Human Fetal Bone Marrow-Derived Mesenchymal Stem Cells Promote the Proliferation and Differentiation of Pancreatic Progenitor Cells and the Engraftment Function of Islet-Like Cell Clusters


Abstract
:1. Introduction
2. Results
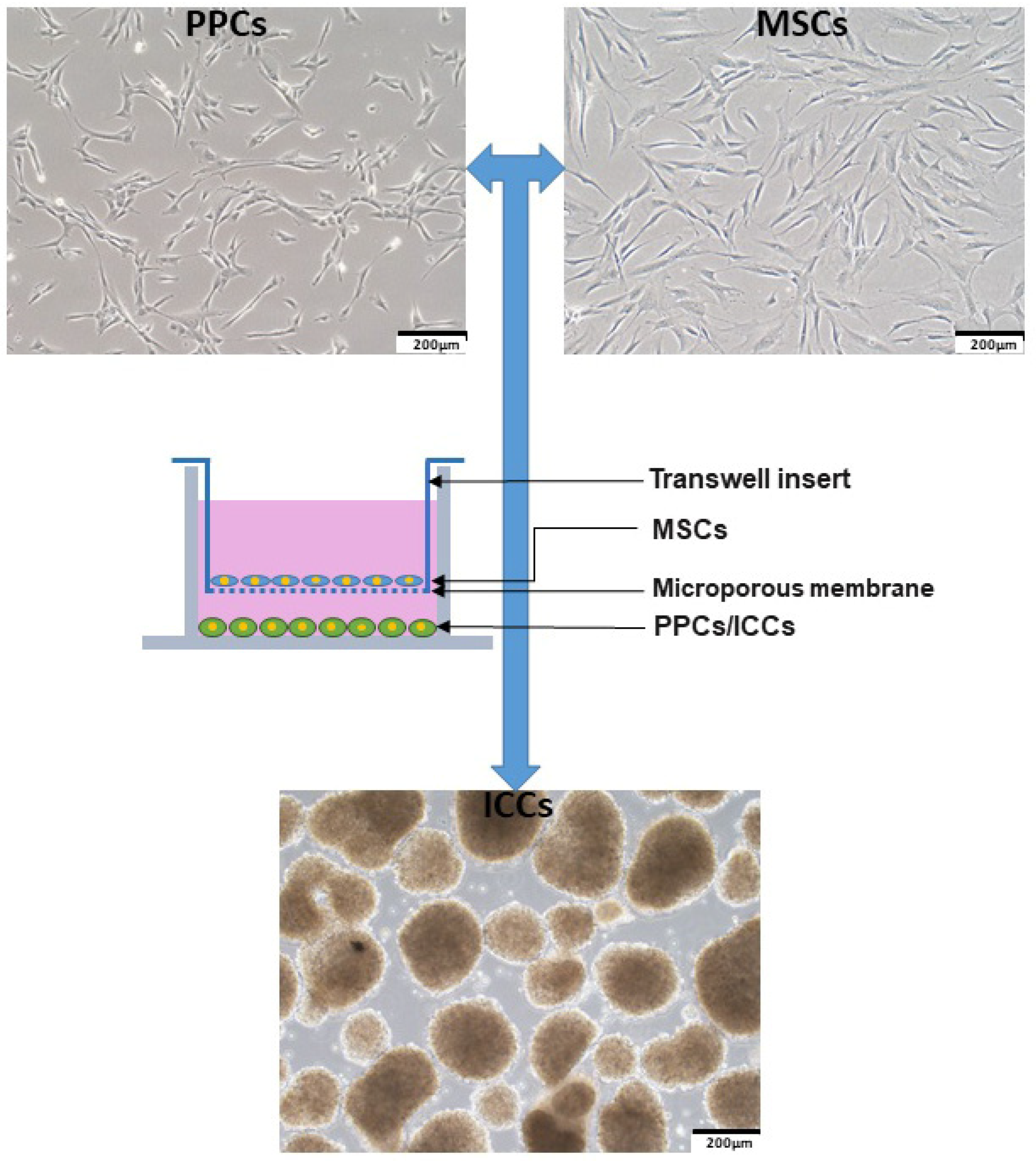
2.1. Human Fetal Bone Marrow-Derived MSCs Enhances PPC Proliferation
2.2. IGF1 is Involved in MSC-Induced PPC Proliferation
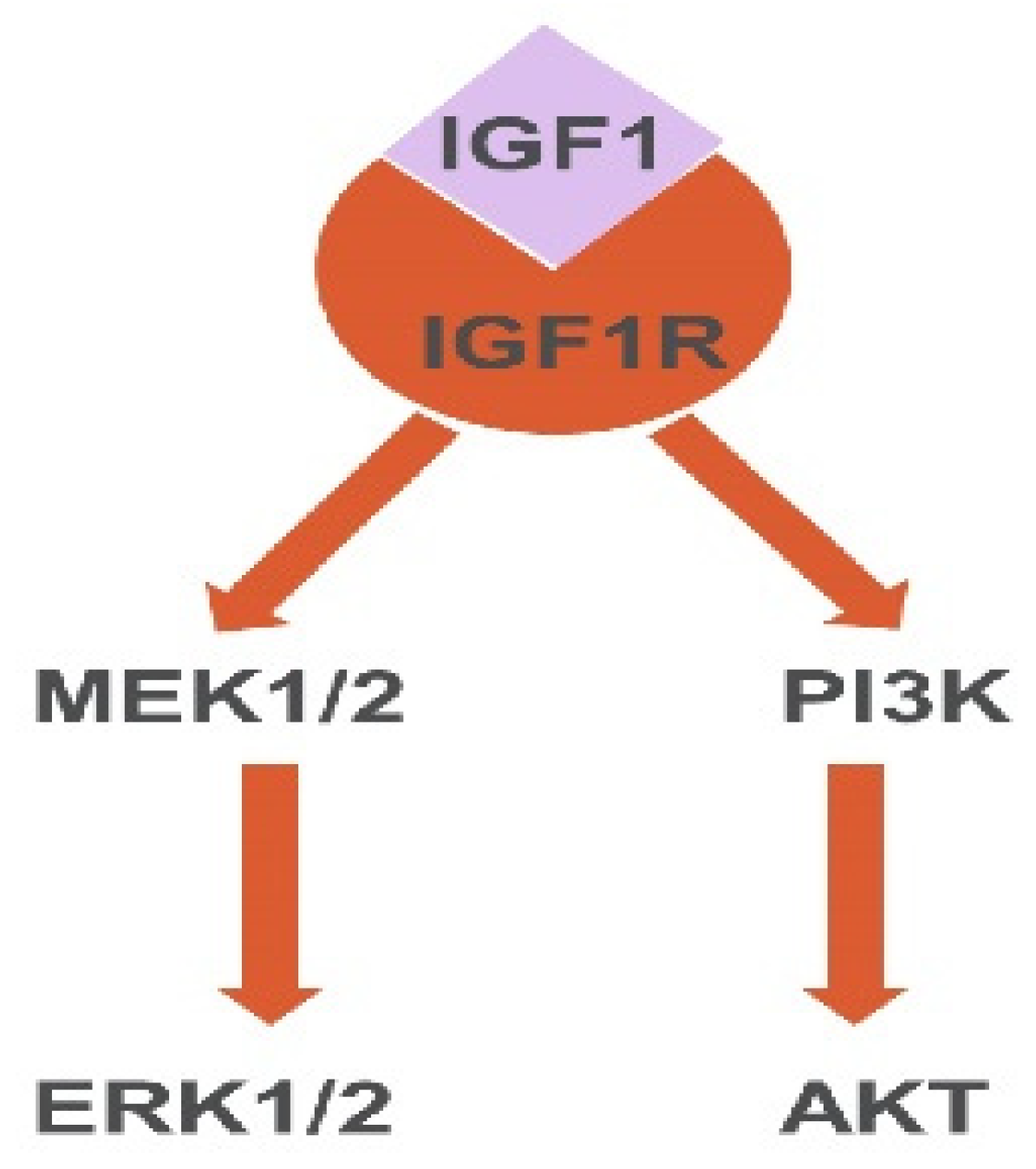
2.3. IGF1 Activates Akt and ERK in MSC-Induced PPC Proliferation
2.4. Human Fetal Bone Marrow-Derived MSCs Promote PPC Differentiation into ICCs
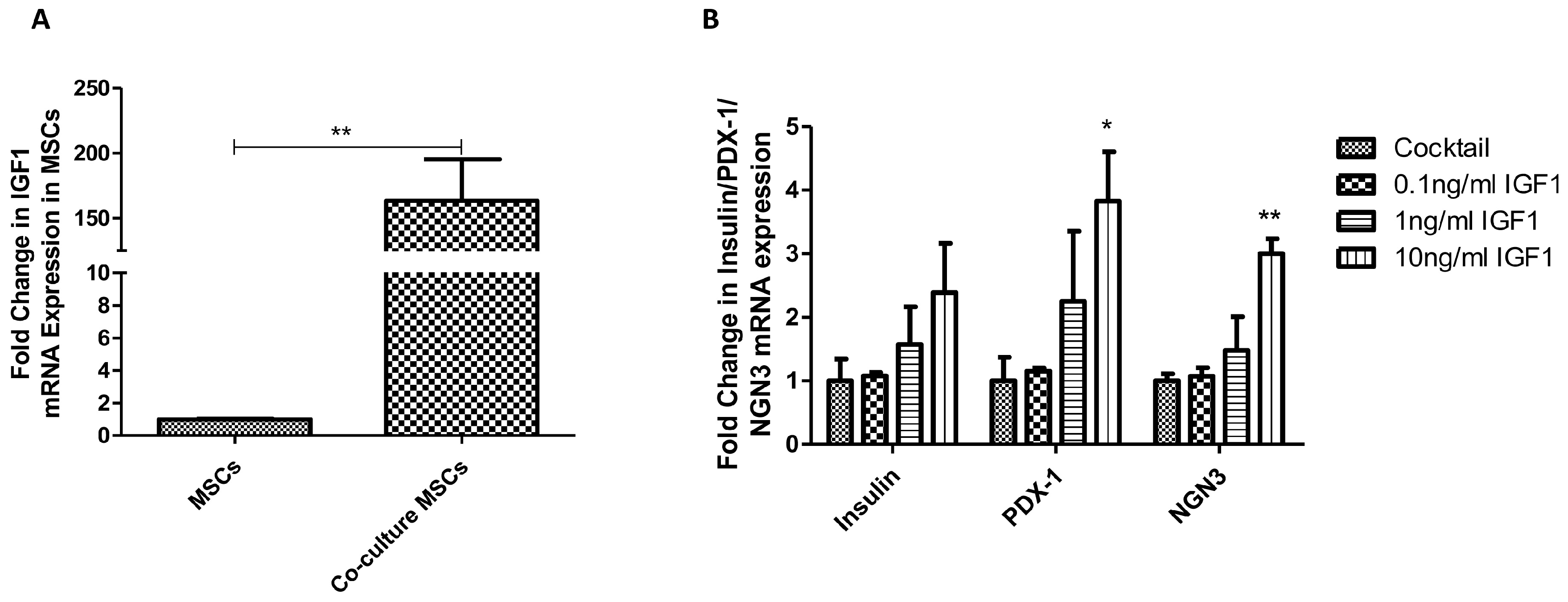
2.5. IGF1 is Involved in MSC Co-Culture Induced PPC Differentiation
2.6. PPC-Conditioned Medium Enhances Human Fetal Bone Marrow-Derived MSC Angiogenic Factor Expression
2.7. MSCs Co-Transplantation Enhances the Engrafted Function of ICCs
3. Discussion
4. Materials and Methods
4.1. Human Ethics for the Use of Human Samples
4.2. Cell Culture of MSCs, PPCs, and ICCs
4.3. Assessment of PPC Proliferation and Apoptosis
4.4. Analysis of mRNA Levels by Quantitative Real-Time PCR
4.5. Measurement of Insulin Content
4.6. Western Blot Analysis
4.7. Immunofluorescent Staining
4.8. Transplantation of ICCs and Blood Glucose Homeostasis Measurement
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PPCs | Pancreatic progenitor cells |
| ICCs | Islet-like cell clusters |
| MSCs | Mesenchymal stem cells |
| IGF1 | Insulin-like growth factor 1 |
| IGF1R | Insulin-like growth factor 1 receptor |
| DM | Diabetes mellitus |
| T1DM | Type 1 diabetes mellitus |
| T2DM | Type 2 diabetes mellitus |
| MSCs-CM | MSC-conditioned medium |
| PPCs-CM | PPC-conditioned medium |
| PPP | Picropodophyllin |
| α-MEM | Alpha-minimum essential medium |
| RPMI | Roswell Park Memorial Institute |
| BSA | Bovine serum albumin |
| FBS | Fetal bovine serum |
| PCR | Polymerase chain reaction |
| PBS | Phosphate-buffered saline |
| PI3K | Phosphoinositide 3-kinases |
| MAPK | Mitogen-activated protein kinase |
| Akt | Protein kinase B |
| ERK | Extracellular signal-regulated kinases |
| MEK | Mitogen-activated protein kinase |
| p- | phospho |
| Bcl-2 | B-cell lymphoma 2 |
| BAX | BCL2 Associated X |
| PDK1 | Phosphoinositide-dependent kinase-1 |
| VEGF | Vascular endothelial growth factor |
| VEGFR | Vascular endothelial growth factor receptor |
| TGF | Transforming growth factor |
| TGF-β | Transforming growth factor-β |
| IL-6 | Interleukin-6 |
| NGN3 | Neurogenin-3 |
| NEUROD | Neurogenic differentiation |
