Dysregulation of Dopaminergic Regulatory Factors TH, Nurr1, and Pitx3 in the Ventral Tegmental Area Associated with Neuronal Injury Induced by Chronic Morphine Dependence


Abstract
:1. Introduction
2. Results
2.1. Thionine Staining Showed Pathological Changes in VTA Dopaminergic Neurons
2.2. TH Expression in the VTA
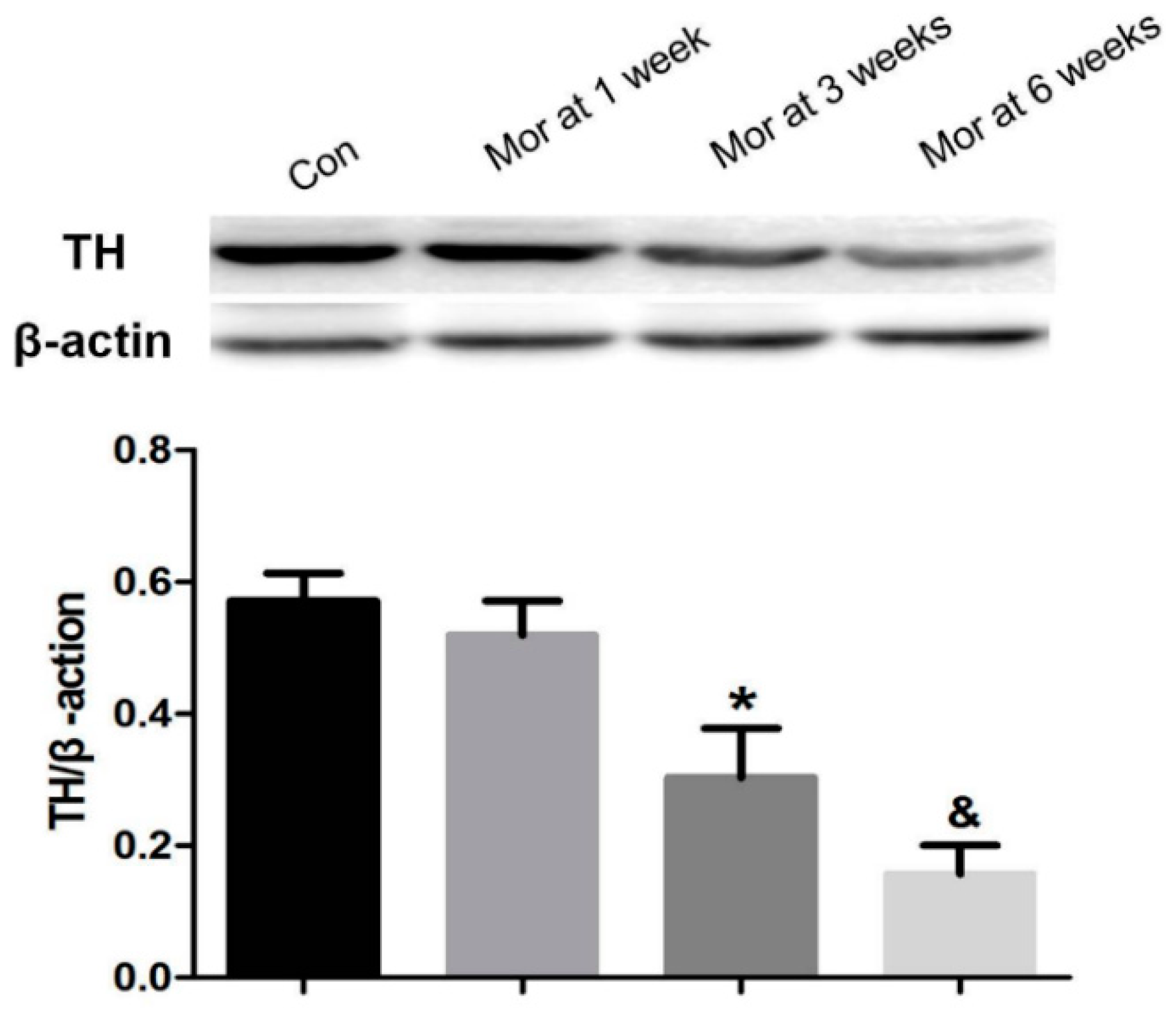
2.3. Western Blot Analysis of TH Expression
2.4. Nurr1 Expression in the VTA
2.5. Western Blot Analysis of Nurr1 Expression
2.6. Pitx3 Expression in the VTA
2.7. Western Blot Analysis of Pitx3 Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals
4.3. Models of Morphine Dependence
4.4. Tissue Preparation
4.5. Immunohistochemistry
4.6. Immunofluorescence Double Staining
4.7. Cell Counting
4.8. Western Blot Analysis
4.9. Statistical Methods
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Compton, W.M.; Volkow, N.D. Major increases in opioid analgesic abuse in the United States: Concerns and strategies. Drug Alcohol Depend. 2006, 81, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Skolnick, P. New medications for substance use disorders: Challenges and opportunities. Neuropsychopharmacology 2012, 37, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Masserano, J.M.; Baker, I.; Natsukari, N.; Wyatt, R.J. Chronic cocaine administration increases tyrosine hydroxylase activity in the ventral tegmental area through glutaminergic- and dopaminergic D2-receptor mechanisms. Neurosci. Lett. 1996, 217, 73–76. [Google Scholar] [CrossRef]
- Schmidt, E.F.; Sutton, M.A.; Schad, C.A.; Karanian, D.A.; Brodkin, E.S.; Self, D.W. Extinction training regulates tyrosine hydroxylase during withdrawal from cocaine self-administration. J. Neurosci. 2001, 21, Rc137. [Google Scholar] [CrossRef] [PubMed]
- Kalivas, P.W.; Volkow, N.D. The neural basis of addiction: A pathology of motivation and choice. Am. J. Psychiatry 2005, 162, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.Y.; Hong, S.; Jeong, J.W.; Choi, S.; Kim, H.; Kim, J.; Kim, K.S. Vesicular monoamine transporter 2 and dopamine transporter are molecular targets of Pitx3 in the ventral midbrain dopamine neurons. J. Neurochem. 2009, 111, 1202–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltran-Campos, V.; Silva-Vera, M.; Garcia-Campos, M.; Diaz-Cintra, S. Effects of morphine on brain plasticity. Neurologia 2015, 30, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Graziane, N.M.; Polter, A.M.; Briand, L.A.; Pierce, R.C.; Kauer, J.A. Kappa opioid receptors regulate stress-induced cocaine seeking and synaptic plasticity. Neuron 2013, 77, 942–954. [Google Scholar] [CrossRef]
- Jankovic, J.; Chen, S.; Le, W.D. The role of Nurr1 in the development of dopaminergic neurons and Parkinson’s disease. Prog. Neurobiol. 2005, 77, 128–138. [Google Scholar] [CrossRef]
