Cellular Responses of Industrially Relevant Silica Dust on Human Glial Cells In Vitro

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
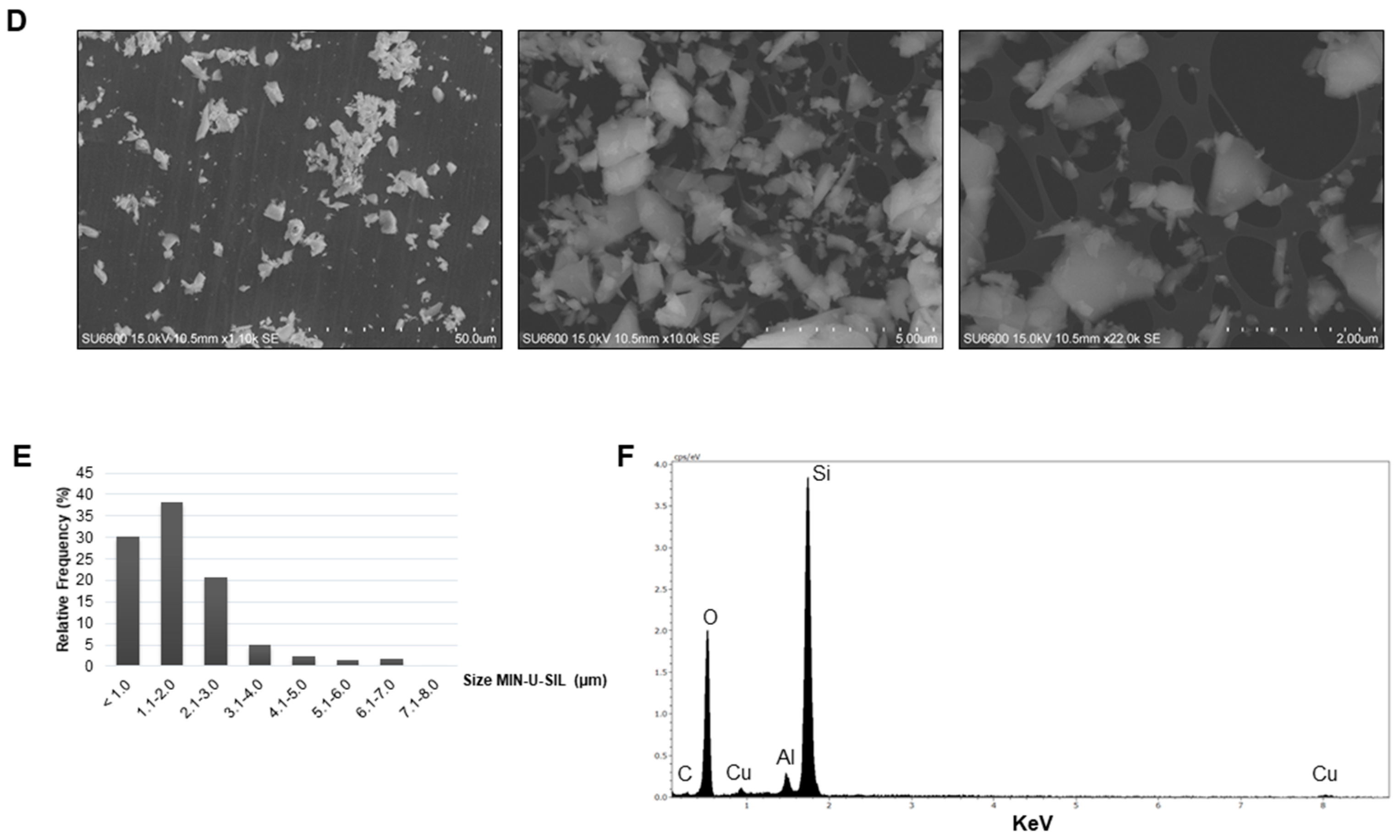
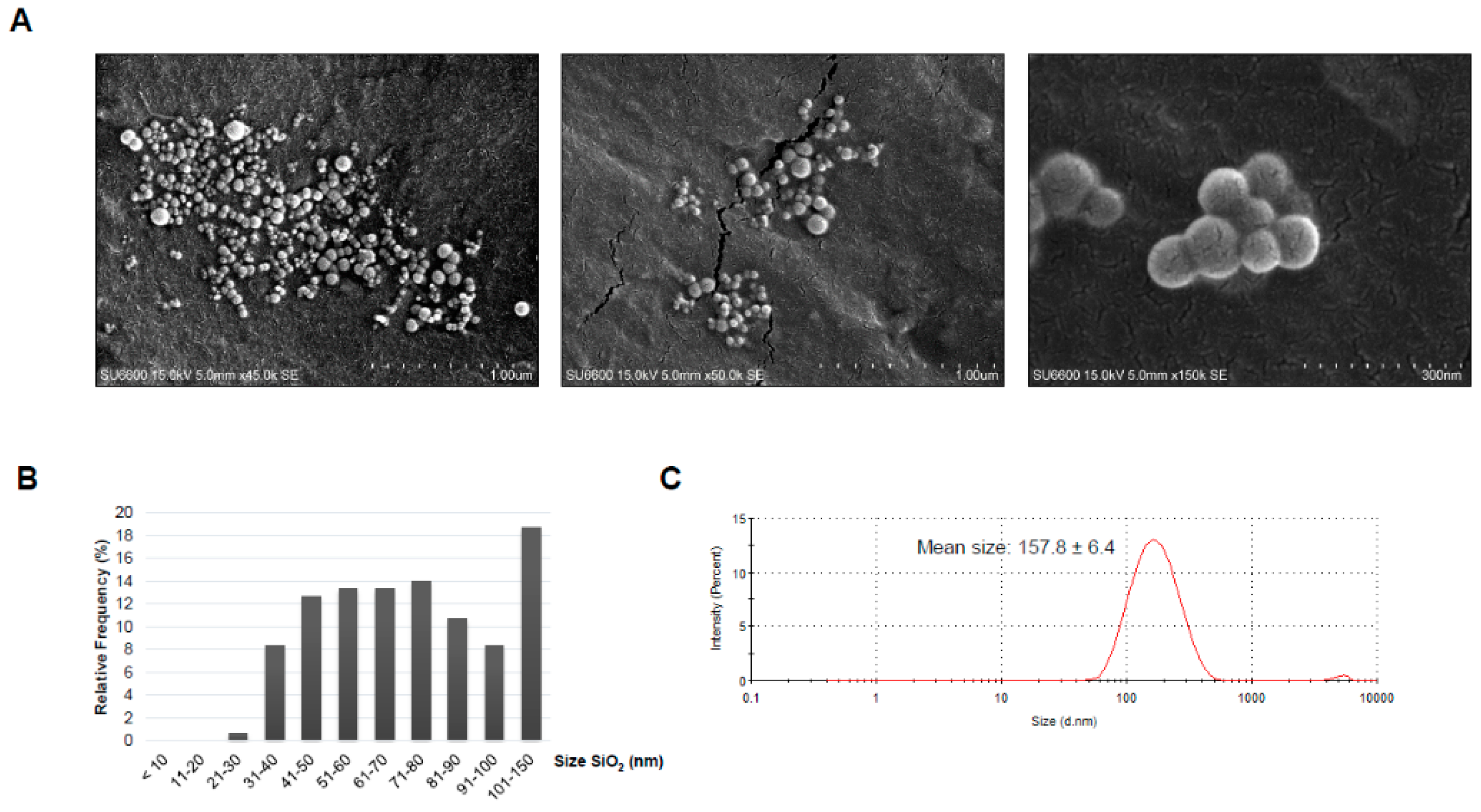
2.1. Characteristics of the Dust
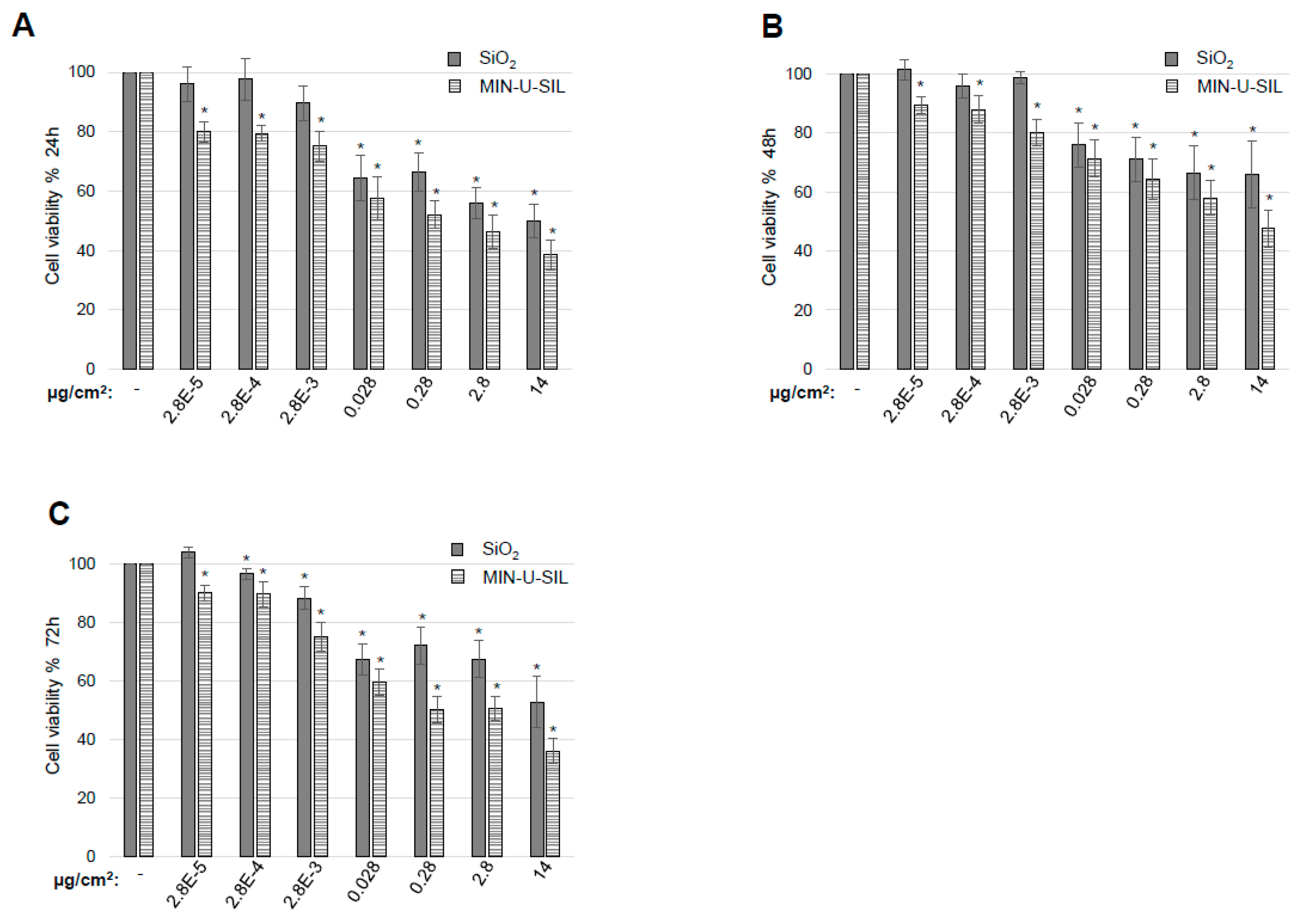
2.2. The Effect of the Two Types of SiO2 Dust on Cellular Endpoints
3. Discussion
4. Materials and Methods
4.1. Generation of the Dust
4.2. Preparation of the Dust for Characterization and Cell Culture Experiments
4.3. Dust Characterization
4.3.1. Scanning Electron Microscopy (SEM)
4.3.2. Dynamic Light Scattering (DLS)
4.4. Cells and Cell Culture
4.5. Estimation of Dust Doses Used for the Experiments
4.6. Cytotoxicity Assay
4.7. Quantitative PCR (qPCR) for Measurement of Gene Expression
4.8. Detection of APP by Western Blotting Analysis
4.9. Functional Assay of Gap Junctional Intercellular Communication (GJIC) by Scrape Loading
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kero, I.; Grådahl, S.; Tranell, G. Airborne Emissions from Si/FeSi Production. JOM 2017, 69, 365–380. [Google Scholar] [CrossRef]
- IARC. Silica. Iarc Monogr. Eval. Carcinog. Risks Hum. 1997, 68, 41–242. [Google Scholar]
- Cassidy, A.; Mannetje, A.; van Tongeren, M.; Field, J.K.; Zaridze, D.; Szeszenia-Dabrowska, N.; Rudnai, P.; Lissowska, J.; Fabianova, E.; Mates, D.; et al. Occupational exposure to crystalline silica and risk of lung cancer: A multicenter case-control study in Europe. Epidemiology 2007, 18, 36–43. [Google Scholar] [CrossRef]
- Cox, L.A., Jr. An exposure-response threshold for lung diseases and lung cancer caused by crystalline silica. Risk Anal. Off. Publ. Soc. Risk Anal. 2011, 31, 1543–1560. [Google Scholar]
- Finkelstein, M.M. Silica, silicosis, and lung cancer: A risk assessment. Am. J. Ind. Med. 2000, 38, 8–18. [Google Scholar] [CrossRef]
- Rappaport, S.M.; Goldberg, M.; Susi, P.; Herrick, R.F. Excessive exposure to silica in the US construction industry. Ann. Occup. Hyg. 2003, 47, 111–122. [Google Scholar]