| NKX2.2 | NK2 homeobox 2 |
| NKX6.1 | NK6 homeobox 1 |
| PDX1 | Pancreas and duodenal homeobox gene 1 |
| ISL-1 | ISL LIM homeobox 1 |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| MIP-2 | Macrophage inflammatory protein 2 |
| IPGTT | Intraperitoneal glucose tolerance test |
| STZ | Streptozocin |
| EGF | Epidermal growth factor |
| HGF | Hepatocyte growth factor |
| KGF | Keratinocyte growth factor |
| Ang 2 | Angiotensin 2 |
| sTNFR1 | Soluble tumor necrosis factor receptor 1 |
| ECM | Extracellular matrix |
| TNF-α | Tumor necrosis factor-alpha |
| ESCs | Embryonic stem cells |
References
- Eventov-Friedman, S.; Tchorsh, D.; Katchman, H.; Shezen, E.; Aronovich, A.; Hecht, G.; Dekel, B.; Rechavi, G.; Blazar, B.R.; Feine, I.; et al. Embryonic Pig Pancreatic Tissue Transplantation for the Treatment of Diabetes. PLoS Med. 2006, 3, e215. [Google Scholar] [CrossRef] [PubMed]
- Naftanel, M.A.; Harlan, D.M. Pancreatic islet transplantation. PLoS Med. 2004, 1, e58. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.; Hagopian, W.A.; Gitelman, S.; Masharani, U.; Cavaghan, M.; Rother, K.I.; Donaldson, D.; Harlan, D.M.; Bluestone, J.; Herold, K.C. Insulin Secretion in Type 1 Diabetes. Diabetes 2004, 53, 426–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, K.K.; Liang, J.; Ma, M.T.; Leung, P.S. Angiotensin II Type 2 Receptor Is Critical for the Development of Human Fetal Pancreatic Progenitor Cells into Islet-like Cell Clusters and Their Potential for Transplantation. Stzem Cells 2012, 30, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Krebsbach, P.; Kuznetsov, S.; Bianco, P.; Robey, P.G. Bone Marrow Stromal Cells: Characterization and Clinical Application. Crit. Rev. Oral Biol. Med. 1999, 10, 165–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parekkadan, B.; Milwid, J.M. Mesenchymal Stem Cells as Therapeutics. Annu. Rev. Biomed. Eng. 2010, 12, 87–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busser, H.; Najar, M.; Raicevic, G.; Pieters, K.; Pombo, R.V.; Philippart, P.; Meuleman, N.; Bron, D.; Lagneaux, L. Isolation and Characterization of Human Mesenchymal Stromal Cell Subpopulations: Comparison of Bone Marrow and Adipose Tissue. Stem Cells Dev. 2015, 24, 2142–2157. [Google Scholar] [CrossRef] [PubMed]
- Moroni, L.; Fornasari, P.M. Human mesenchymal stem cells: A bank perspective on the isolation, characterization and potential of alternative sources for the regeneration of musculoskeletal tissues. J. Cell. Physiol. 2013, 228, 680–687. [Google Scholar] [CrossRef]