- Smits, S.M.; Smidt, M.P. The role of Pitx3 in survival of midbrain dopaminergic neurons. J. Neural Transm. Suppl. 2006, 70, 57–60. [Google Scholar]
- Kadkhodaei, B.; Ito, T.; Joodmardi, E.; Mattsson, B.; Rouillard, C.; Carta, M.; Muramatsu, S.; Sumi-Ichinose, C.; Nomura, T.; Metzger, D.; et al. Nurr1 is required for maintenance of maturing and adult midbrain dopamine neurons. J. Neurosci. 2009, 29, 15923–15932. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.D.; Rayala, S.K.; Ohshiro, K.; Pakala, S.B.; Kobori, N.; Dash, P.; Yun, S.; Qin, J.; O’Malley, B.W.; Kumar, R. Multiple coregulatory control of tyrosine hydroxylase gene transcription. Proc. Natl. Acad. Sci. USA 2011, 108, 4200–4205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saucedo-Cardenas, O.; Quintana-Hau, J.D.; Le, W.D.; Smidt, M.P.; Cox, J.J.; De Mayo, F.; Burbach, J.P.; Conneely, O.M. Nurr1 is essential for the induction of the dopaminergic phenotype and the survival of ventral mesencephalic late dopaminergic precursor neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 4013–4018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, W.D.; Xu, P.; Jankovic, J.; Jiang, H.; Appel, S.H.; Smith, R.G.; Vassilatis, D.K. Mutations in NR4A2 associated with familial Parkinson disease. Nat. Genet. 2003, 33, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ohashi, R.; Naito, M. Expression of the nerve growth factor-induced gene B-beta in the developing rat brain and retina. Arch. Histol. Cytol. 2009, 72, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cong, B.; Ma, C.; Qi, Q.; Fu, L.; Zhang, G.; Min, Z. Expression of Nurr1 during rat brain and spinal cord development. Neurosci. Lett. 2011, 488, 49–54. [Google Scholar] [CrossRef]
- Li, Y.; Qi, Q.; Cong, B.; Shi, W.; Liu, X.; Zhang, G.; Ma, C. Expression patterns of Nurr1 in rat retina development. J. Mol. Histol. 2012, 43, 633–639. [Google Scholar] [CrossRef]
- Horvath, M.C.; Kovacs, G.G.; Kovari, V.; Majtenyi, K.; Hurd, Y.L.; Keller, E. Heroin abuse is characterized by discrete mesolimbic dopamine and opioid abnormalities and exaggerated nuclear receptor-related 1 transcriptional decline with age. J. Neurosci. 2007, 27, 13371–13375. [Google Scholar] [CrossRef]
- Garcia-Perez, D.; Lopez-Bellido, R.; Rodriguez, R.E.; Laorden, M.L.; Nunez, C.; Milanes, M.V. Dysregulation of dopaminergic regulatory mechanisms in the mesolimbic pathway induced by morphine and morphine withdrawal. Brain Struct. Funct. 2015, 220, 1901–1919. [Google Scholar] [CrossRef]
- Dejean, C.; Boraud, T.; Le Moine, C. Opiate dependence induces network state shifts in the limbic system. Neurobiol. Dis. 2013, 59, 220–229. [Google Scholar] [CrossRef]