- Perkins, T.N.; Shukla, A.; Peeters, P.M.; Steinbacher, J.L.; Landry, C.C.; Lathrop, S.A.; Steele, C.; Reynaert, N.L.; Wouters, E.F.; Mossman, B.T. Differences in gene expression and cytokine production by crystalline vs. amorphous silica in human lung epithelial cells. Part. Fibre Toxicol. 2012, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Staronova, K.; Nielsen, J.B.; Roursgaard, M.; Knudsen, L.E. Transport of SiO(2) nanoparticles through human skin. Basic Clin. Pharmacol. Toxicol. 2012, 111, 142–144. [Google Scholar] [PubMed]
- Kreyling, W.G. Discovery of unique and ENM- specific pathophysiologic pathways: Comparison of the translocation of inhaled iridium nanoparticles from nasal epithelium versus alveolar epithelium towards the brain of rats. Toxicol. Appl. Pharmacol. 2016, 299, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Oberdorster, G.; Elder, A.; Rinderknecht, A. Nanoparticles and the brain: Cause for concern? J. Nanosci. Nanotechnol. 2009, 9, 4996–5007. [Google Scholar] [CrossRef]
- Capeletti, L.B.; Loiola, L.M.D.; Picco, A.S.; da Silva Liberato, M.; Cardoso, M.B. 8-Silica Nanoparticle Applications in the Biomedical Field. In Smart Nanoparticles for Biomedicine; Ciofani, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 115–129. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Ito, A.; Watanabe, Y.; Sogo, Y.; Tsuji, N.M.; Ohno, T. Stimulation of In Vivo Antitumor Immunity with Hollow Mesoporous Silica Nanospheres. Angew. Chem. (Int. Ed. Engl.) 2016, 55, 1899–1903. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Ito, A.; Yoshiyuki, K.; Sogo, Y.; Watanabe, Y.; Yamazaki, A.; Ohno, T.; Tsuji, N.M. Hollow Structure Improved Anti-Cancer Immunity of Mesoporous Silica Nanospheres In Vivo. Small 2016, 12, 3510–3515. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Yoshiyuki, K.; Watanabe, Y.; Sogo, Y.; Ohno, T.; Tsuji, N.M.; Ito, A. Cancer Immunotherapy: Comprehensive Mechanism Analysis of Mesoporous-Silica-Nanoparticle-Induced Cancer Immunotherapy (Adv. Healthcare Mater. 