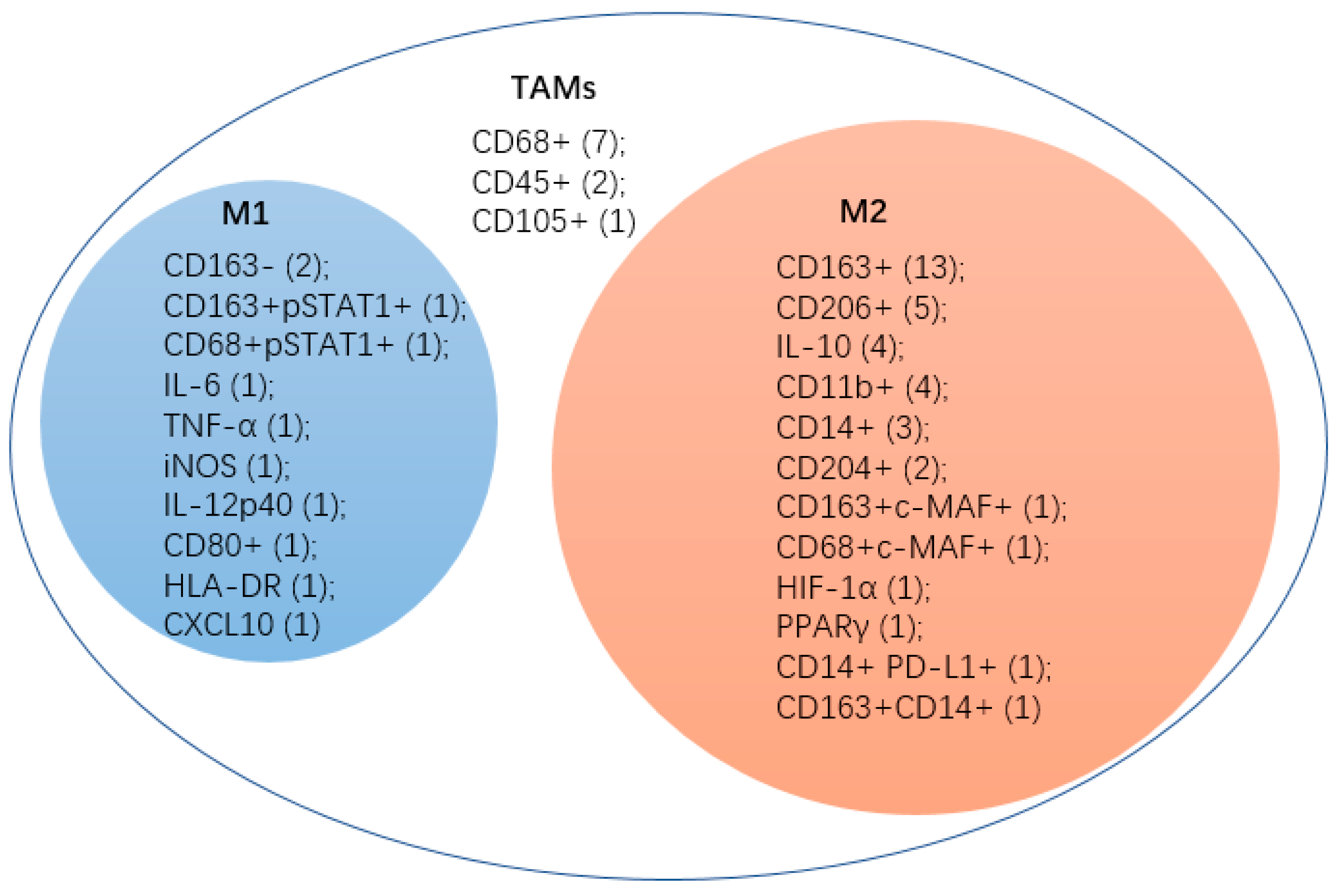
By comparing papers published within the last ten years in the field of macrophage and cervical cancer (Figure 2), we could find that CD68+ was used as a marker to isolate TAMs at an early stage [81,82,83,84,85,86]. Scientists found that CD68+ TAMs in cervical tumor presented a mixed CXCL10 (M1)/CD163+ (M2) pattern [35]. CD45+ and CD105+ are also unspecific for certain subtypes [81,87,88]. Increasing studies have focused on which subtype of macrophages the TAMs belongs to. These studies showed TAMs in CC have a M2-like phenotype because most infiltrating and pro-tumor macrophages were M2-like. M1-like macrophages existed in the CC tumor stroma with a declined expression of M1-like markers such as IL-6, TNF-α and iNOS and tendedto transfer to M2-like phenotype under the influence of tumor cells [27]. Other markers of M1-like were also used in CC research such as CD163- [25,89], CD163+ pSTAT1 [90], IL-12p40 [27], CD80+ [26], HLA-DR [26] and CXCL10 [35]. The most common marker for M2 is CD163+ [25,26,35,73,78,89,91,92,93,94,95,96,97]. CD206+ [26,78,98,99,100] and IL-10 [88,98,100,101]. These are also often used. CD11b+ was considered not very specific for M2-like macrophages. An article pointed out that CD11b+ tumor infiltrating cells may correspond to numerous populations from myeloid-derived suppressor cells to TAMs, or even granulocytes [102]. However, further research has verified that most CD11b+ cells were TAMs with an M2-like phenotype [69] and were adopted by three later studies about CC as an M2-like marker [83,103,104]. CD14+ has been not only used as an M2-like marker in these three articles [84,96,105] but also as the marker of monocytes or macrophages in another article [98]. Many other infrequent markers were also used to recognize M2-like macrophages, such as CD204+ [106], CD163+ c-MAF+ [90], CD68+c-MAF+ [90], HIF-1α [98], PPARγ [98], CD14+PD-L1+ [105] and CD163+CD14+ [28].
5. The Switch from M2 to M1-like Macrophages
Since M2-like TAMs are established to exert tumorigenic effect while M1-like TAMs present anti-tumor effect in CC, some researchers have started to focus on the switch from M2- to M1-like macrophages as a possible breakthrough in treatment (Table 4).
Heusinkveld et al. found that upon interaction with CD4+ Th1 cells, cervical cancer cell-induced M2-like macrophages could be switched to activated M1-like macrophages that express high levels of costimulatory molecules and acquire the lymphoid homing marker CCR7, thus creating a tumor-rejecting milieu [28]. But the concrete component of Th1 on M2-like macrophages was unclear.
Saito T et al. showed that treatment with recombinant human γ-interferon (rIFN-γ) acted direct dose-dependent inhibition of cervical cancer cell line. Moreover, they also found that human adherent ascites cells (greater than 80% macrophages) e showed strong inhibitory properties towards colony growth of both ovarian carcinoma and melanoma cell lines when treated with rIFN-γ [107]. Although lacking direct evidence, these results suggested that TAMs may produce a diffusible substance under the influence of rIFN-γ to become tumoricidal in CC.
6. Development of Therapy by Targeting TAMs
An abundance of molecules has been discovered to act in the formation of TAMs, so the translation of basic research to clinical application is a worthy subject. Here, we will discuss the development of therapy targeting TAMs (Table 5).
6.1. Macrophages and Its Prognostic Value in CC
Findings relating to the role of total macrophages in cervical cancer have been controversial. It has been noted that there are higher macrophage counts in invasive carcinomas as opposed to cervical squamous intraepithelial lesions [117]. Moreover, squamous intraepithelial lesions contain more macrophages than normal cervical tissue [118]. In contrast, a study demonstrated that the presence of this prominent infiltration of macrophages did not correlate with the tumor grade or lymph node status but rather showed a strong negative correlation with the tumor stage [119]. However, Davidson et al. proved that macrophage density did not correlate with patient survival in cervical cancer [120]. What is more, some research has proven that CD68 macrophages which represent all activated macrophages were not a prognostic marker [92,106].
