MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Involvement in the Regulation of Detoxification Mechanisms

 , and
, and
Abstract
:1. Introduction
2. Results
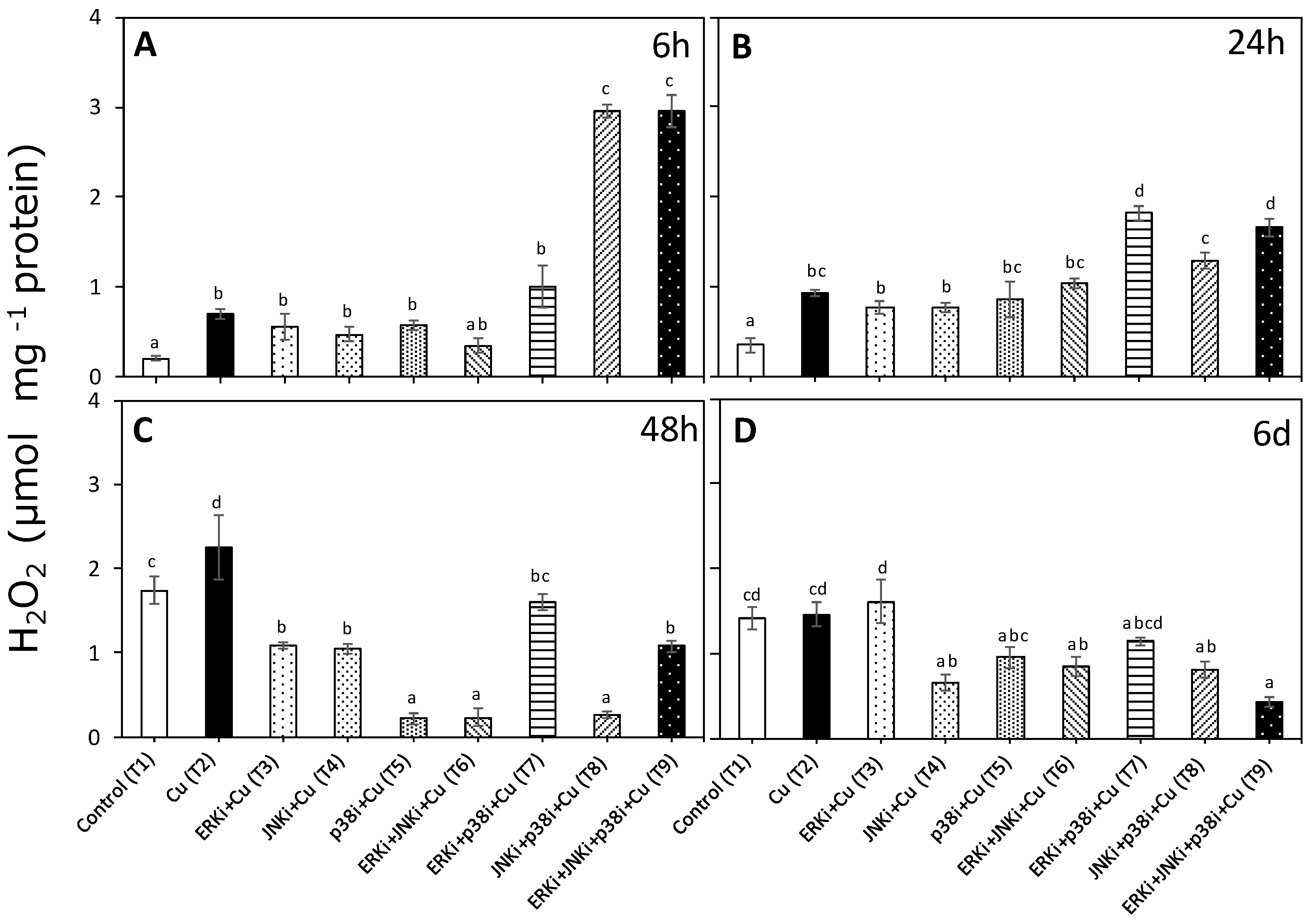
2.1. Hydrogen Peroxide Levels
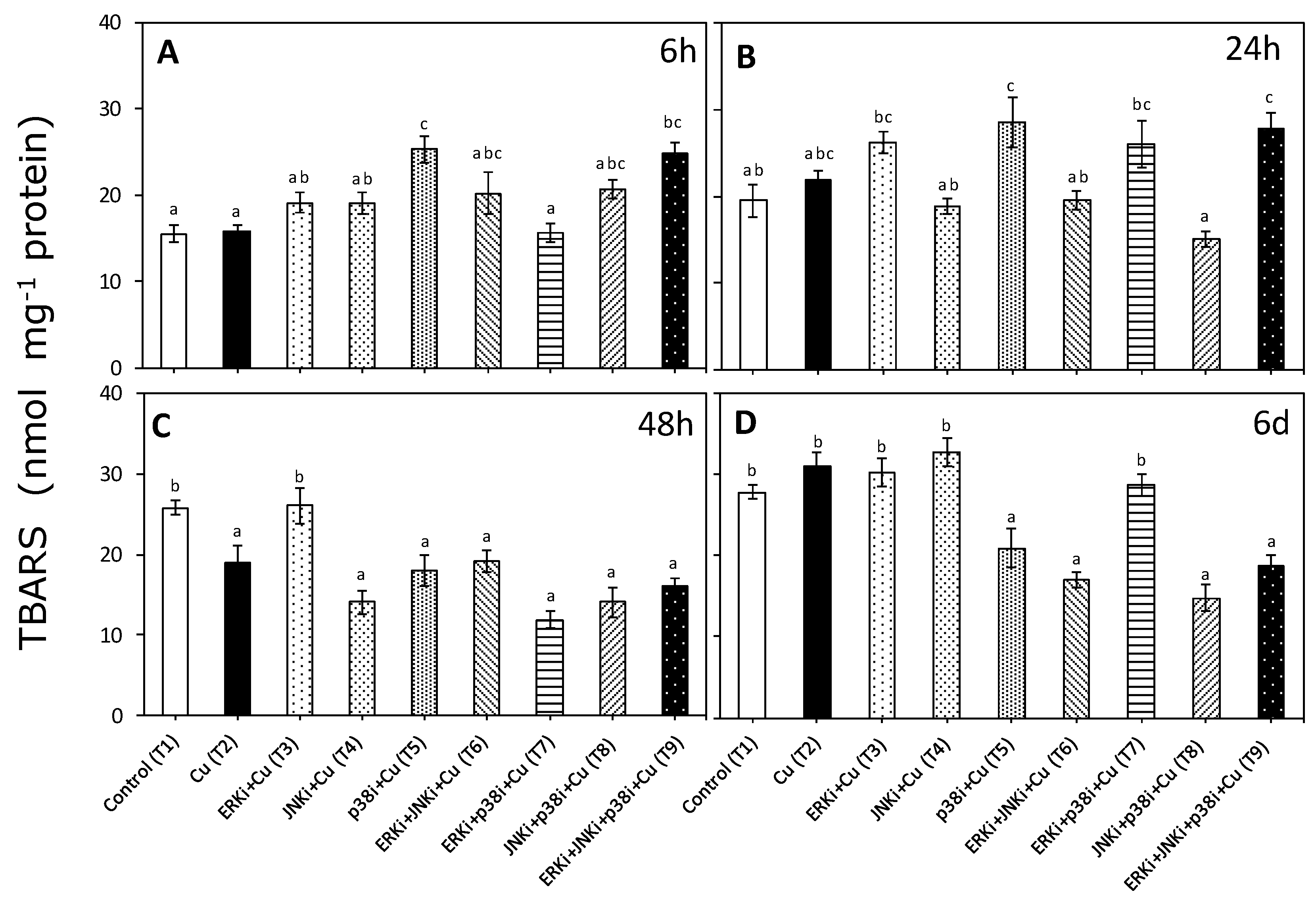
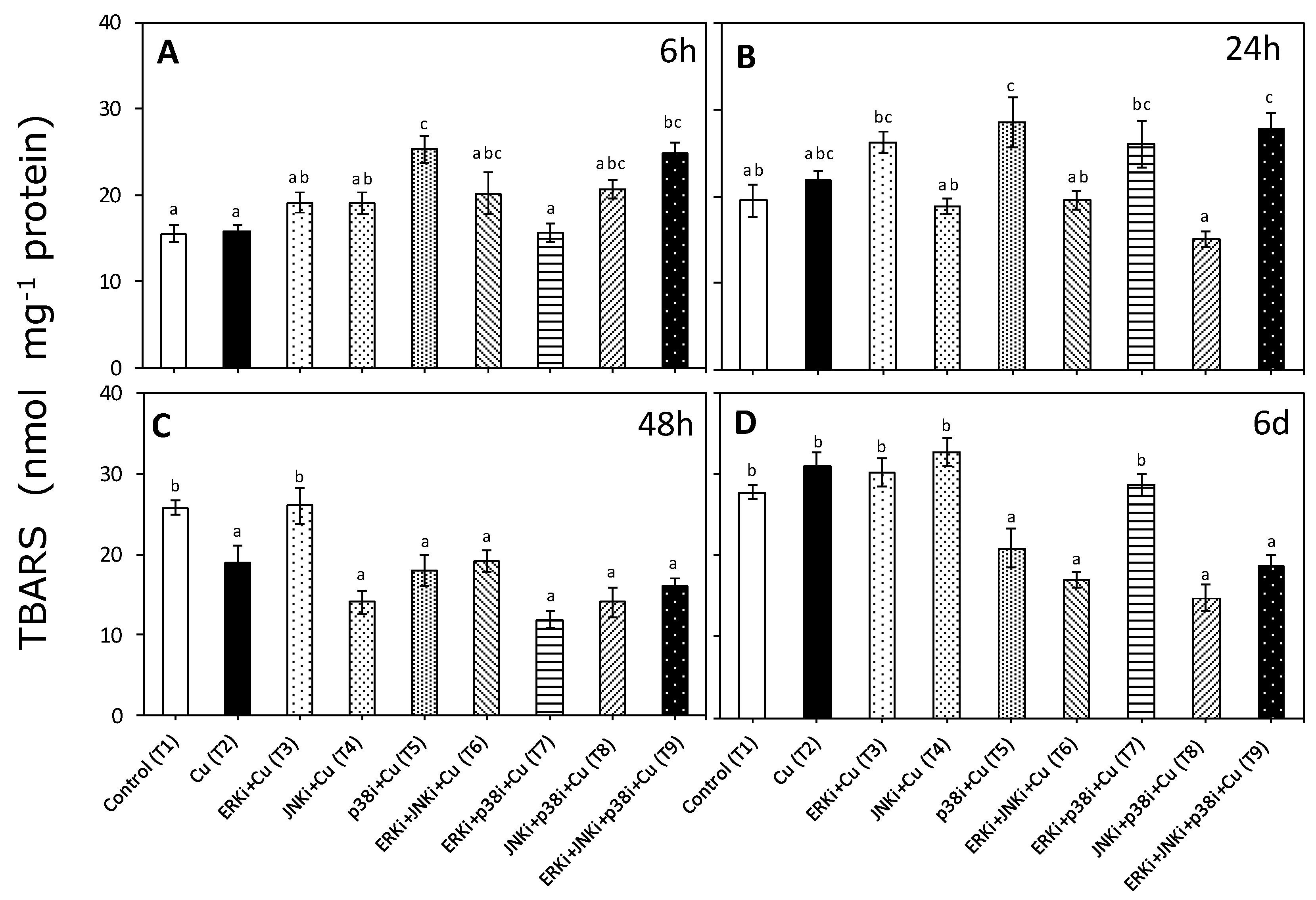
2.2. Lipid Peroxidation
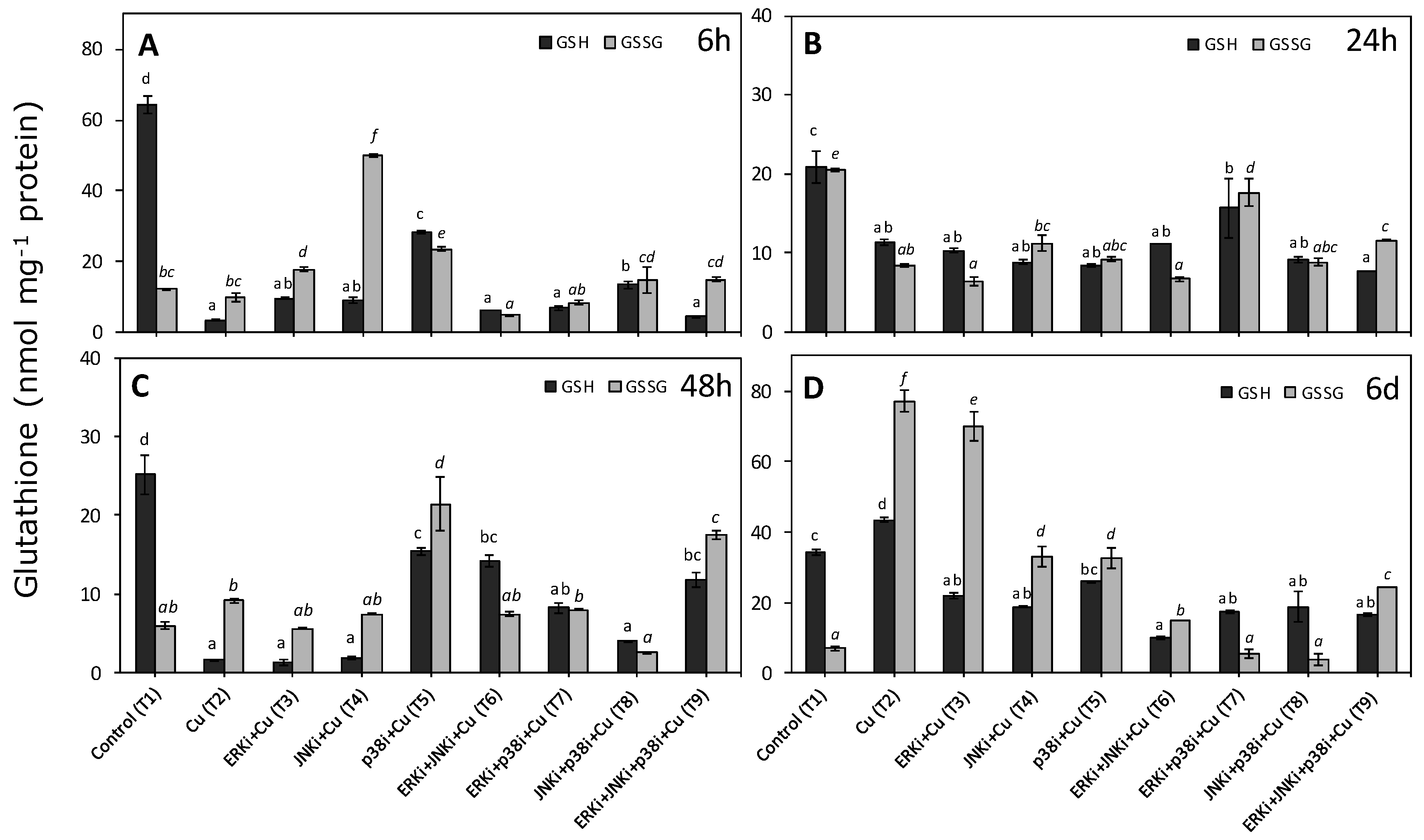
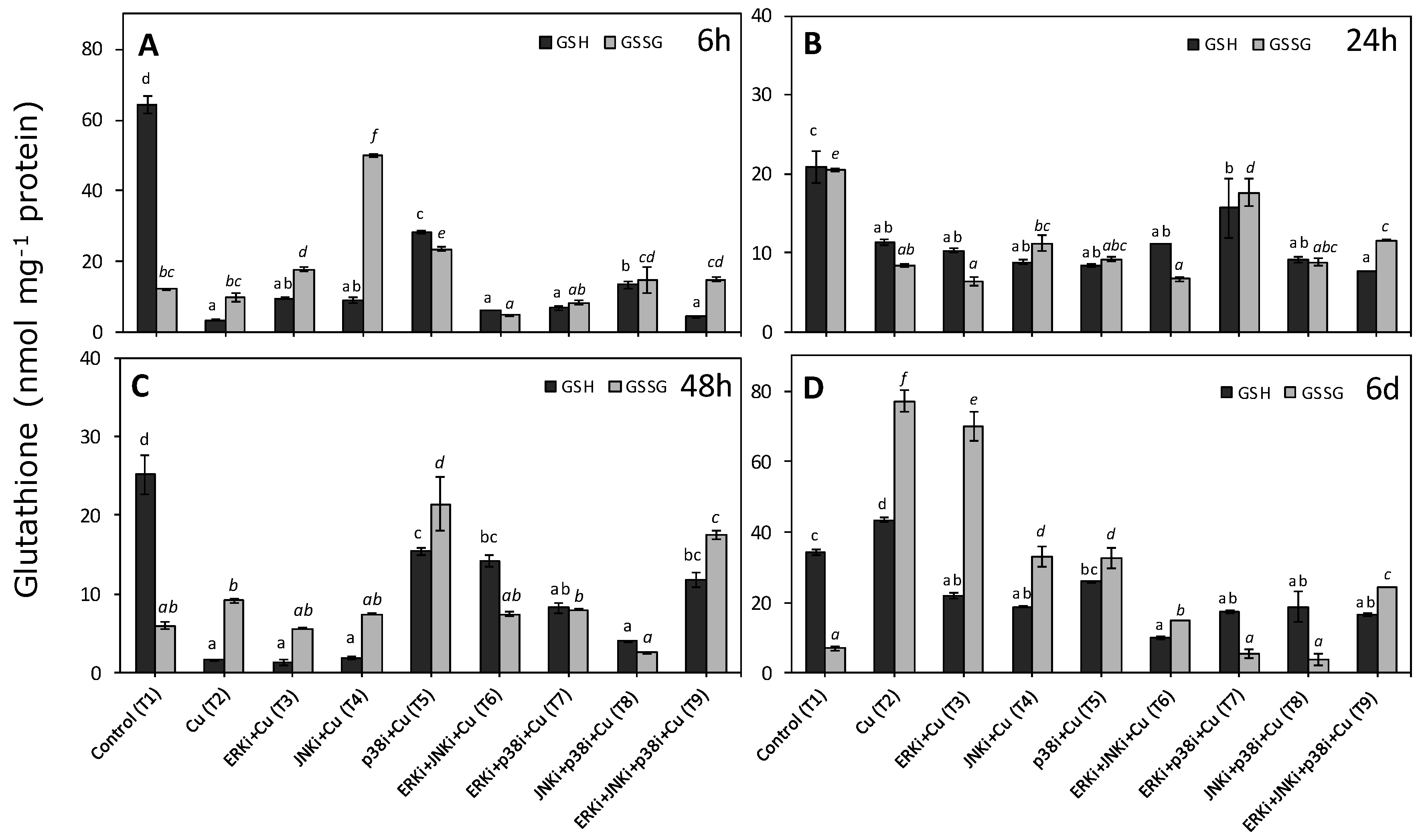
2.3. Levels of Reduced and Oxidised Glutathione
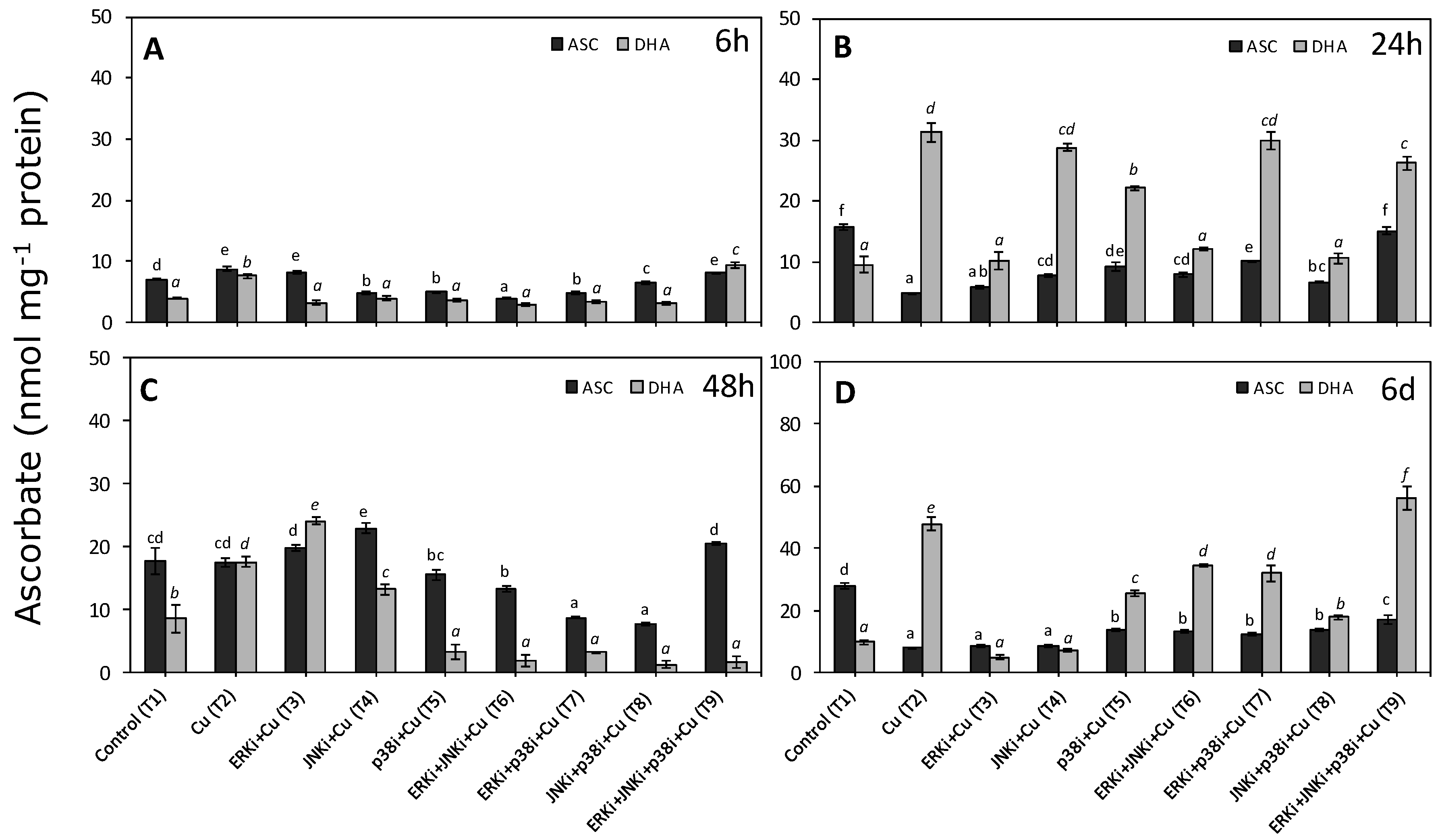
2.4. Concentrations of Reduced and Oxidised Ascorbate
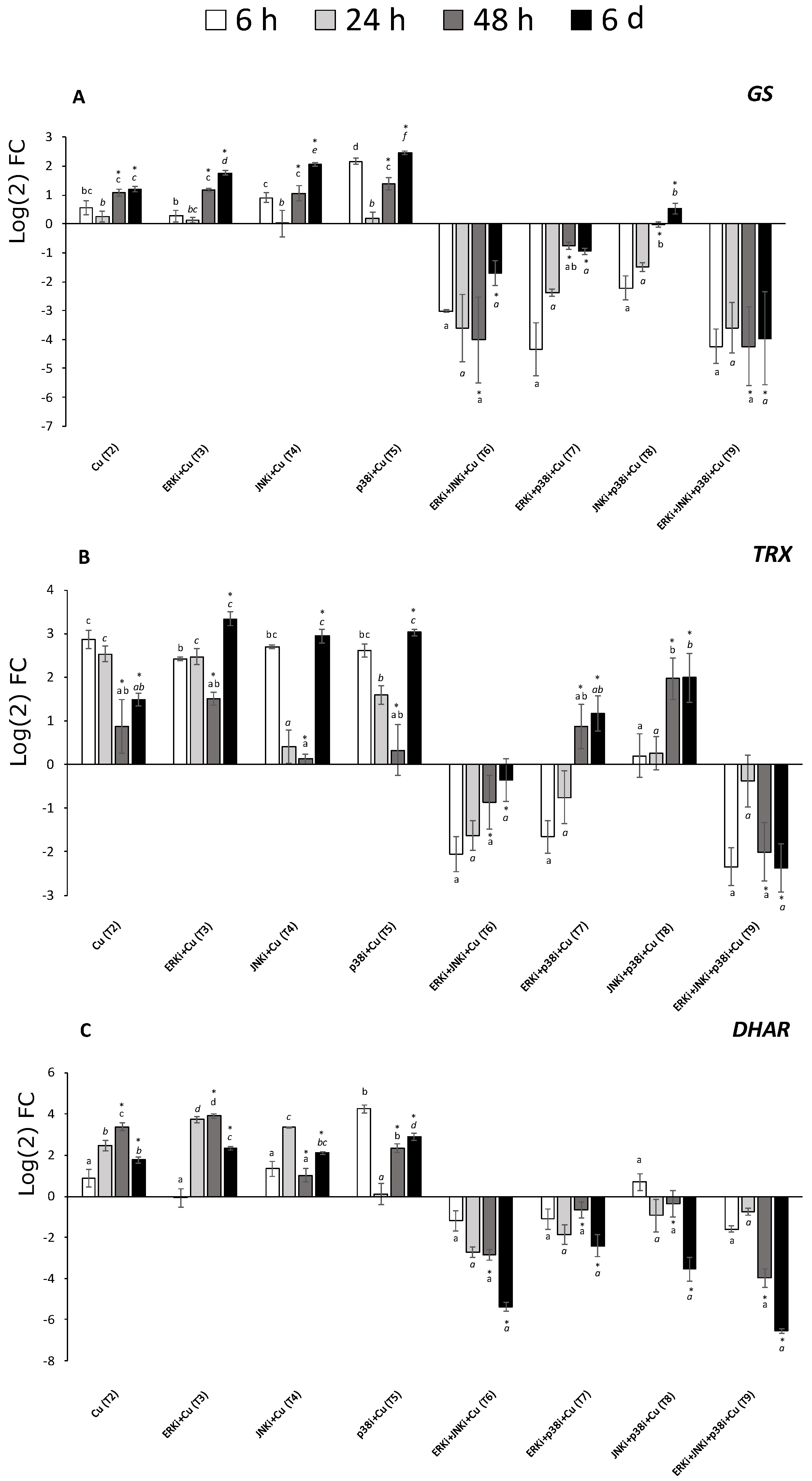
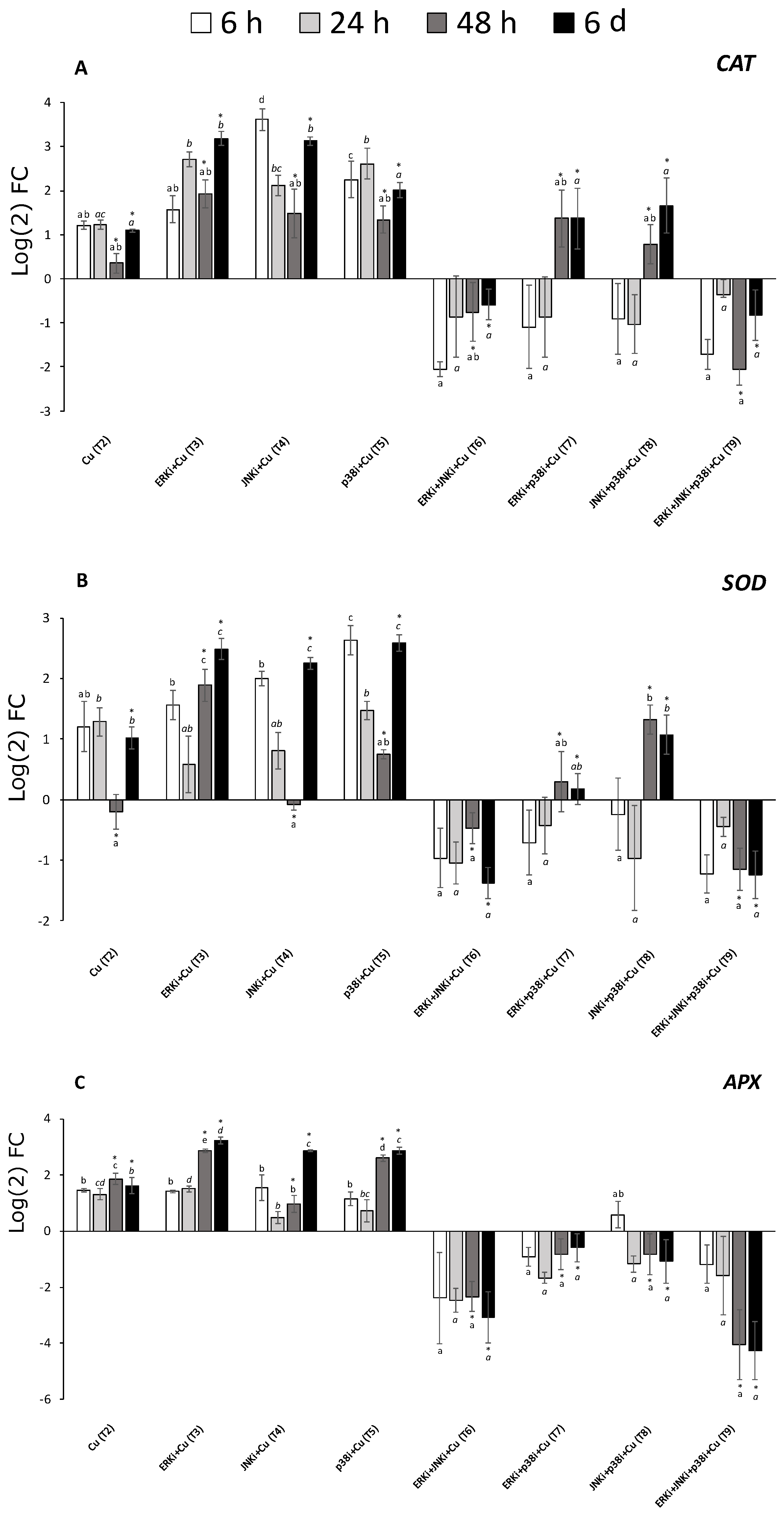
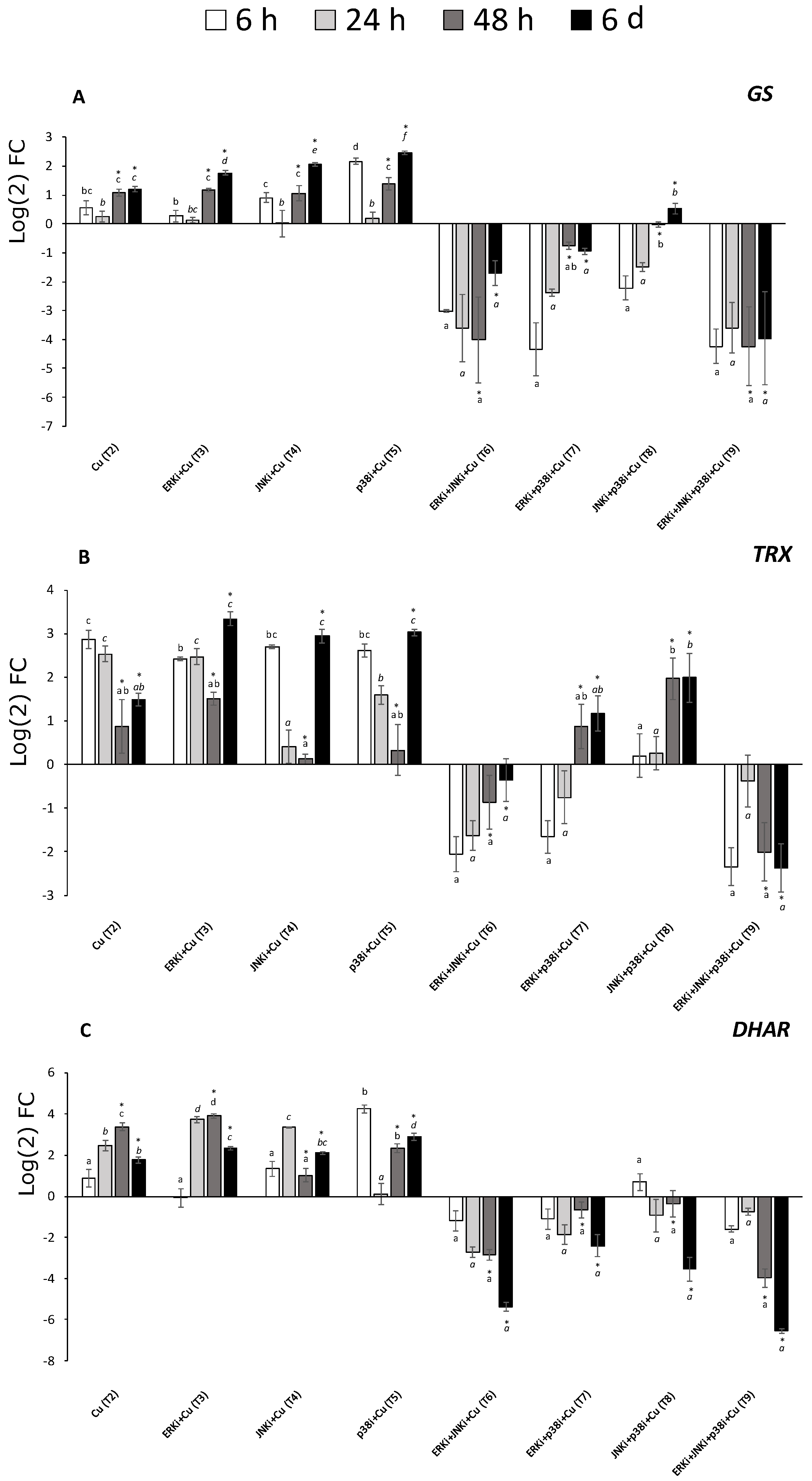
2.5. Gene Expression Profiles
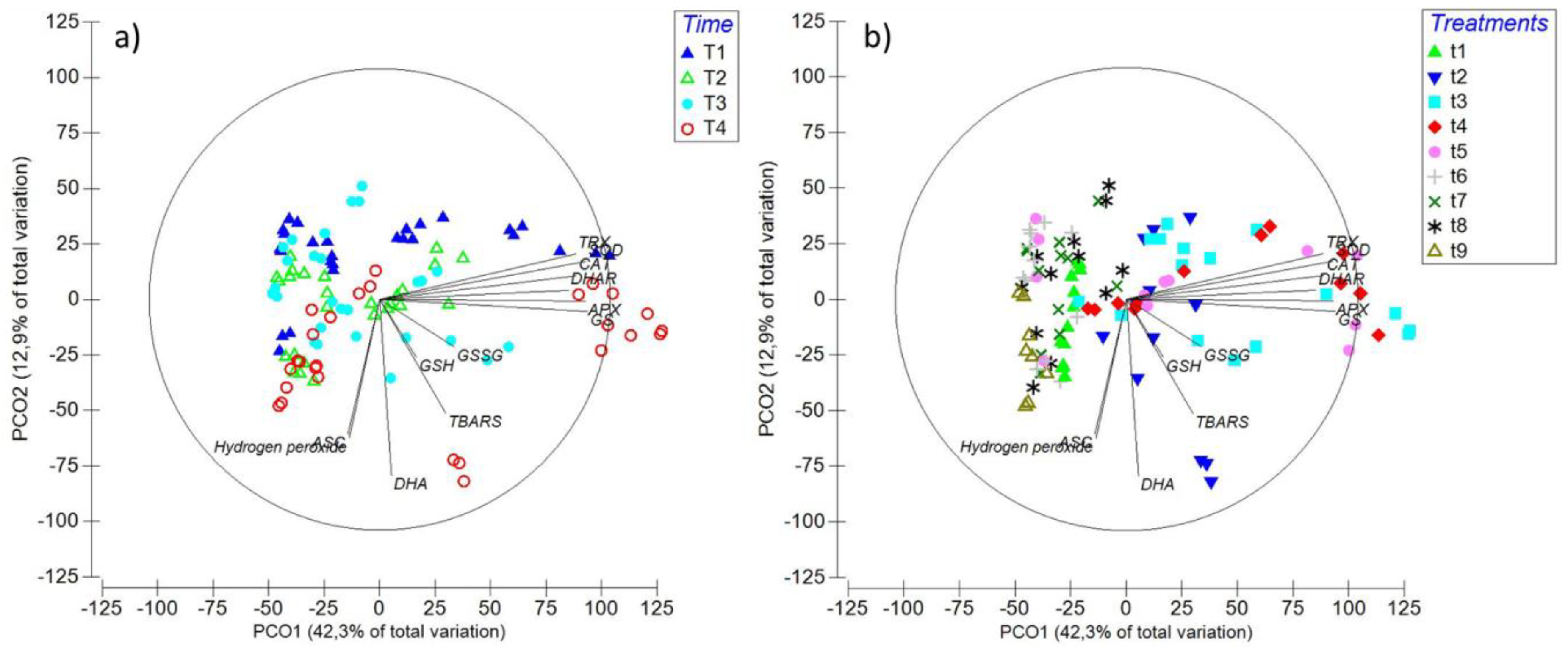
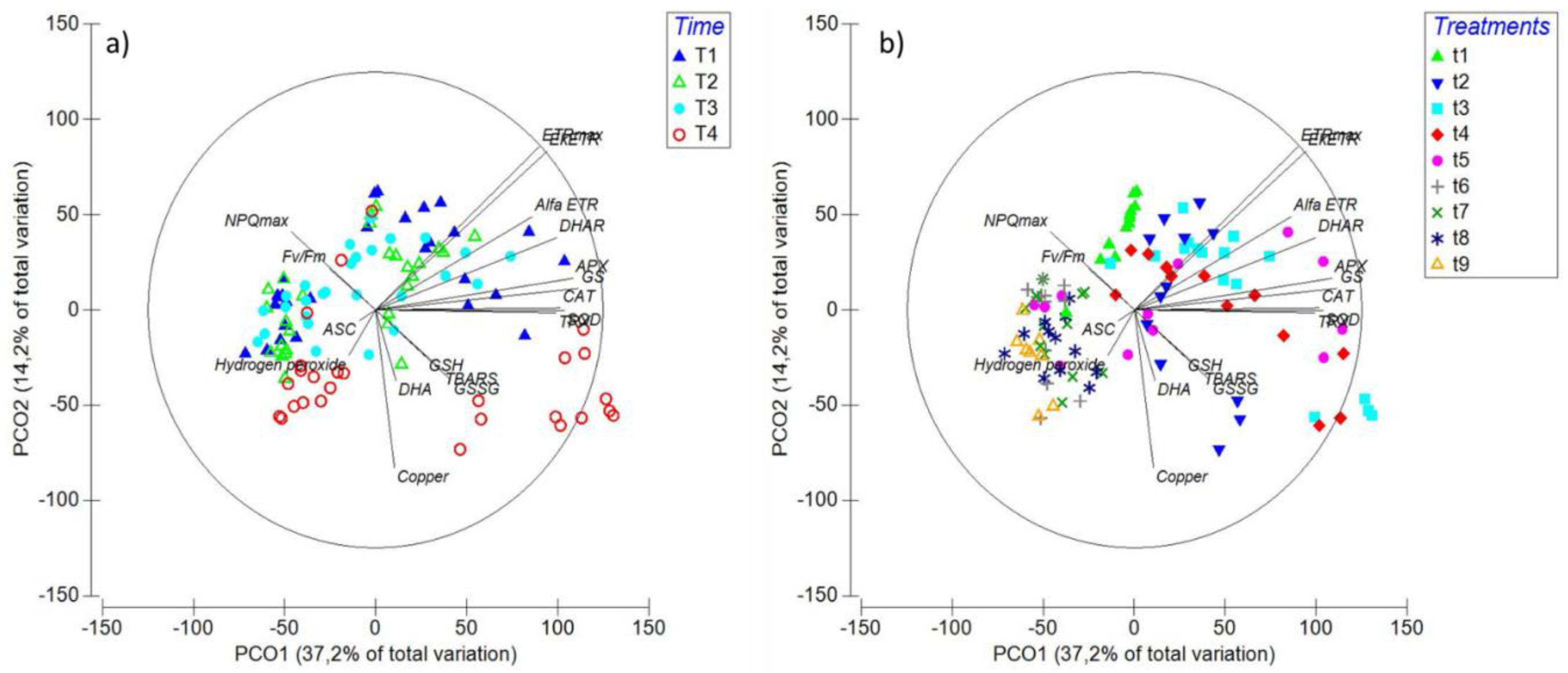
2.6. Principal Coordinate Ordination (PCO) Analysis
3. Discussion
4. Methodology
4.1. Samples Collection and Culture Conditions
4.2. Copper and MAPK Inhibitors Treatments
4.3. Hydrogen Peroxide (H2O2) Quantification
4.4. Determination of Lipid Peroxidation
4.5. Oxidised and Reduced Glutathione
4.6. Oxidised and Reduced Ascorbate
4.7. RNA Extraction
4.8. cDNA Synthesis and qPCR
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MAPK | mitogen activated protein kinases |
| ERK | Extracellular Signal Regulated Kinases |
| JNK | c-Jun N-terminal kinases |
| p38 | cytokinin specific binding protein |
| ROS | reactive oxygen species |
| GSH | reduced glutathione |
| GSSG | oxidised glutathione |
| ASC | reduced ascorbate |
| DHA | dehydroascorbate (form of oxidised ascorbate) |
| CAT | catalase |
| SOD | superoxide dismutase |
| TRX | thioredoxin |
| APX | ascorbate peroxidase |
| DAHR | dehydroascorbate reductase |
| GS | glutathione synthase |
References