- Hu, L.; Jing, X.H.; Cui, C.L.; Xing, G.G.; Zhu, B. NMDA receptors in the midbrain play a critical role in dopamine-mediated hippocampal synaptic potentiation caused by morphine. Addict. Biol. 2014, 19, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Mazei-Robison, M.S.; Koo, J.W.; Friedman, A.K.; Lansink, C.S.; Robison, A.J.; Vinish, M.; Krishnan, V.; Kim, S.; Siuta, M.A.; Galli, A.; et al. Role for mTOR signaling and neuronal activity in morphine-induced adaptations in ventral tegmental area dopamine neurons. Neuron 2011, 72, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Ma, C.; Qi, Q.; Liu, L.; Bi, H.; Cong, B.; Li, Y. The effect of different durations of morphine exposure on mesencephalic dopaminergic neurons in morphine dependent rats. Neurotoxicology 2015, 51, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Mazei-Robison, M.S.; Nestler, E.J. Opiate-induced molecular and cellular plasticity of ventral tegmental area and locus coeruleus catecholamine neurons. Cold Spring Harbor Perspect. Med. 2012, 2, a012070. [Google Scholar] [CrossRef]
- Nestler, E.J. Transcriptional mechanisms of drug addiction. Clin. Psychopharmacol. Neurosci. 2012, 10, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, F.M.; van der Linden, A.J.; Wang, Y.; von Oerthel, L.; Sul, H.S.; Burbach, J.P.; Smidt, M.P. Identification of Dlk1, Ptpru and Klhl1 as novel Nurr1 target genes in meso-diencephalic dopamine neurons. Development 2009, 136, 2363–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, F.M.; van Erp, S.; van der Linden, A.J.; von Oerthel, L.; Burbach, J.P.; Smidt, M.P. Pitx3 potentiates Nurr1 in dopamine neuron terminal differentiation through release of SMRT-mediated repression. Development 2009, 136, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Volpicelli, F.; De Gregorio, R.; Pulcrano, S.; Perrone-Capano, C.; di Porzio, U.; Bellenchi, G.C. Direct regulation of Pitx3 expression by Nurr1 in culture and in developing mouse midbrain. PLoS ONE 2012, 7, e30661. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Inoue, K.; Ishii, J.; Vanti, W.B.; Voronov, S.V.; Murchison, E.; Hannon, G.; Abeliovich, A. A MicroRNA feedback circuit in midbrain dopamine neurons. Science 2007, 317, 1220–1224. [Google Scholar] [CrossRef] [PubMed]
- Meye, F.J.; van Zessen, R.; Smidt, M.P.; Adan, R.A.; Ramakers, G.M. Morphine withdrawal enhances constitutive mu-opioid receptor activity in the ventral tegmental area. J. Neurosci. 2012, 32, 16120–16128. [Google Scholar] [CrossRef] [PubMed]
- Radke, A.K.; Rothwell, P.E.; Gewirtz, J.C. An anatomical basis for opponent process mechanisms of opiate withdrawal. J. Neurosci. 2011, 31, 7533–7539. [Google Scholar] [CrossRef] [PubMed]
- Allouche, S.; Noble, F.; Marie, N. Opioid receptor desensitization: Mechanisms and its link to tolerance. Front. Pharmacol. 2014, 5, 280. [Google Scholar] [CrossRef] [PubMed]