The relationship between M2-like macrophages and CC is specific. The number of intra-tumoral M2 TAMs in cervical SCC tissue samples was significantly higher than that of intraepithelial M2-like TAMs in non-tumorous cervical samples. The number of peritumoral M2 TAMs in cervical SCC was higher than in non-tumorous cervical tissues [91]. A high index of CD163+ macrophages was significantly associated with higher FIGO stages and lymph node metastasis [92]. A higher density of tumor infiltrating CD204+ M2 macrophages in uterine cervical adenocarcinoma was significantly associated with shorter disease-free survival [106].
6.2. Therapy Targeting the Differentiation of TAMs
Indomethacin (Ind.) is a kind of prostaglandin inhibitor. The combination therapy of OK-432 and Ind. reinforced the M phi-mediated immunopotentiation, resulting in a stronger antitumor effect [108]. In addition, treatment with the COX-inhibitor indomethacin and/or the clinical monoclonal antibody against IL-6R, tocilizumab, prevented M2-like-differentiation [109].
Peritoneal macrophages obtained from patients with CC, when treated with sizofiran (SPG) and rIFN-γ, showed a potent tumoricidal quality, as well as, an increase in the section and the inhibition of the secretion of PGE2. [16].
The synthetic long peptide (SLP) vaccination could induce cytokine-producing T-cells to develop a strong macrophage-skewing capacity, which is necessary for CC tissue shrinkage [110].
6.3. Therapy Targeting the Improvement of TAMs Anti-Tumor Activity
SPG-immunotherapy combined with radiotherapy not only induced the cytotoxic activity of macrophages but also augmented NK activity in patients with uterine cervical cancer [111].
Monocytic chemotactic protein-3 (MCP-3) is a CC chemokine originally purified from osteosarcoma cells [121]. In conditions when activated T cells are lacking, heavily infiltrating tumors from hH1/MCP-3-infected HeLa cells with activated macrophages lead to the significant retardation of tumor growth in recipient mice. The result indicates that MCP-3 might activate the phagocytic ability of macrophages [112].
Researchers have found that some HeLa cells produce an uncleavable transmembrane form of TNF (pre-TNF) and that these transformed tumors were compromised in both immunosuppressed and severe combined immunodeficient mice. Macrophages co-cultured with the transformed cells showed increased phagocytosis and cytokine production [113].
Antibodies in serum from a recombinant vaccinia virus expressing the E2 gene of bovine papilloma virus were capable of activating cytotoxicity mediated by infiltrating macrophages for the efficient killing of papilloma tumor cells [114].
Poly methyl methacrylate (PMMA) is a synthetic polymer approved by the Food and Drug Administration for certain human clinical applications such as a bone cement. PMMA 4 particles stimulated the highest level of TNF-α production by macrophages in vitro and yielded the best result of antitumor protection in vivo [115].
A new synthetically prepared fluoroquinolone derivative 6-fluoro-8-nitro-4-oxo-1,4-dihydroquinoline-3-carboxylate (6FN) could induce RAW 264.7 macrophages cell-release of pro- and anti-inflammatory TH1, TH2 and TH17 cytokines with anti-cancer and/or anti-infection activities. A significant inhibition of growth in cancer cells HeLa was detected [116].
7. Conclusions
Circulating monocytes are recruited into local lesions and differentiate into two main subtypes of M1-like and M2-like macrophages, which is modulated by various inflammatory cytokines. In cervical cancer, the functional macrophages in tumor progression are the M2-like subtype formed by molecules from CC cells, T-cells, seminal plasma, and an anaerobic microenvironment. Molecules from CC cells play a major role in their differentiation and their alternative activation. HPV oncoproteins, especially E6 and E7, are the initial factor in affecting the expression of these functional molecules. Molecules from the other sources act mainly in the differentiation of monocytes towards the M2-like phenotype. As M1-like macrophages exert an anti-tumor function in CC, the switch of M2 to M1-like macrophages has captured the attention of scientists. How to re-educate the immunosuppressive characteristics of TAMs is still a challenging and valuable problem to be solved.
Funding
Q.W. was funded by the Chinese Scholarship Council Program (CSC).
Acknowledgments
We thank Heather Mullikin for revision as English native speaker.
Conflicts of Interest
S.M. reports grants and personal fees from AstraZeneca, personal fees from Clovis, grants and personal fees from Medac, grants and personal fees from MSD, personal fees from Novartis, grants and personal fees from PharmaMar, grants and personal fees from Roche, personal fees from Sensor Kinesis, grants and personal fees from Tesaro, grants and personal fees from Teva, outside the submitted work. The other authors declare no conflict of interest.
References
- Naga, C.P.; Gurram, L.; Chopra, S.; Mahantshetty, U. The management of locally advanced cervical cancer. Curr. Opin. Oncol. 2018, 30, 323–329. [Google Scholar]
- Bosch, F.X.; Lorincz, A.; Munoz, N.; Meijer, C.J.; Shah, K.V. The causal relation between human papillomavirus and cervical cancer. J. Clin. Pathol. 2002, 55, 244–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruttkay-Nedecky, B.; Jimenez Jimenez, A.M.; Nejdl, L.; Chudobova, D.; Gumulec, J.; Masarik, M.; Adam, V.; Kizek, R. Relevance of infection with human papillomavirus: The role of the p53 tumor suppressor protein and E6/E7 zinc finger proteins (Review). Int. J. Oncol. 2013, 43, 1754–1762. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, X. Characteristics and significance of the pre-metastatic niche. Cancer Cell 2016, 30, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Schwertfeger, K.L.; Cowman, M.K.; Telmer, P.G.; Turley, E.A.; McCarthy, J.B. Hyaluronan, inflammation, and breast cancer progression. Front. Immunol. 2015, 6, 236. [Google Scholar] [CrossRef] [PubMed]
- Gunther, J. [Experiences with the macrophage electrophoresis mobility test (MEM test) in the diagnosis of malignant gynecological diseases]. Zent. Bakteriol. Orig. B 1975, 91, 529–539. [Google Scholar]