10/2016). Adv. Healthc. Mater. 2016, 5, 1246. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.Y.; Joachim, E.; Choi, H.; Kim, K. Toxicity of silica nanoparticles depends on size, dose, and cell type. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Nabeshi, H.; Yoshikawa, T.; Matsuyama, K.; Nakazato, Y.; Tochigi, S.; Kondoh, S.; Hirai, T.; Akase, T.; Nagano, K.; Abe, Y.; et al. Amorphous nanosilica induce endocytosis-dependent ROS generation and DNA damage in human keratinocytes. Part. Fibre Toxicol. 2011, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, C.; Sun, J.; Xue, Y. Neurotoxicity of silica nanoparticles: Brain localization and dopaminergic neurons damage pathways. ACS Nano 2011, 5, 4476–4489. [Google Scholar] [CrossRef] [PubMed]
- Kretowski, R.; Kusaczuk, M.; Naumowicz, M.; Kotynska, J.; Szynaka, B.; Cechowska-Pasko, M. The Effects of Silica Nanoparticles on Apoptosis and Autophagy of Glioblastoma Cell Lines. Nanomaterials 2017, 7, 230. [Google Scholar] [CrossRef]
- Liu, X.; Sui, B.; Sun, J. Blood-brain barrier dysfunction induced by silica NPs in vitro and in vivo: Involvement of oxidative stress and Rho-kinase/JNK signaling pathways. Biomaterials 2017, 121, 64–82. [Google Scholar] [CrossRef]
- Pekny, M.; Wilhelmsson, U.; Pekna, M. The dual role of astrocyte activation and reactive gliosis. Neurosci. Lett. 2014, 565, 30–38. [Google Scholar] [CrossRef]
- Næss, M.K.; Young, D.J.; Zhang, J.; Olsen, J.E.; Tranell, G. Active Oxidation of Liquid Silicon: Experimental Investigation of Kinetics. Oxid. Met. 2012, 78, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Frost, G.R.; Li, Y.M. The role of astrocytes in amyloid production and Alzheimer’s disease. Open Biol. 2017, 7. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Wong, P.C. Amyloid precursor protein processing and Alzheimer’s disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Smith, K. Industrial Applications of X-ray Diffraction; Taylor & Francis Inc.: Bosa Roca, FL, USA, 2000. [Google Scholar]