- Celis-Plá, P.S.M.; Rodríguez-Rojas, F.; Méndez, L.; Moenne, F.; Muñoz, P.; Lobos, M.; Díaz, P.; Sánchez-Lizaso, J.L.; Brown, M.; Moenne, A.; et al. MAPK pathway under chronic copper excess in green macroalgae (Chlorophyta): Influence on metal exclusion/extrusion mechanisms and photosynthesis. Int. J. Mol. Sci. 2019, in press. [Google Scholar]
- Moenne, A.; González, A.; Sáez, C.A. Mechanisms of metal tolerance in marine macroalgae, with emphasis on copper tolerance in Chlorophyta and Rhodophyta. Aquat. Toxicol. 2016, 176, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, A.; González, A.; Gómez, M.; Contreras, R.A.; Díaz, P.; Lobos, G.; Brown, M.T.; Sáez, C.A.; Moenne, A. Copper excess detoxification is mediated by a coordinated and complementary induction of glutathione, phytochelatins and metallothioneins in the green seaweed Ulva compressa. Plant Physiol. Biochem. 2019, 135, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Sáez, C.A.; Roncarati, F.; Moenne, A.; Moody, J.A.; Brown, M.T. Copper-induced intra-specific oxidative damage and antioxidant responses in strains of the brown alga Ectocarpus siliculosus with different pollution histories. Aquat. Toxicol. 2015, 159, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Burkhead, J.L.; Reynolds, K.A.G.; Abdel-Ghany, S.E.; Cohu, C.M.; Pilon, M. Copper homeostasis. New Phytol. 2009, 182, 799–816. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Hendrix, S.; dos Reis, R.A.; De Smet, S.; Deckers, J.; Gielen, H.; Jozefczak, M.; Loix, C.; Vercampt, H.; Vangronsveld, J.; et al. Hydrogen peroxide, signaling in disguise during metal phytotoxicity. Front. Plant Sci. 2016, 7, 470. [Google Scholar] [CrossRef]
- Buchanan, B.B. The path to thioredoxin and redox regulation beyond chloroplasts. Plant Cell Physiol. 2017, 58, 1826–1832. [Google Scholar] [CrossRef]
