A Mechanistic Perspective on PEX1 and PEX6, Two AAA+ Proteins of the Peroxisomal Protein Import Machinery

 and
and 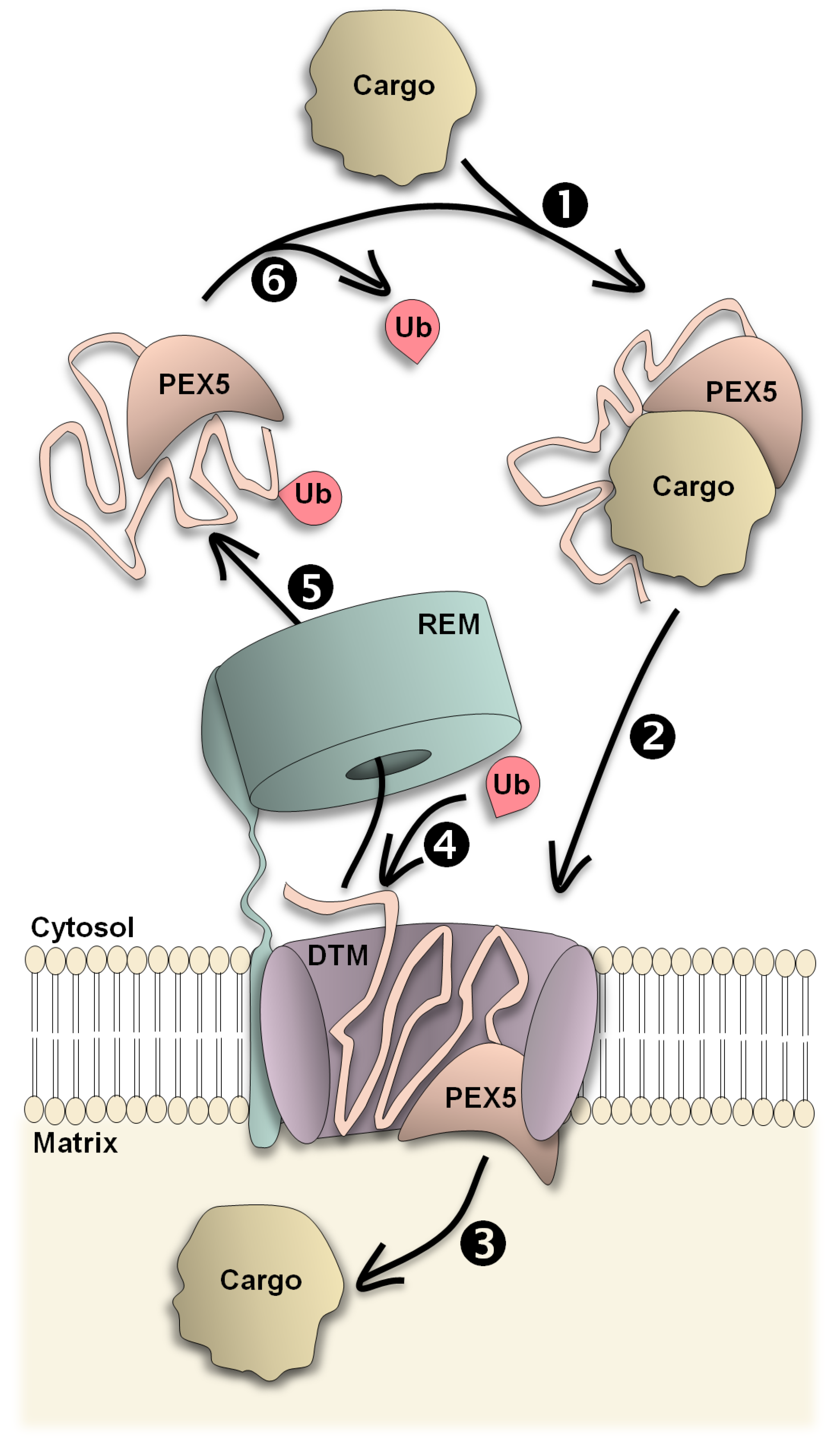{kind=link}
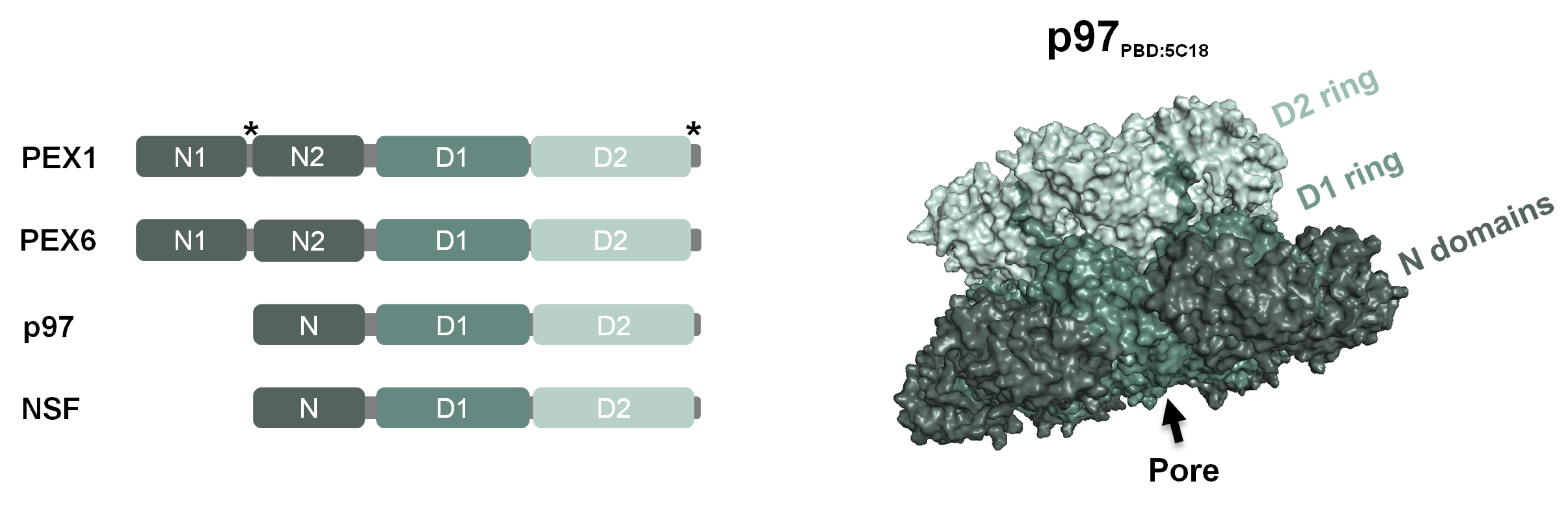{kind=link}
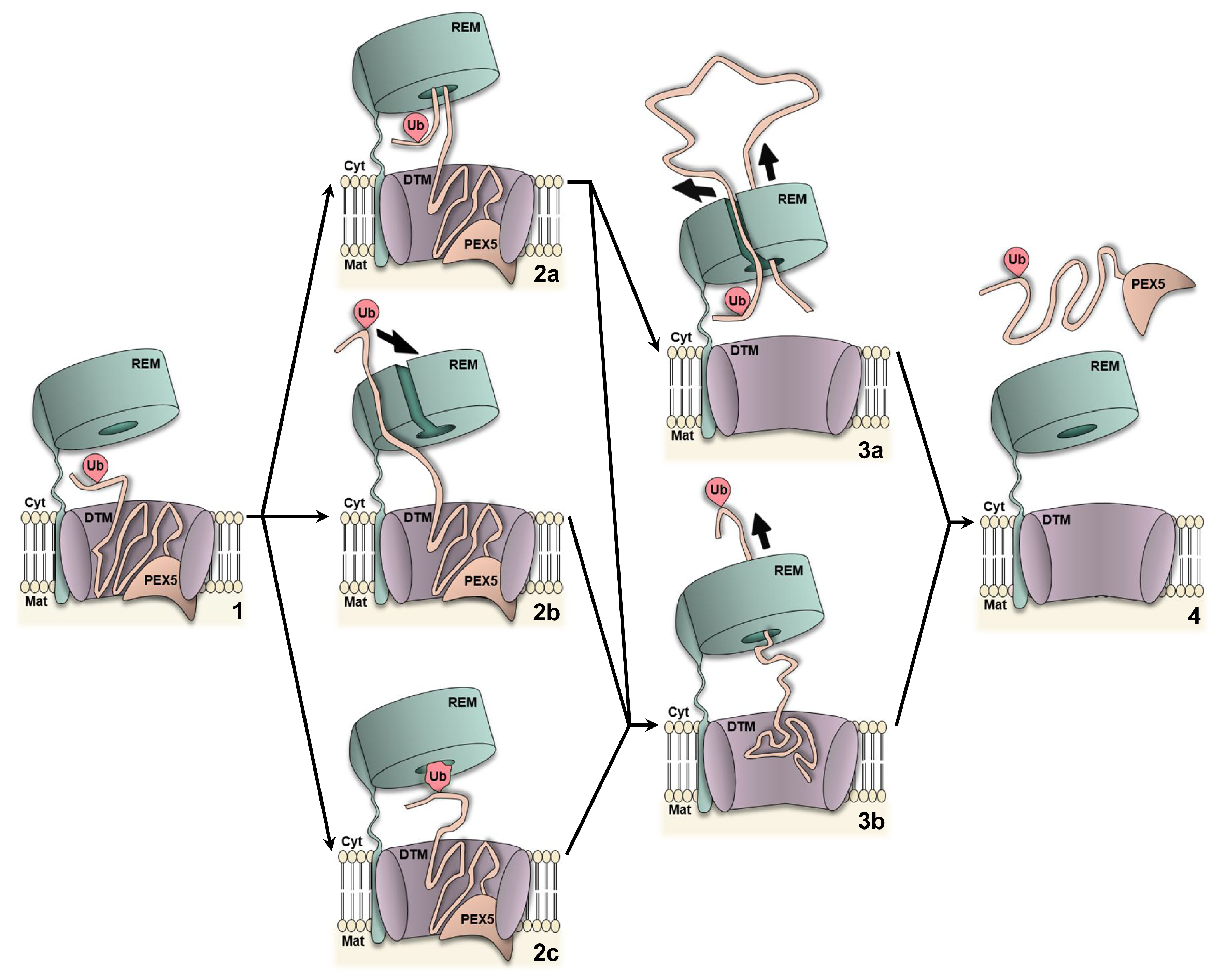{kind=link}
Abstract
1. Introduction
2. Going through the Peroxisomal Membrane—the Docking/Translocation Module and PEX5
3. The Receptor Export Module—the Initial Findings
4. The Mechanism of the Receptor Export Module
5. Unsolved Mechanistic Aspects: Substrate Engagement and Regulation
6. The Energetic Cost of Protein Translocation across the Peroxisomal Membrane
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAA+ | ATPases Associated with diverse cellular Activities |
| Cys11 | Cysteine 11 |
| Cyt | Cytosol |
| D1 | AAA+ domain 1 |
| D2 | AAA+ domain 2 |
| DHFR | Dihydrofolate reductase |
| DTM | Docking/translocation module |
| GFP | Green fluorescent protein |
| Kd | Dissociation constant |
| Mat | Matrix |
| N1 | N-terminal domain 1 |
| N2 | N-terminal domain 2 |
| NSF | N-ethylmaleimide sensitive factor |
| PTS | Peroxisomal targeting signal |