- Pandurangi, R.S.; Seehra, M.S.; Razzaboni, B.L.; Bolsaitis, P. Surface and bulk infrared modes of crystalline and amorphous silica particles: A study of the relation of surface structure to cytotoxicity of respirable silica. Environ. Health Perspect. 1990, 86, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Dingsøyr, E.; Dåstøl, M.; Wedberg, W.C. Particle size and particle size distribution of Elkem Microsilica and its relevance to technical application. In Proceedings of the 5. European Symposium Particle Characterization, Nurnberg, Germany, 24–26 March 1992. [Google Scholar]
- Kolderup, H. Particle Size Distribution of Fumes Formed by Ferrosilicon Production. J. Air Pollut. Control Assoc. 1977, 27, 127–130. [Google Scholar] [CrossRef] [Green Version]
- Kero, I.; Naess, M.K.; Tranell, G. Particle size distributions of particulate emissions from the ferroalloy industry evaluated by electrical low pressure impactor (ELPI). J. Occup. Environ. Hyg. 2015, 12, 37–44. [Google Scholar] [CrossRef]
- Kero, I.T.; Jorgensen, R.B. Comparison of Three Real-Time Measurement Methods for Airborne Ultrafine Particles in the Silicon Alloy Industry. Int. J. Environ. Res. Public Health 2016, 13, 871. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.L.; Gao, J.Q. Potential neurotoxicity of nanoparticles. Int. J. Pharm. 2010, 394, 115–121. [Google Scholar] [CrossRef]
- Murugadoss, S.; Lison, D.; Godderis, L.; Van Den Brule, S.; Mast, J.; Brassinne, F.; Sebaihi, N.; Hoet, P.H. Toxicology of silica nanoparticles: An update. Arch. Toxicol. 2017, 91, 2967–3010. [Google Scholar] [CrossRef]
- Zhou, M.; Xie, L.; Fang, C.-J.; Yang, H.; Wang, Y.-J.; Zhen, X.-Y.; Yan, C.-H.; Wang, Y.; Zhao, M.; Peng, S. Implications for blood-brain-barrier permeability, in vitro oxidative stress and neurotoxicity potential induced by mesoporous silica nanoparticles: Effects of surface modification. Rsc Adv. 2016, 6, 2800–2809. [Google Scholar] [CrossRef]