- Ratkevicius, N.; Correa, J.A.; Moenne, A. Copper accumulation, synthesis of ascorbate and activation of ascorbate peroxidase in Enteromorpha compressa (L.) Grev. (Chlorophyta) from heavy metal-enriched environments in northern Chile. Plant Cell Environ. 2003, 26, 1599–1608. [Google Scholar] [CrossRef]
- González, A.; Vera, J.; Castro, J.; Dennett, G.; Mellado, M.; Morales, B.; Correa, J.A.; Moenne, A. Co-occurring increases of calcium and organellar reactive oxygen species determine differential activation of antioxidant and defense enzymes in Ulva compressa (Chlorophyta) exposed to copper excess. Plant Cell Environ. 2010, 33, 1627–1640. [Google Scholar] [CrossRef]
- González, A.; Cabrera Mde, L.; Henriquez, M.J.; Contreras, R.A.; Morales, B.; Moenne, A. Cross talk among calcium, hydrogen peroxide, and nitric oxide and activation of gene expression involving calmodulins and calcium-dependent protein kinases in Ulva compressa exposed to copper excess. Plant Physiol. 2012, 158, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Laporte, D.; Valdés, N.; González, A.; Sáez, C.A.; Zúñiga, A.; Navarrete, A.; Meneses, C.; Moenne, A. Copper-induced overexpression of genes encoding antioxidant system enzymes and metallothioneins involve the activation of CaMs, CDPKs and MEK1/2 in the marine alga Ulva compressa. Aquat. Toxicol. 2016, 177, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Mellado, M.; Contreras, R.A.; González, A.; Dennett, G.; Moenne, A. Copper-induced synthesis of ascorbate, glutathione and phytochelatins in the marine alga Ulva compressa (Chlorophyta). Plant Physiol. Biochem. 2012, 51, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Jervis, L.; Rees-Naesborg, R.; Brown, M. Biochemical responses of the marine macroalgae Ulva lactuca and Fucus vesiculosus to cadmium and copper-from sequestration to oxidative stress. Biochem. Soc. Trans. 