- Bonab, M.M.; Alimoghaddam, K.; Talebian, F.; Ghaffari, S.H.; Ghavamzadeh, A.; Nikbin, B. Aging of mesenchymal stem cell in vitro. BMC Cell Biol. 2006, 7, 14. [Google Scholar] [CrossRef]
- Gao, F.; Chiu, S.M.; Motan, D.A.L.; Zhang, Z.; Chen, L.; Ji, H.L.; Tse, H.F.; Fu, Q.L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef]
- Rasmusson, I.; Ringden, O.; Sundberg, B.; Le Blanc, K. Mesenchymal stem cells inhibit the formation of cytotoxic T lymphocytes, but not activated cytotoxic T lymphocytes or natural killer cells. Transplantion 2003, 76, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Papazova, D.A.; Oosterhuis, N.R.; Gremmels, H.; Van Koppen, A.; Joles, J.A.; Verhaar, M.C. Cell-based therapies for experimental chronic kidney disease: A systematic review and meta-analysis. Dis. Models Mech. 2015, 8, 281–293. [Google Scholar] [CrossRef]
- Riecke, J.; Johns, K.M.; Cai, C.; Vahidy, F.S.; Parsha, K.; Furr-Stimming, E.; Schiess, M.; Savitz, S.I. A Meta-Analysis of Mesenchymal Stem Cells in Animal Models of Parkinson’s Disease. Stem Cells Dev. 2015, 24, 2082–2090. [Google Scholar] [CrossRef]
- Zhang, L.B.; He, M. Effect of mesenchymal stromal (stem) cell (MSC) transplantation in asthmatic animal models: A systematic review and meta-analysis. Pulm. Pharmacol. Ther. 2019, 54, 39–52. [Google Scholar] [CrossRef]
- Hatzistergos, K.E.; Quevedo, H.; Oskouei, B.N.; Hu, Q.; Feigenbaum, G.S.; Margitich, I.S.; Mazhari, R.; Boyle, A.J.; Zambrano, J.P.; Rodriguez, J.E.; et al. Bone marrow mesenchymal stem cells stimulate cardiac stem cell proliferation and differentiation. Circ. Res. 2010, 107, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.N.; Da Costa e Silva, B.F.B.; Soares, J.C.; Carvalho, A.F.; Quevedo, J. Role of trophic factors GDNF, IGF-1 and VEGF in major depressive disorder: A comprehensive review of human studies. J. Affect. Disord. 2016, 197, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renehan, A.G.; Zwahlen, M.; Minder, C.; O’Dwyer, S.T.; Shalet, S.M.; Egger, M. Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: Systematic review and meta-regression analysis. Lancet 2004, 363, 1346–1353. [Google Scholar] [CrossRef]
- An, C.; Cheng, Y.; Yuan, Q.; Li, J. IGF-1 and BMP-2 Induces Differentiation of Adipose-Derived Mesenchymal Stem Cells into Chondrocytes-Like Cells. Ann. Biomed. Eng. 2010, 38, 1647–1654. [Google Scholar] [CrossRef]