- Castranova, V.; Porter, D.; Millecchia, L.; Ma, J.Y.; Hubbs, A.F.; Teass, A. Effect of inhaled crystalline silica in a rat model: Time course of pulmonary reactions. Mol. Cell. Biochem. 2002, 234–235, 177–184. [Google Scholar] [CrossRef]
- Gilberti, R.M.; Joshi, G.N.; Knecht, D.A. The phagocytosis of crystalline silica particles by macrophages. Am. J. Respir. Cell Mol. Biol. 2008, 39, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Sellamuthu, R.; Umbright, C.; Li, S.; Kashon, M.; Joseph, P. Mechanisms of crystalline silica-induced pulmonary toxicity revealed by global gene expression profiling. Inhal. Toxicol. 2011, 23, 927–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batarseh, Y.S.; Duong, Q.V.; Mousa, Y.M.; Al Rihani, S.B.; Elfakhri, K.; Kaddoumi, A. Amyloid-beta and Astrocytes Interplay in Amyloid-beta Related Disorders. Int. J. Mol. Sci. 2016, 17, 338. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; He, C.; Li, J.; Chen, H.; Ma, Q.; Sui, X.; Tian, S.; Ying, M.; Zhang, Q.; Luo, Y.; et al. Uptake of silica nanoparticles: Neurotoxicity and Alzheimer-like pathology in human SK-N-SH and mouse neuro2a neuroblastoma cells. Toxicol. Lett. 2014, 229, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Almenar-Queralt, A.; Falzone, T.L.; Shen, Z.; Lillo, C.; Killian, R.L.; Arreola, A.S.; Niederst, E.D.; Ng, K.S.; Kim, S.N.; Briggs, S.P.; et al. UV irradiation accelerates amyloid precursor protein (APP) processing and disrupts APP axonal transport. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 3320–3339. [Google Scholar] [CrossRef]
- Le, H.T.; Sin, W.C.; Lozinsky, S.; Bechberger, J.; Vega, J.L.; Guo, X.Q.; Saez, J.C.; Naus, C.C. Gap junction intercellular communication mediated by connexin43 in astrocytes is essential for their resistance to oxidative stress. J. Biol. Chem. 2014, 289, 1345–1354. [Google Scholar] [CrossRef] [PubMed]
- Sin, W.C.; Aftab, Q.; Bechberger, J.F.; Leung, J.H.; Chen, H.; Naus, C.C. Astrocytes promote glioma invasion via the gap junction protein connexin43. Oncogene 2016, 35, 1504–1516. [Google Scholar] [CrossRef]