- Cho, J.W.; Kim, W.W.; Lee, Y.M.; Jeon, M.J.; Kim, W.G.; Song, D.E.; Park, Y.; Chung, K.W.; Hong, S.J.; Sung, T.Y. Impact of tumor-associated macrophages and BRAF(V600E) mutation on clinical outcomes in patients with various thyroid cancers. Head Neck 2019, 41, 686–691. [Google Scholar] [CrossRef]
- Hoeffel, G.; Chen, J.; Lavin, Y.; Low, D.; Almeida, F.F.; See, P.; Beaudin, A.E.; Lum, J.; Low, I.; Forsberg, E.C.; et al. C-Myb(+) erythro-myeloid progenitor-derived fetal monocytes give rise to adult tissue-resident macrophages. Immunity 2015, 42, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Gibbings, S.L.; Goyal, R.; Desch, A.N.; Leach, S.M.; Prabagar, M.; Atif, S.M.; Bratton, D.L.; Janssen, W.; Jakubzick, C.V. Transcriptome analysis highlights the conserved difference between embryonic and postnatal-derived alveolar macrophages. Blood 2015, 126, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Kratofil, R.M.; Kubes, P.; Deniset, J.F. Monocyte conversion during inflammation and injury. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Chanmee, T.; Ontong, P.; Konno, K.; Itano, N. Tumor-associated macrophages as major players in the tumor microenvironment. Cancers 2014, 6, 1670–1690. [Google Scholar] [CrossRef] [PubMed]
- Gautier, E.L.; The Immunological Genome Consortium; Shay, T.; Miller, J.; Greter, M.; Jakubzick, C.; Ivanov, S.; Helft, J.; Chow, A.; Elpek, K.G.; et al. Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat. Immunol. 2012, 13, 1118–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantakari, P.; Jäppinen, N.; Lokka, E.; Mokkala, E.; Gerke, H.; Peuhu, E.; Ivaska, J.; Elima, K.; Auvinen, K.; Salmi, M. Fetal liver endothelium regulates the seeding of tissue-resident macrophages. Nature 2016, 538, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Misharin, A.V.; Morales-Nebreda, L.; Reyfman, P.A.; Cuda, C.M.; Walter, J.M.; McQuattie-Pimentel, A.C.; Chen, C.-I.; Anekalla, K.R.; Joshi, N.; Williams, K.J.; et al. Monocyte-derived alveolar macrophages drive lung fibrosis and persist in the lung over the life span. J. Exp. Med. 2017, 214, 2387–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, K.; Harvey, J.; Bradbury, A.W.; Hargreave, T.B.; Cullen, R.T. The effect of seminal plasma on macrophage function—A possible contributory factor in sexually transmitted disease. AIDS Res. 1983, 1, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.T.; Teshima, H.; Shimizu, Y.; Hasumi, K.; Masubuchi, K.; Suzuki, M. [Sizofiran and recombinant interferon γ stimulate peritoneal macrophages obtained from patients with gynecologic malignancies—Increased secretion of tumor necrosis factor, IL-1 and interferon-γ]. Gan Kagaku Ryoho 1990, 17, 1365–1369. [Google Scholar]
- Rojas, J.; Salazar, J.; Martinez, M.S.; Palmar, J.; Bautista, J.; Chavez-Castillo, M.; Gomez, A.; Bermúdez, V. Macrophage heterogeneity and plasticity: Impact of macrophage biomarkers on atherosclerosis. Scientifica (Cairo) 2015, 2015, 851252. [Google Scholar] [CrossRef]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Hao, N.-B.; Lü, M.-H.; Fan, Y.-H.; Cao, Y.-L.; Zhang, Z.-R.; Yang, S.-M. Macrophages in tumor microenvironments and the progression of tumors. Clin. Dev. Immunol. 2012, 2012, 948098. [Google Scholar] [CrossRef]
- Ma, X.; Wu, D.; Zhou, S.; Wan, F.; Liu, H.; Xu, X.; Xu, X.; Zhao, Y.; Tang, M. The pancreatic cancer secreted REG4 promotes macrophage polarization to M2 through EGFR/AKT/CREB pathway. Oncol. Rep. 2016, 35, 189–196. [Google Scholar] [CrossRef]
- Chen, P.-C.; Cheng, H.-C.; Wang, J.; Wang, S.-W.; Tai, H.-C.; Lin, C.-W.; Tang, C.-H. Prostate cancer-derived CCN3 induces M2 macrophage infiltration and contributes to angiogenesis in prostate cancer microenvironment. Oncotarget 2014, 5, 1595–1608. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xu, Y.; Zhong, J.; Wang, H.; Weng, M.; Cheng, Q.; Wu, Q.; Sun, Z.; Jiang, H.; Zhu, M.; et al. MFHAS1 promotes colorectal cancer progress by regulating polarization of tumor-associated macrophages via STAT6 signaling pathway. Oncotarget 2016, 7, 78726–78735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoni, M.; Mauro, G.; Erreni, M.; Romeo, P.; Minna, E.; Vizioli, M.G.; Belgiovine, C.; Rizzetti, M.G.; Pagliardini, S.; Avigni, R.; et al. Senescent thyrocytes and thyroid tumor cells induce M2-like macrophage polarization of human monocytes via a PGE2-dependent mechanism. J. Exp. Clin. Cancer Res. 2019, 38, 208. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K. [Effect of gynecologic cancer sera on functions of monocyte from healthy volunteers]. Nihon Sanka Fujinka Gakkai Zasshi 1982, 34, 1528–1534. [Google Scholar] [PubMed]
- De Vos van Steenwijk, P.J.; Ramwadhdoebe, T.H.; Goedemans, R.; Doorduijn, E.M.; van Ham, J.J.; Gorter, A.; van Hall, T.; Kuijjer, M.L.; van Poelgeest, M.I.; van der Burg, S.H.; et al. Tumor-infiltrating CD14-positive myeloid cells and CD8-positive T-cells prolong survival in patients with cervical carcinoma. Int. J. Cancer 2013, 133, 2884–2894. [Google Scholar] [CrossRef]
- Pedraza-Brindis, E.J.; Sánchez-Reyes, K.; Hernández-Flores, G.; Bravo-Cuellar, A.; Jave-Suárez, L.F.; Aguilar-Lemarroy, A.; Gómez-Lomelí, P.; López-López, B.A.; Ortiz-Lazareno, P.C. Culture supernatants of cervical cancer cells induce an M2 phenotypic profile in THP-1 macrophages. Cell Immunol. 2016, 310, 42–52. [Google Scholar] [CrossRef]
- Li, L.; Yu, S.; Zang, C. Low necroptosis process predicts poor treatment outcome of human papillomavirus positive cervical cancers by decreasing tumor-associated macrophages M1 polarization. Gynecol. Obstet. Investig. 2018, 83, 259–267. [Google Scholar] [CrossRef]
- Heusinkveld, M.; van Steenwijk, P.J.; Goedemans, R.; Ramwadhdoebe, T.H.; Gorter, A.; Welters, M.J.; van Hall, T.; van der Burg, S.H. M2 macrophages induced by prostaglandin E2 and IL-6 from cervical carcinoma are switched to activated M1 macrophages by CD4+ Th1 cells. J. Immunol. 2011, 187, 1157–1165. [Google Scholar] [CrossRef]
- Stelma, T.; Leaner, V.D. KPNB1-mediated nuclear import is required for motility and inflammatory transcription factor activity in cervical cancer cells. Oncotarget 2017, 8, 32833–32847. [Google Scholar] [CrossRef] [Green Version]