- Huat, T.J.; Khan, A.A.; Pati, S.; Mustafa, Z.; Abdullah, J.M.; Jaafar, H. IGF-1 enhances cell proliferation and survival during early differentiation of mesenchymal stem cells to neural progenitor-like cells. BMC Neurosci. 2014, 15, 91. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tredget, E.E.; Wu, P.Y.G.; Wu, Y. Paracrine Factors of Mesenchymal Stem Cells Recruit Macrophages and Endothelial Lineage Cells and Enhance Wound Healing. PLoS ONE 2008, 3, e1886. [Google Scholar] [CrossRef]
- Liu, J.P.; Baker, J.; Perkins, A.S.; Robertson, E.J.; Efstratiadis, A. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (Igf1r). Cell 1993, 75, 59–72. [Google Scholar] [CrossRef]
- Lingohr, M.K.; Dickson, L.M.; McCuaig, J.F.; Hugl, S.R.; Twardzik, D.R.; Rhodes, C.J. Activation of IRS-2—Mediated Signal Transduction by IGF-1, but not TGF-or EGF, Augments Pancreatic-Cell Proliferation. Diabetes 2002, 51, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Hugl, S.R.; White, M.F.; Rhodes, C.J. Insulin-like Growth Factor I (IGF-I)-stimulated Pancreatic-cell growth is glucose-dependent: synergistic activation of insulin receptor substrate-mediated signal transduction pathways by glucose and IGF-I in INS-1 cells. J. Biol. Chem. 1998, 273, 17771–17779. [Google Scholar] [CrossRef] [PubMed]
- Withers, D.J.; Burks, D.J.; Towery, H.H.; Altamuro, S.L.; Flint, C.L.; White, M.F. Irs-2 coordinates Igf-1 receptor-mediated β-cell development and peripheral insulin signalling. Nat. Genet. 1999, 23, 32–40. [Google Scholar] [CrossRef] [PubMed]
- George, M.; Ayuso, E.; Casellas, A.; Costa, C.; Devedjian, J.C.; Bosch, F. β cell expression of IGF-I leads to recovery from type 1 diabetes. J. Clin. Investig. 2002, 109, 1153–1163. [Google Scholar] [CrossRef]
- Landsman, L.; Nijagal, A.; Whitchurch, T.J.; Vanderlaan, R.L.; Zimmer, W.E.; MacKenzie, T.C.; Hebrok, M. Pancreatic Mesenchyme Regulates Epithelial Organogenesis throughout Development. PLoS Biol. 2011, 9, e1001143. [Google Scholar] [CrossRef] [PubMed]
- Duvillié, B.; Attali, M.; Bounacer, A.; Ravassard, P.; Basmaciogullari, A.; Scharfmann, R. The Mesenchyme Controls the Timing of Pancreatic-Cell Differentiation. Diabetes 2006, 55, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Dionne, K.E.; Colton, C.K.; Lyarmush, M. Effect of Hypoxia on Insulin Secretion by Isolated Rat and Canine Islets of Langerhans. Diabetes 1993, 42, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Kusminski, C.M.; Shetty, S.; Orci, L.; Unger, R.H.; Scherer, P.E. Diabetes and apoptosis: Lipotoxicity. Apoptosis 2009, 14, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, T.; Oh, S.H.; Pi, L.; Hatch, H.M.; Shupe, T.; Petersen, B.E. Teratoma Formation Leads to Failure of Treatment for Type I Diabetes Using Embryonic Stem Cell-Derived Insulin-Producing Cells. Am. J. Pathol. 