- Soroceanu, L.; Manning, T.J., Jr.; Sontheimer, H. Reduced expression of connexin-43 and functional gap junction coupling in human gliomas. Glia 2001, 33, 107–117. [Google Scholar] [CrossRef]
- Suadicani, S.O.; Flores, C.E.; Urban-Maldonado, M.; Beelitz, M.; Scemes, E. Gap junction channels coordinate the propagation of intercellular Ca2+ signals generated by P2Y receptor activation. Glia 2004, 48, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Arnoldussen, Y.J.; Ervik, T.K.; Berlinger, B.; Kero, I.; Shaposhnikov, S.; Zienolddiny, S. Cellular responses of human astrocytoma cells to dust from the Acheson process: An in vitro study. Neurotoxicology 2017. [Google Scholar] [CrossRef]
- Jensen, K.; Kembouche, Y.; Christiansen, E.; Jacobsen, N.; Wallin, H.; Guiot, C. The generic NANOGENOTOX dispersion protocol. Stand. Oper. Proced. Backgr. Doc. Final Protoc. Prod. Suitable Manuf. Nanomater. Expo. Media 2011, 1–31. [Google Scholar]
- Phuyal, S.; Kasem, M.; Rubio, L.; Karlsson, H.L.; Marcos, R.; Skaug, V.; Zienolddiny, S. Effects on human bronchial epithelial cells following low-dose chronic exposure to nanomaterials: A 6-month transformation study. Toxicol. Vitr. Int. J. Publ. Assoc. Bibra 2017, 44, 230–240. [Google Scholar] [CrossRef]
- Macintyre, E.H.; Wintersgill, C.J.; Thormar, H. The establishment of a line of visna virus-producing human astrocytes (V-1181N1). Med. Res. Eng. 1972, 11, 7–13. [Google Scholar]
- Antonini, J.M.; Roberts, J.R.; Chapman, R.S.; Soukup, J.M.; Ghio, A.J.; Sriram, K. Pulmonary toxicity and extrapulmonary tissue distribution of metals after repeated exposure to different welding fumes. Inhal. Toxicol. 2010, 22, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Antonini, J.M.; Roberts, J.R.; Schwegler-Berry, D.; Mercer, R.R. Comparative microscopic study of human and rat lungs after overexposure to welding fume. Ann. Occup. Hyg. 2013, 57, 1167–1179. [Google Scholar] [PubMed]