- Sierra-Filardi, E.; Nieto, C.; Domínguez-Soto, Á.; Barroso, R.; Sánchez-Mateos, P.; Puig-Kroger, A.; López-Bravo, M.; Joven, J.; Ardavín, C.; Rodríguez-Fernández, J.L.; et al. CCL2 shapes macrophage polarization by GM-CSF and M-CSF: Identification of CCL2/CCR2-dependent gene expression profile. J. Immunol. 2014, 192, 3858–3867. [Google Scholar] [CrossRef]
- Riley, J.K.; Takeda, K.; Akira, S.; Schreiber, R.D. Interleukin-10 receptor signaling through the JAK-STAT pathway. Requirement for two distinct receptor-derived signals for anti-inflammatory action. J. Biol. Chem. 1999, 274, 16513–16521. [Google Scholar] [CrossRef] [PubMed]
- Ramírez de Arellano, A.; Riera Leal, A.; Lopez-Pulido, E.I.; González-Lucano, L.R.; Macías Barragan, J.; del Toro Arreola, S.; García-Chagollan, M.; Palafox-Sánchez, C.A.; Muñoz-Valle, J.F.; Pereira-Suárez, A.L. A 60 kDa prolactin variant secreted by cervical cancer cells modulates apoptosis and cytokine production. Oncol. Rep. 2018, 39, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, A.; Tripathi, A. Prolactin and growth hormone induce differential cytokine and chemokine profile in murine peritoneal macrophages in vitro: Involvement of p-38 MAP kinase, STAT3 and NF-kappaB. Cytokine 2008, 41, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.J. Specificity of macrophage-mediated cytotoxicity: Role of target and effector cell fucose. Immunol. Lett. 1985, 11, 39–44. [Google Scholar] [CrossRef]
- Rigo, A.; Gottardi, M.; Zamò, A.; Mauri, P.; Bonifacio, M.; Krampera, M.; Damiani, E.; Pizzolo, G.; Vinante, F. Macrophages may promote cancer growth via a GM-CSF/HB-EGF paracrine loop that is enhanced by CXCL12. Mol. Cancer 2010, 9, 273. [Google Scholar] [CrossRef] [PubMed]
- Yaddanapudi, K.; Putty, K.; Rendon, B.E.; Lamont, G.J.; Faughn, J.D.; Satoskar, A.; Lasnik, A.; Eaton, J.W.; Mitchell, R.A. Control of tumor-associated macrophage alternative activation by macrophage migration inhibitory factor. J. Immunol. 2013, 190, 2984–2993. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Bae, J.S. Tumor-associated macrophages and neutrophils in tumor microenvironment. Mediat. Inflamm. 2016, 2016, 6058147. [Google Scholar] [CrossRef]
- Nees, M.; Geoghegan, J.M.; Hyman, T.; Frank, S.; Miller, L.; Woodworth, C.D. Papillomavirus type 16 oncogenes downregulate expression of interferon-responsive genes and upregulate proliferation-associated and NF-kappaB-responsive genes in cervical keratinocytes. J. Virol. 2001, 75, 4283–4296. [Google Scholar] [CrossRef]
- Divya, C.S.; Pillai, M.R. Antitumor action of curcumin in human papillomavirus associated cells involves downregulation of viral oncogenes, prevention of NFkB and AP-1 translocation, and modulation of apoptosis. Mol. Carcinog. 2006, 45, 320–332. [Google Scholar] [CrossRef]
- Zijlmans, H.J.; Fleuren, G.J.; Baelde, H.J.; Eilers, P.H.; Kenter, G.G.; Gorter, A. The absence of CCL2 expression in cervical carcinoma is associated with increased survival and loss of heterozygosity at 17q11.2. J. Pathol. 2006, 208, 507–517. [Google Scholar] [CrossRef]
- Alcocer-González, J.M.; Berumen, J.; Tamez-Guerra, R.; Bermúdez-Morales, V.; Peralta-Zaragoza, O.; Hernández-Pando, R.; Moreno, J.; Gariglio, P.; Madrid-Marina, V. In vivo expression of immunosuppressive cytokines in human papillomavirus-transformed cervical cancer cells. Viral. Immunol. 2006, 19, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, F.; Roberts, S.; Mandal, D.; Ollier, B.; Winters, U.; Kitchener, H.; Brabin, L. The IL-10-1082G polymorphism is associated with clearance of HPV infection. BJOG 2006, 113, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Brower, V. Researchers attempting to define role of cytokines in cancer risk. J. Natl. Cancer Inst. 2005, 97, 1175–1177. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Peghini, B.C.; Abdalla, D.R.; Barcelos, A.C.M.; Teodoro, L.D.G.V.L.; Murta, E.F.C.; Michelin, M.A. Local cytokine profiles of patients with cervical intraepithelial and invasive neoplasia. Hum. Immunol. 2012, 73, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Riteau, B.; Rouas-Freiss, N.; Menier, C.; Paul, P.; Dausset, J.; Carosella, E.D. HLA-G2, -G3, and -G4 isoforms expressed as nonmature cell surface glycoproteins inhibit NK and antigen-specific CTL cytolysis. J. Immunol. 2001, 166, 5018–5026. [Google Scholar] [CrossRef]
- Rodríguez, J.A.; Galeano, L.; Palacios, D.M.; Gómez, C.; Serrano, M.L.; Bravo, M.M.; Combita, A.L. Altered HLA class I and HLA-G expression is associated with IL-10 expression in patients with cervical cancer. Pathobiology 2012, 79, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Vesce, F.; Biondi, C. α-l-Fucosidase activity in endometrial, cervical and ovarian cancer. Eur. J. Gynaecol. Oncol. 1983, 4, 135–138. [Google Scholar] [PubMed]
- Chopra, V.; Dinh, T.V.; Hannigan, E.V. Circulating serum levels of cytokines and angiogenic factors in patients with cervical cancer. Cancer Investig. 1998, 16, 152–159. [Google Scholar] [CrossRef]
- Zijlmans, H.J.; Fleuren, G.J.; Baelde, H.J.; Eilers, P.H.; Kenter, G.G.; Gorter, A. Role of tumor-derived proinflammatory cytokines GM-CSF, TNF-alpha, and IL-12 in the migration and differentiation of antigen-presenting cells in cervical carcinoma. Cancer 2007, 109, 556–565. [Google Scholar] [CrossRef]
- Krockenberger, M.; Engel, J.B.; Kolb, J.; Dombrowsky, Y.; Häusler, S.F.; Kohrenhagen, N.; Dietl, J.; Wischhusen, J.; Honig, A. Macrophage migration inhibitory factor expression in cervical cancer. J. Cancer Res. Clin. Oncol. 2010, 136, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Beswick, E.J.; Reyes, V.E. CD74 in antigen presentation, inflammation, and cancers of the gastrointestinal tract. World J. Gastroenterol. 2009, 15, 2855–2861. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lian, J.; Tao, H.; Shang, H.; Zhang, L. Correlation of macrophage migration inhibitory factor gene polymorphism with the risk of early-stage cervical cancer and lymphatic metastasis. Oncol. Lett. 2011, 2, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.J.; Deng, W.G.; Niu, C.B.; Li, Y.Y.; Fu, Y. Expression of macrophage migration inhibitory factor and CD74 in cervical squamous cell carcinoma. Int. J. Gynecol. Cancer 2011, 21, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.Z.; Dai, B.; Chen, J.; Luo, Q.; Liu, X.Y.; Lin, Q.X.; Li, X.H.; Huang, W.; Yu, X.Y. Loss of macrophage migration inhibitory factor impairs the growth properties of human HeLa cervical cancer cells. Cell Prolif. 2011, 44, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Corraliza, I.M.; Soler, G.; Eichmann, K.; Modolell, M. Arginase induction by suppressors of nitric oxide synthesis (IL-4, IL-10 and PGE2) in murine bone-marrow-derived macrophages. Biochem. Biophys. Res. Commun. 1995, 206, 667–673. [Google Scholar] [CrossRef]