2005, 166, 1781–1791. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo, M.; Banerjee, I. Endothelial Cell Co-culture Mediates Maturation of Human Embryonic Stem Cell to Pancreatic Insulin Producing Cells in a Directed Differentiation Approach. J. Vis. Exp. 2012, e3759. [Google Scholar] [CrossRef] [PubMed]
- Goers, L.; Freemont, P.; Polizzi, K.M. Co-culture systems and technologies: Taking synthetic biology to the next level. J. R. Soc. Interface 2014, 11, 20140065. [Google Scholar] [CrossRef] [PubMed]
- De Lima, M.; Mcniece, I.; Robinson, S.N.; Munsell, M.; Eapen, M.; Horowitz, M.; Alousi, A.; Saliba, R.; McMannis, J.D.; Kaur, I.; et al. Cord-Blood Engraftment with Ex Vivo Mesenchymal-Cell Coculture. N. Engl. J. Med. 2012, 367, 2305–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volarevic, V.; Arsenijevic, N.; Lukic, M.L.; Stojkovic, M. Mesenchymal stem cell treatment of the complications of diabetes mellitus. Stem Cells 2011, 29, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Seeberger, K.; Salama, B.; Mulet-Sierra, A.; Kuppan, P.; Hayward, J.A.; Ellis, C.E.; Lee, T.; Adesida, A.; Korbutt, G.S. Cotransplantation of Mesenchymal Stem Cells With Neonatal Porcine Islets Improve Graft Function in Diabetic Mice. Diabetes 2017, 66, 1312–1321. [Google Scholar] [Green Version]
- Kerby, A.; Jones, E.S.; Jones, P.M.; King, A.J. Co-transplantation of islets with mesenchymal stem cells in microcapsules demonstrates graft outcome can be improved in an isolated-graft model of islet transplantation in mice. Cytotherapy 2013, 15, 192–200. [Google Scholar] [CrossRef]
- Figliuzzi, M.; Cornolti, R.; Perico, N.; Rota, C.; Morigi, M.; Remuzzi, G.; Remuzzi, A.; Benigni, A. Bone Marrow–Derived Mesenchymal Stem Cells Improve Islet Graft Function in Diabetic Rats. Transplant. Proc. 2009, 41, 1797–1800. [Google Scholar] [CrossRef] [PubMed]
- Rackham, C.L.; Chagastelles, P.C.; Nardi, N.B.; Hauge-Evans, A.C.; Jones, P.M.; King, A.J.F. Co-transplantation of mesenchymal stem cells maintains islet organisation and morphology in mice. Diabetologia 2011, 54, 1127–1135. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Itakura, S.; Todorov, I.; Rawson, J.; Asari, S.; Shintaku, J.; Nair, I.; Ferreri, K.; Kandeel, F.; Mullen, Y. Mesenchymal stem cell and islet co-transplantation promotes graft revascularization and function. Transplantation 2010, 89, 1438–1445. [Google Scholar] [CrossRef]