- Volakakis, N.; Kadkhodaei, B.; Joodmardi, E.; Wallis, K.; Panman, L.; Silvaggi, J.; Spiegelman, B.M.; Perlmann, T. NR4A orphan nuclear receptors as mediators of CREB-dependent neuroprotection. Proc. Natl. Acad. Sci. USA 2010, 107, 12317–12322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malewicz, M.; Kadkhodaei, B.; Kee, N.; Volakakis, N.; Hellman, U.; Viktorsson, K.; Leung, C.Y.; Chen, B.; Lewensohn, R.; van Gent, D.C.; et al. Essential role for DNA-PK-mediated phosphorylation of NR4A nuclear orphan receptors in DNA double-strand break repair. Genes Dev. 2011, 25, 2031–2040. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.; Aron, L.; Klein, R.; Li, M.; Wurst, W.; Prakash, N.; Le, W. Pitx3 is a critical mediator of GDNF-induced BDNF expression in nigrostriatal dopaminergic neurons. J. Neurosci. 2011, 31, 12802–12815. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, R.; Negus, S.; Koob, G.F. Precipitation of morphine withdrawal syndrome in rats by administration of mu-, delta- and kappa-selective opioid antagonists. Neuropharmacology 1992, 31, 1231–1241. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: Amsterdam, The Netherlands, 2007. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symptom | Morphine-Dependent Group | Control Group |
|---|---|---|
| wet dog shakes | 8.35 ± 1.86 * | 0.74 ± 0.61 |
| stretching | 15.52 ± 4.23 ** | 0.63 ± 0.50 |
| cleaning fur | 4.96 ± 1.59 * | 0.76 ± 0.53 |
| swallowing | 11.75 ± 1.78 ** | 0.89 ± 0.83 |
| standing | 8.00 ± 0.86 ** | 0.88 ± 0.63 |
| jumping | 3.45 ± 1.28 ** | 0.60 ± 0.76 |
| teeth chattering | 7.30 ± 1.78 ** | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.; Zhang, Y.; Zhao, G.; Wang, S.; Zhang, G.; Ma, C.; Li, Y.; Cong, B. Dysregulation of Dopaminergic Regulatory Factors TH, Nurr1, and Pitx3 in the Ventral Tegmental Area Associated with Neuronal Injury Induced by Chronic Morphine Dependence. Int. J. Mol. Sci. 2019, 20, 250. https://doi.org/10.3390/ijms20020250
Shi W, Zhang Y, Zhao G, Wang S, Zhang G, Ma C, Li Y, Cong B. Dysregulation of Dopaminergic Regulatory Factors TH, Nurr1, and Pitx3 in the Ventral Tegmental Area Associated with Neuronal Injury Induced by Chronic Morphine Dependence. International Journal of Molecular Sciences. 2019; 20(2):250. https://doi.org/10.3390/ijms20020250
Chicago/Turabian StyleShi, Weibo, Yaxing Zhang, Guoting Zhao, Songjun Wang, Guozhong Zhang, Chunling Ma, Yingmin Li, and Bin Cong. 2019. "Dysregulation of Dopaminergic Regulatory Factors TH, Nurr1, and Pitx3 in the Ventral Tegmental Area Associated with Neuronal Injury Induced by Chronic Morphine Dependence" International Journal of Molecular Sciences 20, no. 2: 250. https://doi.org/10.3390/ijms20020250