- Taylor, P.; Martinez-Pomares, L.; Stacey, M.; Lin, H.-H.; Brown, G.; Gordon, S.; Taylor, P. Macrophage receptors and immune recognition. Annu. Rev. Immunol. 2005, 23, 901–944. [Google Scholar] [CrossRef]
- Chen, Z.; Terai, M.; Fu, L.; Herrero, R.; DeSalle, R.; Burk, R.D. Diversifying selection in human papillomavirus type 16 lineages based on complete genome analyses. J. Virol. 2005, 79, 7014–7023. [Google Scholar] [CrossRef]
- Bravo, I.G.; Felez-Sanchez, M. Papillomaviruses: Viral evolution, cancer and evolutionary medicine. Evol. Med. Public Health 2015, 2015, 32–51. [Google Scholar] [CrossRef]
- Doorbar, J. The E4 protein; structure, function and patterns of expression. Virology 2013, 445, 80–98. [Google Scholar] [CrossRef] [Green Version]
- Galloway, D.A.; Laimins, L.A. Human papillomaviruses: Shared and distinct pathways for pathogenesis. Curr. Opin. Virol. 2015, 14, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25 (Suppl. 1), 2–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, N.A.; Galloway, D.A. Novel functions of the human papillomavirus E6 oncoproteins. Annu. Rev. Virol. 2015, 2, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Riethdorf, S.; Riethdorf, L.; Richter, N.; Loning, T. Expression of the MCP-1 gene and the HPV 16 E6/E7 oncogenes in squamous cell carcinomas of the cervix uteri and metastases. Pathobiology 1998, 66, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Berti, F.C.B.; Pereira, A.P.L.; Cebinelli, G.C.M.; Trugilo, K.P.; de Oliveira, K.B. The role of interleukin 10 in human papilloma virus infection and progression to cervical carcinoma. Cytokine Growth Factor Rev. 2017, 34, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pahler, J.C.; Tazzyman, S.; Erez, N.; Chen, Y.-Y.; Murdoch, C.; Nozawa, H.; Lewis, C.E.; Hanahan, D. Plasticity in tumor-promoting inflammation: Impairment of macrophage recruitment evokes a compensatory neutrophil response. Neoplasia 2008, 10, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.M.; Kim, S.H.; Lee, Y.I.; Seo, M.; Kim, S.Y.; Song, Y.S.; Kim, W.H.; Juhnn, Y.S. Human papillomavirus E5 protein induces expression of the EP4 subtype of prostaglandin E2 receptor in cyclic AMP response element-dependent pathways in cervical cancer cells. Carcinogenesis 2009, 30, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Subbaramaiah, K.; Dannenberg, A.J. Cyclooxygenase-2 transcription is regulated by human papillomavirus 16 E6 and E7 oncoproteins: Evidence of a corepressor/coactivator exchange. Cancer Res. 2007, 67, 3976–3985. [Google Scholar] [CrossRef] [PubMed]
- Bolpetti, A.; Silva, J.S.; Villa, L.L.; Lepique, A.P. Interleukin-10 production by tumor infiltrating macrophages plays a role in Human Papillomavirus 16 tumor growth. BMC Immunol. 2010, 11, 27. [Google Scholar] [CrossRef]
- Liang, F.; Kina, S.; Takemoto, H.; Matayoshi, A.; Phonaphonh, T.; Sunagawa, N.; Arakaki, K.; Arasaki, A.; Kuang, H.; Sunakawa, H. HPV16E6-dependent c-fos expression contributes to AP-1 complex formation in SiHa cells. Mediat. Inflamm. 2011, 2011, 263216. [Google Scholar] [CrossRef]
- Balkwill, F.; Mantovani, A.; Balkwill, F. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Tartour, E.; Fossiez, F.; Joyeux, I.; Galinha, A.; Gey, A.; Claret, E.; Sastre-Garau, X.; Couturier, J.; Mosseri, V.; Vives, V.; et al. Interleukin 17, a T-cell-derived cytokine, promotes tumorigenicity of human cervical tumors in nude mice. Cancer Res. 1999, 59, 3698–3704. [Google Scholar] [PubMed]
- Yang, W.; Lu, Y.-P.; Yang, Y.-Z.; Kang, J.-R.; Jin, Y.-D.; Wang, H.-W.; Lu, Y.; Kang, J.; Jin, Y.; Wang, H. Expressions of programmed death (PD)-1 and PD-1 ligand (PD-L1) in cervical intraepithelial neoplasia and cervical squamous cell carcinomas are of prognostic value and associated with human papillomavirus status. J. Obstet. Gynaecol. Res. 2017, 43, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Makita, N.; Hizukuri, Y.; Yamashiro, K.; Murakawa, M.; Hayashi, Y. IL-10 enhances the phenotype of M2 macrophages induced by IL-4 and confers the ability to increase eosinophil migration. Int. Immunol. 2015, 27, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Sales, K.J.; Katz, A.A.; Millar, R.P.; Jabbour, H.N. Seminal plasma activates cyclooxygenase-2 and prostaglandin E2 receptor expression and signalling in cervical adenocarcinoma cells. Mol. Hum. Reprod. 2002, 8, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.; Sales, K.J.; Katz, A.A.; Jabbour, H.N. Seminal plasma promotes the expression of tumorigenic and angiogenic genes in cervical adenocarcinoma cells via the E-series prostanoid 4 receptor. Endocrinology 2006, 147, 3356–3365. [Google Scholar] [CrossRef]
- Rametse, C.L.; Adefuye, A.O.; Olivier, A.J.; Curry, L.; Gamieldien, H.; Burgers, W.A.; Lewis, D.A.; Williamson, A.-L.; Katz, A.A.; Passmore, J.-A.S. Inflammatory cytokine profiles of semen influence cytokine responses of cervicovaginal epithelial cells. Front. Immunol. 2018, 9, 2721. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Wu, S.; Yan, R.M.; Fan, L.S.; Yu, L.; Zhang, Y.M.; Wei, W.F.; Zhou, C.F.; Wu, X.G.; Zhong, M.; et al. The role of the hypoxia-Nrp-1 axis in the activation of M2-like tumor-associated macrophages in the tumor microenvironment of cervical cancer. Mol. Carcinog. 2019, 58, 388–397. [Google Scholar] [CrossRef]
- Adurthi, S.; Mukherjee, G.; Krishnamurthy, H.; Sudhir, K.; Bafna, U.D.; Umadevi, K.; Jayshree, R.S. Functional tumor infiltrating TH1 and TH2 effectors in large early-stage cervical cancer are suppressed by regulatory T cells. Int. J. Gynecol. Cancer 2012, 22, 1130–1137. [Google Scholar] [CrossRef]