- Sakata, N.; Chan, N.K.; Chrisler, J.; Obenaus, A.; Hathout, E. Bone Marrow Cell Co-Transplantation with Islets Improves their Vascularization and Function. Transplantation 2010, 89, 686–693. [Google Scholar] [CrossRef]
- Sunderkötter, C.; Steinbrink, K.; Goebeler, M.; Bhardwaj, R.; Sorg, C. Macrophages and angiogenesis. J. Leukoc. Biol. 1994, 55, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Nör, J.E.; Christensen, J.; Mooney, D.J.; Polverini, P.J. Vascular Endothelial Growth Factor (VEGF)-Mediated Angiogenesis Is Associated with Enhanced Endothelial Cell Survival and Induction of Bcl-2 Expression. Am. J. Pathol. 1999, 154, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Yagi, H.; Soto-Gutierrez, A.; Navarro-Alvarez, N.; Nahmias, Y.; Goldwasser, Y.; Kitagawa, Y.; Tilles, A.W.; Tompkins, R.G.; Parekkadan, B.; Yarmush, M.L. Reactive Bone Marrow Stromal Cells Attenuate Systemic Inflammation via sTNFR. Mol. Ther. 2010, 18, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Daley, W.P.; Peters, S.B.; Larsen, M. Extracellular matrix dynamics in development and regenerative medicine. J. Cell Sci. 2008, 121, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Hoben, G.M.; Koay, E.J.; Athanasiou, K.A. Fibrochondrogenesis in Two Embryonic Stem Cell Lines: Effects of Differentiation Timelines. Stem Cells 2008, 26, 422–430. [Google Scholar] [CrossRef] [PubMed]
- McBride, S.H.; Tate, M.L.K. Modulation of Stem Cell Shape and Fate A: The Role of Density and Seeding Protocol on Nucleus Shape and Gene Expression. Tissue Eng. Part A 2008, 14, 1561–1572. [Google Scholar] [CrossRef]
- Schmidt, A.; Ladage, D.; Steingen, C.; Brixius, K.; Schinköthe, T.; Klinz, F.J.; Schwinger, R.H.; Mehlhorn, U.; Bloch, W. Mesenchymal stem cells transmigrate over the endothelial barrier. Eur. J. Cell Biol. 2006, 85, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Van Haeften, T.W.; Twickler, T.B. Insulin-like growth factors and pancreas beta cells. Eur. J. Clin. Investig. 2004, 34, 249–255. [Google Scholar] [CrossRef]
- Holt, R. Fetal programming of the growth hormone–insulin-like growth factor axis. Trends Endocrinol. Metab. 2002, 13, 392–397. [Google Scholar] [CrossRef]