1997, 25, 63S. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.M.; Lee, T.M. Regulation of activity and gene expression of antioxidant enzymes in Ulva fasciata Delile (Ulvales, Chlorophyta) in response to excess copper. Phycologia 2008, 47, 346–360. [Google Scholar] [CrossRef]
- Song, Y.; Cheong, Y.K.; Kim, N.H.; Chung, H.T.; Kang, D.G.; Pae, H.O. Mitogen-activated protein kinases and reactive oxygen species: How can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar]
- Sinha, A.K.; Jaggi, M.; Raghuram, B.; Tuteja, N. Mitogen-activated protein kinase signaling in plants under abiotic stress. Plant Signal. Behav. 2011, 6, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Janitza, P.; Ullrich, K.K.; Quint, M. Toward a comprehensive phylogenetic reconstruction of the evolutionary history of mitogen-activated protein kinases in the plant kingdom. Front. Plant Sci. 2012, 3, 271. [Google Scholar] [CrossRef]
- Parages, M.L.; Capasso, J.M.; Niell, F.X.; Jimenez, C. Responses of cyclic phosphorylation of MAPK-like proteins in intertidal macroalgae after environmental stress. J. Plant Physiol. 2014, 171, 276–284. [Google Scholar] [CrossRef]
- Charneco, G.O.; Parages, M.L.; Camarena-Gomez, T.; Jimenez, C. Phosphorylation of MAP Kinases crucially controls the response to environmental stress in Dunaliella viridis. Environ. Exp. Bot. 2018, 156, 203–213. [Google Scholar] [CrossRef]
- Parages, M.L.; Figueroa, F.L.; Conde-Alvarez, R.M.; Jimenez, C. Phosphorylation of MAPK-like proteins in three intertidal macroalgae under stress conditions. Aquat. Biol. 2014, 22, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Ismail, G.A.; Ismail, M.M. Variation in oxidative stress indices of two green seaweeds growing under different heavy metal stresses. Environ. Monit. Assess. 