- Oberdorster, G.; Maynard, A.; Donaldson, K.; Castranova, V.; Fitzpatrick, J.; Ausman, K.; Carter, J.; Karn, B.; Kreyling, W.; Lai, D.; et al. Principles for characterizing the potential human health effects from exposure to nanomaterials: Elements of a screening strategy. Part. Fibre Toxicol. 2005, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Oberdorster, G.; Oberdorster, E.; Oberdorster, J. Nanotoxicology: An emerging discipline evolving from studies of ultrafine particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Stone, K.C.; Mercer, R.R.; Gehr, P.; Stockstill, B.; Crapo, J.D. Allometric relationships of cell numbers and size in the mammalian lung. Am. J. Respir. Cell Mol. Biol. 1992, 6, 235–243. [Google Scholar] [CrossRef]
- Arnoldussen, Y.J.; Skogstad, A.; Skaug, V.; Kasem, M.; Haugen, A.; Benker, N.; Weinbruch, S.; Apte, R.N.; Zienolddiny, S. Involvement of IL-1 genes in the cellular responses to carbon nanotube exposure. Cytokine 2015, 73, 128–137. [Google Scholar] [CrossRef]
- Opsahl, H.; Rivedal, E. Quantitative determination of gap junction intercellular communication by scrape loading and image analysis. Cell Adhes. Commun. 2000, 7, 367–375. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arnoldussen, Y.J.; Kringlen Ervik, T.; Baarnes Eriksen, M.; Kero, I.; Skaug, V.; Zienolddiny, S. Cellular Responses of Industrially Relevant Silica Dust on Human Glial Cells In Vitro. Int. J. Mol. Sci. 2019, 20, 358. https://doi.org/10.3390/ijms20020358
Arnoldussen YJ, Kringlen Ervik T, Baarnes Eriksen M, Kero I, Skaug V, Zienolddiny S. Cellular Responses of Industrially Relevant Silica Dust on Human Glial Cells In Vitro. International Journal of Molecular Sciences. 2019; 20(2):358. https://doi.org/10.3390/ijms20020358
Chicago/Turabian StyleArnoldussen, Yke Jildouw, Torunn Kringlen Ervik, Mina Baarnes Eriksen, Ida Kero, Vidar Skaug, and Shanbeh Zienolddiny. 2019. "Cellular Responses of Industrially Relevant Silica Dust on Human Glial Cells In Vitro" International Journal of Molecular Sciences 20, no. 2: 358. https://doi.org/10.3390/ijms20020358