- Akiyama, S.K.; Yamada, S.S.; Yamada, K.M. Analysis of the role of glycosylation of the human fibronectin receptor. J. Biol. Chem. 1989, 264, 18011–18018. [Google Scholar]
- Jiang, S.; Yang, Y.; Fang, M.; Li, X.; Yuan, X.; Yuan, J. Co-evolution of tumor-associated macrophages and tumor neo-vessels during cervical cancer invasion. Oncol. Lett. 2016, 12, 2625–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoppmann, S.F.; Birner, P.; Stöckl, J.; Kalt, R.; Ullrich, R.; Caucig, C.; Kriehuber, E.; Nagy, K.; Alitalo, K.; Kerjaschki, D. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Am. J. Pathol. 2002, 161, 947–956. [Google Scholar] [CrossRef]
- Prata, T.T.M.; Bonin, C.M.; Ferreira, A.M.T.; Padovani, C.T.J.; Fernandes, C.E.D.S.; Machado, A.P.; Tozetti, I.A. Local immunosuppression induced by high viral load of human papillomavirus: Characterization of cellular phenotypes producing interleukin-10 in cervical neoplastic lesions. Immunology 2015, 146, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Cai, J.; Mao, M.; Fang, Y.; Huang, Z.; Jia, J.; Li, T.; Xu, L.; Wang, J.; Zhou, J.; et al. Tumor-associated macrophages induce lymphangiogenesis in cervical cancer via interaction with tumor cells. APMIS 2014, 122, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kang, S.; Chen, X.; Kim, E.J.; Kim, J.; Kim, N.; Kim, J.; Jin, M.M. Tumor suppression via paclitaxel-loaded drug carriers that target inflammation marker upregulated in tumor vasculature and macrophages. Biomaterials 2013, 34, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Ozhegov, E.; Berg, Y.W.V.D.; Aronow, B.J.; Franco, R.S.; Palascak, M.B.; Fallon, J.T.; Ruf, W.; Versteeg, H.; Bogdanov, V.; et al. Splice variants of tissue factor promote monocyte-endothelial interactions by triggering the expression of cell adhesion molecules via integrin-mediated signaling. J. Thromb. Haemost. 2011, 9, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Silveira, C.R.; Cipelli, M.; Manzine, C.; Rabelo-Santos, S.H.; Zeferino, L.C.; Rodriguez, G.R.; de Assis, J.B.; Hebster, S.; Bernadinelli, I.; Laginha, F.; et al. Swainsonine, an alpha-mannosidase inhibitor, may worsen cervical cancer progression through the increase in myeloid derived suppressor cells population. PLoS ONE 2019, 14, e0213184. [Google Scholar] [CrossRef]
- Lepique, A.P.; Daghastanli, K.R.; Cuccovia, I.M.; Villa, L.L. HPV16 tumor-associated macrophages suppress antitumor T cell responses. Clin. Cancer Res. 2009, 15, 4391–4400. [Google Scholar] [CrossRef]
- Sánchez-Reyes, K.; Bravo-Cuellar, A.; Hernández-Flores, G.; Lerma-Díaz, J.M.; Jave-Suarez, L.F.; Gómez-Lomeli, P.; de Celis, R.; Aguilar-Lemarroy, A.; Domínguez-Rodríguez, J.R.; Ortiz-Lazareno, P.C. Cervical cancer cell supernatants induce a phenotypic switch from U937-derived macrophage-activated M1 state into M2-like suppressor phenotype with change in Toll-like receptor profile. Biomed. Res. Int. 2014, 2014, 683068. [Google Scholar] [CrossRef]
- Petrillo, M.; Zannoni, G.F.; Martinelli, E.; Anchora, L.P.; Ferrandina, G.; Tropeano, G.; Fagotti, A.; Scambia, G. Polarisation of tumor-associated macrophages toward M2 phenotype correlates with poor response to chemoradiation and reduced survival in patients with locally advanced cervical cancer. PLoS ONE 2015, 10, e0136654. [Google Scholar] [CrossRef]
- Li, Y.; Huang, G.; Zhang, S. Associations between intratumoral and peritumoral M2 macrophage counts and cervical squamous cell carcinoma invasion patterns. Int. J. Gynaecol. Obstet. 2017, 139, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Han, L.F.; Wu, X.G.; Wei, W.F.; Wu, L.F.; Yi, H.Y.; Yan, R.M.; Bai, X.Y.; Zhong, M.; Yu, Y.H.; et al. Clinical significance of CD163+ and CD68+ tumor-associated macrophages in high-risk HPV-related cervical cancer. J. Cancer 2017, 8, 3868–3875. [Google Scholar] [CrossRef] [PubMed]
- Rotman, J.; Mom, C.H.; Jordanova, E.S.; de Gruijl, T.D.; Kenter, G.G. ‘DURVIT’: A phase-I trial of single low-dose durvalumab (Medi4736) IntraTumourally injected in cervical cancer: Safety, toxicity and effect on the primary tumour- and lymph node microenvironment. BMC Cancer 2018, 18, 888. [Google Scholar] [CrossRef]
- Swangphon, P.; Pientong, C.; Sunthamala, N.; Bumrungthai, S.; Azuma, M.; Kleebkaow, P.; Tangsiriwatthana, T.; Sangkomkamhang, U.; Kongyingyoes, B.; Ekalaksananan, T. Correlation of circulating CD64(+)/CD163(+) monocyte ratio and stroma/peri-tumoral CD163(+) monocyte density with human papillomavirus infected cervical lesion severity. Cancer Microenviron. 2017, 10, 77–85. [Google Scholar] [CrossRef]
- Li, L.; Ma, Y.; Liu, S.; Zhang, J.; Xu, X.Y. Interleukin 10 promotes immune response by increasing the survival of activated CD8(+) T cells in human papillomavirus 16-infected cervical cancer. Tumour Biol. 2016, 37, 16093–16101. [Google Scholar] [CrossRef] [PubMed]
- Heeren, A.M.; Punt, S.; Bleeker, M.C.; Gaarenstroom, K.N.; Van Der Velden, J.; Kenter, G.G.; De Gruijl, T.D.; Jordanova, E.S. Prognostic effect of different PD-L1 expression patterns in squamous cell carcinoma and adenocarcinoma of the cervix. Mod. Pathol. 2016, 29, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Punt, S.; Thijssen, V.L.; Vrolijk, J.; De Kroon, C.D.; Gorter, A.; Jordanova, E.S. Galectin-1, -3 and -9 expression and clinical significance in squamous cervical cancer. PLoS ONE 2015, 10, e0129119. [Google Scholar] [CrossRef]
- Stone, S.C.; Rossetti, R.A.M.; Alvarez, K.L.F.; Carvalho, J.P.; Margarido, P.F.R.; Baracat, E.C.; Tacla, M.; Boccardo, E.; Yokochi, K.; Lorenzi, N.P.; et al. Lactate secreted by cervical cancer cells modulates macrophage phenotype. J. Leukoc. Biol. 2019, 105, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, K.L.; Beldi, M.; Sarmanho, F.; Rossetti, R.A.; Silveira, C.R.; Mota, G.R.; Andreoli, M.A.; de Carvalho Caruso, E.D.; Kamillos, M.F.; Souza, A.M.; et al. Local and systemic immunomodulatory mechanisms triggered by Human Papillomavirus transformed cells: A potential role for G-CSF and neutrophils. Sci. Rep. 2017, 7, 9002. [Google Scholar] [CrossRef]