2017, 189, 12. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signalling: what’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Sáez, C.A.; González, A.; Contreras, R.A.; Moody, A.J.; Moenne, A.; Brown, M.T. A novel field transplantation technique reveals intra-specific metal-induced oxidative responses in strains of Ectocarpus siliculosus with different pollution histories. Environ. Pollut. 2015, 199, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Karuppanapandian, T.; Kim, W. Cobalt-induced oxidative stress causes growth inhibition associated with enhanced lipid peroxidation and activates antioxidant responses in Indian mustard (Brassica juncea L.) leaves. Acta Physiol. Plant. 2013, 35, 2429–2443. [Google Scholar] [CrossRef]
- Shan, C.; Sun, H. Jasmonic acid-induced NO activates MEK1/2 in regulating the metabolism of ascorbate and glutathione in maize leaves. Protoplasma 2018, 255, 977–983. [Google Scholar] [CrossRef]
- Qi, Q.; Guo, Z.; Liang, Y.; Li, K.; Xu, H. Hydrogen sulfide alleviates oxidative damage under excess nitrate stress through MAPK/NO signaling in cucumber. Plant Physiol. Biochem. 2019, 135, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Ashraf, M.; Bajguz, A.; Ahmad, P. Brassinosteroids regulate growth in plants under stressful environments and crosstalk with other potential phytohormones. J. Plant Growth Regul. 2018, 37, 1007–1024. [Google Scholar] [CrossRef]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef]