- Rhodes, C.J.; White, M.F. Molecular insights into insulin action and secretion. Eur. J. Clin. Investig. 2002, 32, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Pratsinis, H.; Kletsas, D. PDGF, bFGF and IGF-I stimulate the proliferation of intervertebral disc cells in vitro via the activation of the ERK and Akt signaling pathways. Eur. Spine J. 2007, 16, 1858–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Montagne, K.; Nishihara, A.; Watabe, T.; Miyazono, K. BMPs promote proliferation and migration of endothelial cells via stimulation of VEGF-A/VEGFR2 and Angiopoietin-1/Tie2 signalling. J. Biochem. 2008, 143, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lee, Y.W.; Rui, Y.F.; Cheng, T.Y.; Jiang, X.H.; Li, G. Bone marrow-derived mesenchymal stem cells promote growth and angiogenesis of breast and prostate tumors. Stem Cell Res. Ther. 2013, 4, 70. [Google Scholar] [CrossRef]
- Suen, P.; Zou, C.; Zhang, Y.; Lau, T.K.; Chan, J.; Yao, K.; Leung, P.S. PDZ-domain containing-2 (PDZD2) is a novel factor that affects the growth and differentiation of human fetal pancreatic progenitor cells. Int. J. Biochem. Cell Biol. 2008, 40, 789–803. [Google Scholar] [CrossRef]
- Leung, K.K.; Suen, P.M.; Lau, T.K.; Ko, W.H.; Yao, K.M.; Leung, P.S. PDZ-Domain Containing-2 (PDZD2) Drives the Maturity of Human Fetal Pancreatic Progenitor-Derived Islet-Like Cell Clusters With Functional Responsiveness Against Membrane Depolarization. Stem Cells Dev. 2009, 18, 979–990. [Google Scholar] [CrossRef]
- Ma, M.T.; Leung, K.K.; Tsang, K.S.; Leung, P.S. Reduced immunogenicity of pancreatic progenitor cells derived from first-trimester human fetal pancreas. Int. J. Biochem. Cell Biol. 2011, 43, 812–820. [Google Scholar] [CrossRef]
- Ng, K.Y.; Ma, M.T.; Leung, K.K.; Leung, P.S. Vitamin D and vitamin A receptor expression and the proliferative effects of ligand activation of these receptors on the development of pancreatic progenitor cells derived from human fetal pancreas. Stem Cell Rev. Rep. 2011, 7, 53–63. [Google Scholar] [CrossRef]
- Liang, J.; Ng, K.Y.; Cheng, Q.; Xia, Y.; Wang, C.C.; Leung, P.S. Human Fetal Liver Stromal Cell Co-Culture Enhances the Differentiation of Pancreatic Progenitor Cells into Islet-Like Cell Clusters. Stem Cell Rev. Rep. 2014, 10, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.K.; Sandler, S. Melatonin protects against streptozotocin, but not interleukin-1beta-induced damage of rodent pancreatic beta-cells. J. Pineal Res. 2001, 30, 157–165. [Google Scholar] [CrossRef]
- Suen, P.M.; Li, K.; Chan, J.C.; Leung, P.S. In vivo treatment with glucagon-like peptide 1 promotes the graft function of fetal islet-like cell clusters in transplanted mice. Int. J. Biochem. Cell Biol. 2006, 38, 951–960. [Google Scholar] [CrossRef]