- Strachan, D.C.; Ruffell, B.; Oei, Y.; Bissell, M.J.; Coussens, L.M.; Pryer, N.; Daniel, D. CSF1R inhibition delays cervical and mammary tumor growth in murine models by attenuating the turnover of tumor-associated macrophages and enhancing infiltration by CD8(+) T cells. Oncoimmunology 2013, 2, e26968. [Google Scholar] [CrossRef]
- Li, Q.; Liu, L.; Zhang, Q.; Liu, S.; Ge, D.; You, Z. Interleukin-17 indirectly promotes M2 macrophage differentiation through stimulation of COX-2/PGE2 pathway in the cancer cells. Cancer Res. Treat. 2014, 46, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Bronte, V. Altered macrophage differentiation and immune dysfunction in tumor development. J. Clin. Investig. 2007, 117, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.V.; Ajay, A.K.; Mohammad, N.; Malvi, P.; Chaube, B.; Meena, A.S.; Bhat, M.K. Proteasomal inhibition sensitizes cervical cancer cells to mitomycin C-induced bystander effect: The role of tumor microenvironment. Cell Death Dis. 2015, 6, e1934. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Muramatsu, M.; Wang, F.; Tsuchida, R.; Kodama, T.; Minami, T.; Shibuya, M. Increased expression of histone demethylase JHDM1D under nutrient starvation suppresses tumor growth via down-regulating angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 20725–20729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heeren, A.M.; Kenter, G.G.; Jordanova, E.S.; de Gruijl, T.D. CD14(+) macrophage-like cells as the linchpin of cervical cancer perpetrated immune suppression and early metastatic spread: A new therapeutic lead? Oncoimmunology 2015, 4, e1009296. [Google Scholar] [CrossRef] [PubMed]
- Kawachi, A.; Yoshida, H.; Kitano, S.; Ino, Y.; Kato, T.; Hiraoka, N. Tumor-associated CD204(+) M2 macrophages are unfavorable prognostic indicators in uterine cervical adenocarcinoma. Cancer Sci. 2018, 109, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Berens, M.E.; Welander, C.E. Direct and indirect effects of human recombinant γ-interferon on tumor cells in a clonogenic assay. Cancer Res. 1986, 46, 1142–1147. [Google Scholar] [PubMed]
- Ohshika, Y.; Umesaki, N.; Sugawa, T. Immunomodulating capacity of the monocyte-macrophage system in patients with uterine cervical cancer. Nihon Sanka Fujinka Gakkai Zasshi 1988, 40, 601–608. [Google Scholar] [PubMed]
- Dijkgraaf, E.M.; Heusinkveld, M.; Tummers, B.; Vogelpoel, L.T.C.; Goedemans, R.; Jha, V.; Nortier, J.W.R.; Welters, M.J.P.; Kroep, J.R.; Van Der Burg, S.H. Chemotherapy alters monocyte differentiation to favor generation of cancer-supporting M2 macrophages in the tumor microenvironment. Cancer Res. 2013, 73, 2480–2492. [Google Scholar] [CrossRef] [PubMed]
- Van der Sluis, T.C.; Sluijter, M.; van Duikeren, S.; West, B.L.; Melief, C.J.; Arens, R.; van der Burg, S.H.; van Hall, T. Therapeutic peptide vaccine-induced CD8 T cells strongly modulate intratumoral macrophages required for tumor regression. Cancer Immunol. Res. 2015, 3, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Nishimura, R.; Kinugasa, M.; Okamura, M.; Kimura, A.; Ohtsu, F.; Takeuchi, K.; Mizuhira, V. [Electron microscopic and immunological studies concerning the effect on the antitumor activity of sizofiran (SPG) combined with radiotherapy for cervical cancer]. Nihon Gan Chiryo Gakkai Shi 1990, 25, 2549–2561. [Google Scholar] [PubMed]
- Wetzel, K.; Menten, P.; Van Damme, J.; Giese, N.; Vecchi, A.; Sozzani, S.; Cornelis, J.J.; Rommelaere, J.; Dinsart, C. Transduction of human MCP-3 by a parvoviral vector induces leukocyte infiltration and reduces growth of human cervical carcinoma cell xenografts. J. Gene Med. 2001, 3, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Nagy, T.; Glavinas, H.; Szincsák, N.; Hunyadi, J.; Jánossy, T.; Duda, E.; Vizler, C.; Juhász, I. Tumor cells expressing membrane-bound tumor necrosis factor activate macrophages and have a compromised growth in immunosuppressed and immunodeficient mice. Cancer Lett. 2003, 196, 49–56. [Google Scholar] [CrossRef]
- Rosales, C.; Graham, V.V.; Rosas, G.A.; Merchant, H.; Rosales, R. A recombinant vaccinia virus containing the papilloma E2 protein promotes tumor regression by stimulating macrophage antibody-dependent cytotoxicity. Cancer Immunol. Immunother. 2000, 49, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Lou, P.-J.; Cheng, W.-F.; Chung, Y.-C.; Cheng, C.-Y.; Chiu, L.-H.; Young, T.-H. PMMA particle-mediated DNA vaccine for cervical cancer. J. Biomed. Mater. Res. A 2009, 88, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Jantová, S.; Paulovičová, E.; Paulovičová, L.; Janošková, M.; Pánik, M.; Milata, V. Immunobiological efficacy and immunotoxicity of novel synthetically prepared fluoroquinolone ethyl 6-fluoro-8-nitro-4-oxo-1,4-dihydroquinoline-3-carboxylate. Immunobiology 2018, 223, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Goldberg, I.; Kopolovic, J. Inflammatory response in cervical intraepithelial neoplasia and squamous cell carcinoma of the uterine cervix. Pathol. Res. Pract. 1997, 193, 491–495. [Google Scholar] [CrossRef]
- Al-Saleh, W.; Delvenne, P.; Arrese, J.E.; Boniver, J.; Nikkels, A.F. Inverse modulation of intraepithelial Langerhans’ cells and stromal macrophage/dendrocyte populations in human papillomavirus-associated squamous intraepithelial lesions of the cervix. Virchows Arch. 1995, 427, 41–48. [Google Scholar] [CrossRef]
- Heller, D.S.; Hameed, M.; Cracchiolo, B.; Wiederkehr, M.; Scott, D.; Skurnick, J.; Ammar, N.; Lambert, W.C. Presence and quantification of macrophages in squamous cell carcinoma of the cervix. Int. J. Gynecol. Cancer 2003, 13, 67–70. [Google Scholar] [CrossRef]
- Davidson, B.; Goldberg, I.; Gotlieb, W.H.; Lerner-Geva, L.; Ben-Baruch, G.; Agulansky, L.; Novikov, I.; Kopolovic, J. Macrophage infiltration and angiogenesis in cervical squamous cell carcinoma—Clinicopathologic correlation. Acta Obstet. Gynecol. Scand. 1999, 78, 240–244. [Google Scholar] [CrossRef]
- Van Damme, J.; Proost, P.; Lenaerts, J.P.; Opdenakker, G. Structural and functional identification of two human, tumor-derived monocyte chemotactic proteins (MCP-2 and MCP-3) belonging to the chemokine family. J. Exp. Med. 1992, 176, 59–65. [Google Scholar] [CrossRef] [PubMed]
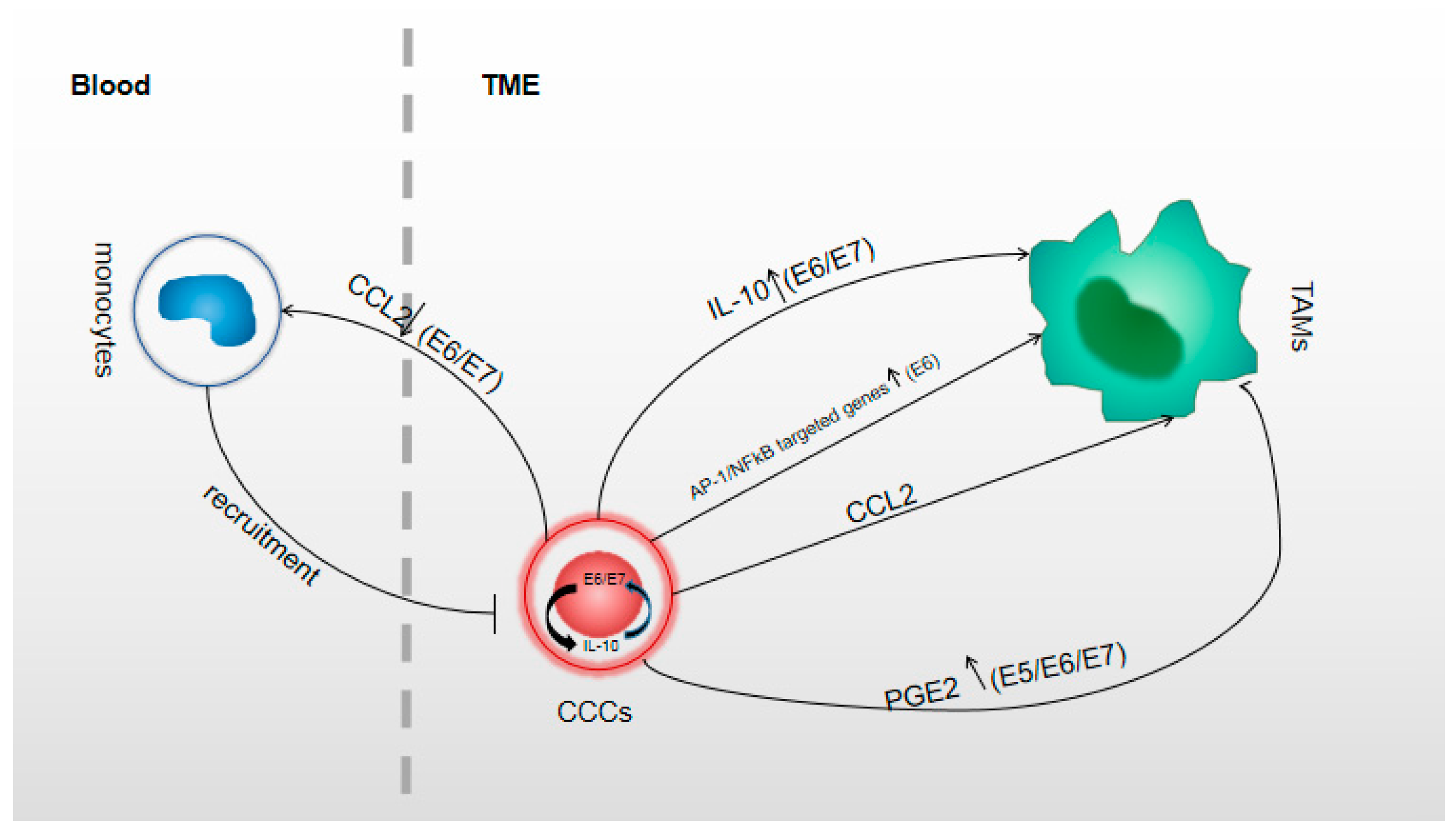
Figure 1.