- Yoo, S.J.; Kim, S.H.; Kim, M.J.; Ryu, C.M.; Kim, Y.C.; Cho, B.H.; Yang, K.Y. Involvement of the OsMKK4-OsMPK1 CASCADE AND ITS DOWNSTREAM TRANSCRIPTION FACTOR OsWRKY53 in the wounding response in rice. Plant Pathol. J. 2014, 30, 168–177. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Wang, K.; Zhang, S.Y.; Ren, J.; Liu, T.; Wang, X. Crosstalk between ABA, auxin, MAPK signaling, and the cell cycle in cadmium-stressed rice seedlings. Acta Physiol. Plant. 2014, 36, 1879–1892. [Google Scholar] [CrossRef]
- Nazarian, R.; Shi, H.; Wang, Q.; Kong, X.; Koya, R.C.; Lee, H.; Chen, Z.; Lee, M.K.; Attar, N.; Sazegar, H.; et al. Melanomas acquire resistance to B-RAF(V600E) inhibition by RTK or N-RAS upregulation. Nature 2010, 468, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, J.; Vultur, A.; Lee, J.T.; Somasundaram, R.; Fukunaga-Kalabis, M.; Cipolla, A.K.; Wubbenhorst, B.; Xu, X.; Gimotty, P.A.; Kee, D.; et al. Acquired resistance to BRAF inhibitors mediated by a RAF kinase switch in melanoma can be overcome by cotargeting MEK and IGF-1R/PI3K. Cancer Cell 2010, 18, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Paraiso, K.H.; Xiang, Y.; Rebecca, V.W.; Abel, E.V.; Chen, Y.A.; Munko, A.C.; Wood, E.; Fedorenko, I.V.; Sondak, V.K.; Anderson, A.R.; et al. PTEN loss confers BRAF inhibitor resistance to melanoma cells through the suppression of BIM expression. Cancer Res. 2011, 71, 2750–2760. [Google Scholar] [CrossRef] [PubMed]
- Gopal, Y.N.; Deng, W.; Woodman, S.E.; Komurov, K.; Ram, P.; Smith, P.D.; Davies, M.A. Basal and treatment-induced activation of AKT mediates resistance to cell death by AZD6244 (ARRY-142886) in Braf-mutant human cutaneous melanoma cells. Cancer Res. 2010, 70, 8736–8747. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.A.; Kopetz, S. Overcoming resistance to MAPK pathway inhibitors. J. Natl. Cancer Inst. 2013, 105, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.V.; Basile, K.J.; Kugel, C.H., 3rd; Witkiewicz, A.K.; Le, K.; Amaravadi, R.K.; Karakousis, G.C.; Xu, X.; Xu, W.; Schuchter, L.M.; et al. Melanoma adapts to RAF/MEK inhibitors through FOXD3-mediated upregulation of ERBB3. J. Clin. Investig. 2013, 123, 2155–2168. [Google Scholar] [CrossRef] [PubMed]