- Cheng, Q.; Law, P.K.; De Gasparo, M.; Leung, P.S. Combination of the dipeptidyl peptidase IV inhibitor LAF237 [(S)-1-[(3-hydroxy-1-adamantyl)ammo]acetyl-2-cyanopyrrolidine] with the angiotensin II type 1 receptor antagonist valsartan [N-(1-oxopentyl)-N-[[2′-(1H-tetrazol-5-yl)-[1,1′-biphenyl]-4-yl]methyl]-L-valine] enhances pancreatic islet morphology and function in a mouse model of type 2 diabetes. J. Pharmacol. Exp. Ther. 2008, 327, 683–691. [Google Scholar] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward (5′-3′) | Reverse (5′-3′) | |
|---|---|---|---|
| VEGF | CTACCTCCACCATGCCAAGT | GCAGTAGCTGCGCTGATAGA | |
| TGF-β | CCCAGCATCTGCAAAGCTC | GTCAATGTACAGCTGCCGCA | |
| IL-6 | GGTACATCCTCGACGGCATCT | GTGCCTCTTTGCTGCTTTCAC | |
| INSULIN | CAGCCTTTGTGAACCAACACC | GGTCTTGGGTGTGTAGAAGAAGC | |
| IGF1 | TGGATGCTCTTCAGTTCGTG | TGGTAGATGGGGGCTGATAC | |
| IGF1R | GCACCATCTTCAAGGGCAATTTG | AGGAAGGACAAGGAGACCAAGG | |
| NGN3 | CGGACCCCATTCTCTCTTCT | ACTTCGTCTTCCGAGGCTCT | |
| NEUROD | TCCAAAATCGAGACTCTGCGC | GCAAAGCGTCTGAACGAAGGA | |
| NKX2.2 | TCTCCTTGGAGTGGCAGATTC | AAACACGGCGTAGAGTTCAGC | |
| NKX6.1 | GACGGGAAGAGAAAACACACG | ACTCTCTGTCATCCCCAACGA | |
| PDX1 | ACTCCACCTTGGGACCTGTTT | TTAAGGTACTCGGCCCAGCTT | |
| ACTIN | TGTCCACCTTCCAGCAGATGT | CGGACTCGTCATACTCCTGCTT | |
| ISL-1 | GATCAAATGCGCCAAGTGCAG | CAGCGGAAACACTCGATGTGA | |
| MIP-2 | CGCCCAAACCGAAGTCAT | GATTTGCCATTTTTCAGCATCTTT | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.Y.; Wu, S.Y.; Leung, P.S. Human Fetal Bone Marrow-Derived Mesenchymal Stem Cells Promote the Proliferation and Differentiation of Pancreatic Progenitor Cells and the Engraftment Function of Islet-Like Cell Clusters. Int. J. Mol. Sci. 2019, 20, 4083. https://doi.org/10.3390/ijms20174083
Li XY, Wu SY, Leung PS. Human Fetal Bone Marrow-Derived Mesenchymal Stem Cells Promote the Proliferation and Differentiation of Pancreatic Progenitor Cells and the Engraftment Function of Islet-Like Cell Clusters. International Journal of Molecular Sciences. 2019; 20(17):4083. https://doi.org/10.3390/ijms20174083
Chicago/Turabian StyleLi, Xing Yu, Shang Ying Wu, and Po Sing Leung. 2019. "Human Fetal Bone Marrow-Derived Mesenchymal Stem Cells Promote the Proliferation and Differentiation of Pancreatic Progenitor Cells and the Engraftment Function of Islet-Like Cell Clusters" International Journal of Molecular Sciences 20, no. 17: 4083. https://doi.org/10.3390/ijms20174083