This figure shows the relationship between human papillomavirus (HPV) oncoproteins and the molecules from CC cells acting in formatting TAMs. In circulating blood, CCL2 (left) was regulated inversely by E6 or E7 [64], and the less expressed CCL2 recruited fewer monocytes into local tumor tissue [30]; IL-10 was upregulated by E6 or E7 and E6 and E7 were upregulated by IL-10 as a circulation in CC cells [65]; Increasing IL-10 could act in M2 polarization process [44]; Transcriptional factors AP-1 and NFκB targeted genes that act in M2 polarization were upregulated by E6 [38]; CCL2 (right) was expressed individualized [30,40,44,66] and CCL2+ CC cells could promote the M2 polarization [30,40]. The molecules which regulated CCL2 expression that act in M2 polarization was unclear. PGE2 which acted in M2 polarization was upregulated by E5 or E6 or E7 [67,68]; TME, tumor microenvironment; CCL2 is the alternative name of monocyte chemoattractant protein (MCP-1); IL-10, interleukin 10; CC cells, cervical cancer cells.
Figure 1.
This figure shows the relationship between human papillomavirus (HPV) oncoproteins and the molecules from CC cells acting in formatting TAMs. In circulating blood, CCL2 (left) was regulated inversely by E6 or E7 [64], and the less expressed CCL2 recruited fewer monocytes into local tumor tissue [30]; IL-10 was upregulated by E6 or E7 and E6 and E7 were upregulated by IL-10 as a circulation in CC cells [65]; Increasing IL-10 could act in M2 polarization process [44]; Transcriptional factors AP-1 and NFκB targeted genes that act in M2 polarization were upregulated by E6 [38]; CCL2 (right) was expressed individualized [30,40,44,66] and CCL2+ CC cells could promote the M2 polarization [30,40]. The molecules which regulated CCL2 expression that act in M2 polarization was unclear. PGE2 which acted in M2 polarization was upregulated by E5 or E6 or E7 [67,68]; TME, tumor microenvironment; CCL2 is the alternative name of monocyte chemoattractant protein (MCP-1); IL-10, interleukin 10; CC cells, cervical cancer cells.

Figure 2.
This figure shows the makers which were used in cervical cancer researches. M1, M1-like macrophages; M2, M2-like macrophages; TAMs, tumor-associated macrophages. The numbers in the parentheses represent the number of published papers.
Figure 2.
This figure shows the makers which were used in cervical cancer researches. M1, M1-like macrophages; M2, M2-like macrophages; TAMs, tumor-associated macrophages. The numbers in the parentheses represent the number of published papers.


{kind=link}
{kind=link}
Table 1.
The categories and characteristics of macrophages.
| Categories | Characteristics |
|---|---|
| Tissue-resident macrophages | |
| Infiltrating macrophages |
|
The table above shows the categories of macrophages and their characteristics respectively.
Table 2.
Tumor-derived molecules acting in tumor-associated macrophage (TAM) formation in cervical cancer (CC).
Table 2.
Tumor-derived molecules acting in tumor-associated macrophage (TAM) formation in cervical cancer (CC).
| Categories | Name of Molecules | Potential Mechanism |
|---|---|---|
| Molecules promoting the differentiation of monocytes to TAMs | PGE2 and IL-6 [28] | |
| Karyopherin β1 | Regulating the expression of transcriptional factors NFκB and AP-1 [29] | |
| CCL2 /MCP-1 | ||
| IL-10 | Inhibit the classic activation of macrophages through the JAK1/Tyk2/STAT3 pathway [31] | |
| PRL | ||
| Molecules promoting the activation of TAMs | FUCA-1 | |
| GM-CSF | Promoting TAMs to release pro-tumor factors [35] | |
| MIF [36] | ||
| IL-10 |
The table shows tumor-derived molecules and their potential mechanism to form TAMs. PGE2, prostaglandin E2; IL-6, interleukin 6; MCP-1, monocyte chemoattractant protein 1, its alternative name is CCL2; IL-10, interleukin 10; PRL, prolactin; FUCA-1, α-l-Fucosidase; GM-CSF, granulocyte macrophage colony-stimulating factor; MIF, Macrophage migration inhibitory factor; LPS, lipopolysaccharide; TNF-α, tumor necrosis factor α; IL-4R, interleukin 4 receptor.
Table 3.
Other molecules acting in the formation of TAMs in CC.
| Source | Name of Molecules | Potential Mechanism |
|---|---|---|
| T cell | IL-17 | |
| IL-4 | Promoting M2a subtype polarization by inducing a transcriptional factor KLF4 [73] | |
| IL-10 | Promoting M2c subtype polarization [44] | |
| IL-4 and IL-10 | Promoting M2a subtype polarization [74] | |
| Seminal plasma | Increasing the expression of COX2, PGE2, IL-6 in CC cells [75,76,77] | |
| Anaerobic microenvironment | Promoting M2 polarization by overexpressing Nrp-1 in CC cells [78] |
The table shows the role of the molecules from T cell, seminal plasma and anaerobic microenvironment to the formation of TAMs. IL-17, interleukin 17; IL-6, interleukin 6; KLF4, Kruppel-like factor 4; IL-10, interleukin 10; COX2, cyclooxygenase-2; PGE2, prostaglandin E2; Nrp-1, neuropilin-1.
Table 4.
Component Involved in the Switch from M2 to M1-like macrophages.
| Component | Potential Mechanism | |
|---|---|---|
| Targeting the differentiation of TAMs | CD4+ Th1 cells | Promoting TAMs to secret costimulatory molecules and the expression of CCR7 [28] |
| rIFN-γ | Promoting TAMs to produce a diffusible tumoricidal substance [107] |
The table above shows the component involved in the switch from M2 to M1-like macrophages. TAMs, tumor-associated macrophages; rIFN-γ, recombinant human γ-interferon; Th1, 1-type helper T cells.
Table 5.
Molecules for Therapy by Targeting TAMs.
| Molecules | Potential Mechanism | |
|---|---|---|
| Targeting the differentiation of TAMs | Ind. | Inhibiting the expression of PG [108] |
| tocilizumab | Blocking IL-6R [109] | |
| rIFN-γ | Simulating the role of IFN-γ to induce M2 to M1-like macrophages [16] | |
| SLP | Inducing T cells to inflow [110] | |
| Improvement in anti-tumor activity of TAMs | SPG | Inducing the cytotoxic activity of macrophage [111] |
| MCP-3 | Activating the phagocytic ability of macrophages [112] | |
| Pre-TNF | Increasing phagocytosis [113] | |
| bovine papilloma virus antibody | Inducing the cytotoxic activity of macrophage [114] | |
| PMMA | Stimulating TAMs to produce TNF-α [115] | |
| 6FN | Inducing TAMs to release anti-tumor cytokines [116] |
The table above shows the molecules used in therapy by targeting TAMs. Ind., Indomethacin; PG, prostaglandin; rIFN-γ, recombined interferon-γ; IL-6R, interleukin-6 receptor; SLP, synthetic long peptide; SPG, sizofiran; MCP-3, monocyte chemoattractant protein; pre-TNF, an uncleavable transmembrane form of tumor necrosis factor; PMMA, Poly methyl methacrylate; 6FN, 6-fluoro-8-nitro-4-oxo-1,4-dihydroquinoline-3-carboxylate.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Q.; Steger, A.; Mahner, S.; Jeschke, U.; Heidegger, H. The Formation and Therapeutic Update of Tumor-Associated Macrophages in Cervical Cancer. Int. J. Mol. Sci. 2019, 20, 3310. https://doi.org/10.3390/ijms20133310
AMA Style
Wang Q, Steger A, Mahner S, Jeschke U, Heidegger H. The Formation and Therapeutic Update of Tumor-Associated Macrophages in Cervical Cancer. International Journal of Molecular Sciences. 2019; 20(13):3310. https://doi.org/10.3390/ijms20133310
Chicago/Turabian StyleWang, Qun, Alexander Steger, Sven Mahner, Udo Jeschke, and Helene Heidegger. 2019. "The Formation and Therapeutic Update of Tumor-Associated Macrophages in Cervical Cancer" International Journal of Molecular Sciences 20, no. 13: 3310. https://doi.org/10.3390/ijms20133310
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.