- Newlaczyl, A.U.; Hood, F.E.; Coulson, J.M.; Prior, I.A. Decoding RAS isoform and codon-specific signalling. Biochem. Soc. Trans. 2014, 42, 742–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáez, C.A.; Ramesh, K.; Greco, M.; Bitonti, M.B.; Brown, M.T. Enzymatic antioxidant defences are transcriptionally regulated in Es524, a copper-tolerant strain of Ectocarpus siliculosus (Ectocarpales; Phaeophyceae). Phycologia 2015, 54, 425–429. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized assay for hydrogen peroxide determination in plant tissue using potassium iodide. Am. J. Anal. Chem. 2014, 5, 730. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 15–27. [Google Scholar]
- Rodríguez, F.E.; Laporte, D.; González, A.; Méndez, K.N.; Castro-Nallar, E.; Meneses, C.; Huidobro-Toro, J.P.; Moenne, A. Copper-induced increased expression of genes involved in photosynthesis, carotenoid synthesis and C assimilation in the marine alga Ulva compressa. BMC Genom. 2018, 19, 829. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Gorley, R.N.; Clarke, K. PERMANOVA+ for Primer: Guide to Software and Statistical Methods; PRIMER-e: Plymouth, UK, 2008; p. 218. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Encoded Product | Primer ID | Sequence 5′-3′ | Start Position | Amplicon Size (bp) |
|---|---|---|---|---|---|
| CAT | Catalase | F1_CAT_Ulva | AGCGGAACAAGTCGGCGGCAA | 717 | 208 |
| R1_CAT_Ulva | CAGCACCATGCGGCCAACGG | 905 | |||
| SOD | Superoxide dismutase | F1_SOD2_Ulva | CCCTGCACCGCCGTCTGTCG | 22 | 255 |
| R1_SOD2_Ulva | CGCGCTGCTGATCTTCGGAGCA | 255 | |||
| TRX | Thioredoxin | F1_Trx2_Ulva | GCTGAGTGGCTCCTCTTCGCTGCAT | 3 | 224 |
| R1_Trx2_Ulva | CGGCTGTGCTCACTGGTGCGT | 206 | |||
| APX | Ascorbate peroxidase | F1c_Apx1_Ulva | AGGACGCGTGGCCCAAGTGC | 131 | 297 |
| R1c_Apx1_Ulva | GTCTCCGCCGGATGGCCAAGG | 408 | |||
| DHAR | Dehydroascorbate reductase | F1_Dhar_Ulva | CGCGGACTCCGGCGACATCT | 309 | 327 |
| R1_Dhar_Ulva | AGCGCTGCGAGCTCCTCTGG | 616 | |||
| GS | Glutathione synthetase | F1_GS_Ulva | TGGCGGCGAAGCTGCAGGAA | 1322 | 245 |
| R1_GS_Ulva | GCCTGCCGCAACACCTCCCT | 1547 | |||
| 18S | 18S rRNA subunit | 18S-F-1183 | AAT TTG ACT CAA CAC GGG | 1183 | 448 |
| 18S-R-1631 | TAC AAA GGG CAG GGA CG | 1631 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Rojas, F.; Celis-Plá, P.S.M.; Méndez, L.; Moenne, F.; Muñoz, P.T.; Lobos, M.G.; Díaz, P.; Sánchez-Lizaso, J.L.; Brown, M.T.; Moenne, A.; et al. MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Involvement in the Regulation of Detoxification Mechanisms. Int. J. Mol. Sci. 2019, 20, 4546. https://doi.org/10.3390/ijms20184546
Rodríguez-Rojas F, Celis-Plá PSM, Méndez L, Moenne F, Muñoz PT, Lobos MG, Díaz P, Sánchez-Lizaso JL, Brown MT, Moenne A, et al. MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Involvement in the Regulation of Detoxification Mechanisms. International Journal of Molecular Sciences. 2019; 20(18):4546. https://doi.org/10.3390/ijms20184546
Chicago/Turabian StyleRodríguez-Rojas, Fernanda, Paula S. M. Celis-Plá, Lorena Méndez, Fabiola Moenne, Pamela T. Muñoz, M. Gabriela Lobos, Patricia Díaz, José Luis Sánchez-Lizaso, Murray T. Brown, Alejandra Moenne, and et al. 2019. "MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Involvement in the Regulation of Detoxification Mechanisms" International Journal of Molecular Sciences 20, no. 18: 4546. https://doi.org/10.3390/ijms20184546