| REM | Receptor export module |
| RING | Really interesting new gene |
| SLiMs | Short linear motifs |
| SRH | Second region of homology |
| Tat | Twin-arginine translocation |
| Ub | Ubiquitin |
| Ub-PEX5 | Monoubiquitinated PEX5 |
References
- Becker, T.; Song, J.; Pfanner, N. Versatility of Preprotein Transfer from the Cytosol to Mitochondria. Trends Cell Biol. 2019, 29, 534–548. [Google Scholar] [CrossRef] [PubMed]
- Haßdenteufel, S.; Nguyen, D.; Helms, V.; Lang, S.; Zimmermann, R. ER import of small human presecretory proteins: components and mechanisms. FEBS Lett. 2019, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Francisco, T.; Rodrigues, T.A.; Dias, A.F.; Barros-Barbosa, A.; Bicho, D.; Azevedo, J.E. Protein transport into peroxisomes: Knowns and unknowns. BioEssays 2017, 39, 1700047. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.G.; Herrmann, J.M. Import of proteins into mitochondria. Protein J. 2019, 38, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Christie, P.J. The Rich Tapestry of Bacterial Protein Translocation Systems. Protein J. 2019, 38, 389–408. [Google Scholar] [CrossRef]
- Smets, D.; Loos, M.S.; Karamanou, S.; Economou, A. Protein Transport Across the Bacterial Plasma Membrane by the Sec Pathway. Protein J. 2019, 38, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Wickner, W.; Schekman, R. Protein Translocation Across Biological Membranes. Science 2005, 310, 1452–1456. [Google Scholar] [CrossRef]
- Corey, R.A.; Allen, W.J.; Collinson, I. Protein translocation: What’s the problem? Biochem. Soc. Trans. 2016, 44, 753–759. [Google Scholar] [CrossRef]
- Hepp, C.; Maier, B. Bacterial Translocation Ratchets: Shared Physical Principles with Different Molecular Implementations. BioEssays 2017, 39, 1700099. [Google Scholar] [CrossRef]
- Crow, A.; Greene, N.P.; Kaplan, E.; Koronakis, V. Structure and mechanotransmission mechanism of the MacB ABC transporter superfamily. Proc. Natl. Acad. Sci. USA 2017, 114, 12572–12577. [Google Scholar] [CrossRef]
- Pfanner, N.; Warscheid, B.; Wiedemann, N. Mitochondrial proteins: from biogenesis to functional networks. Nat. Rev. Mol. Cell Biol. 2019, 20, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Galán, J.E.; Waksman, G. Protein-Injection Machines in Bacteria. Cell 2018, 172, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.S.; Douzi, B.; Durand, E.; Roussel, A.; Cascales, E.; Cambillau, C. Towards a complete structural deciphering of Type VI secretion system. Curr. Opin. Struct. Biol. 2018, 49, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Dodt, G.; Gould, S.J. Multiple PEX genes are required for proper subcellular distribution and stability of Pex5p, the PTS1 receptor: evidence that PTS1 protein import is mediated by a cycling receptor. J. Cell Biol. 1996, 135, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Marzioch, M.; Erdmann, R.; Veenhuis, M.; Kunau, W.H. PAS7 encodes a novel yeast member of the WD-40 protein family essential for import of 3-oxoacyl-CoA thiolase, a PTS2-containing protein, into peroxisomes. EMBO J. 1994, 13, 4908–4918. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.E.; Gouveia, A.M.; Pinto, R.A.; Sá-Miranda, C.; Azevedo, J.E. The energetics of Pex5p-mediated peroxisomal protein import. J. Biol. Chem. 2003, 278, 39483–39488. [Google Scholar] [CrossRef] [PubMed]
- Alencastre, I.S.; Rodrigues, T.A.; Grou, C.P.; Fransen, M.; Sá-Miranda, C.; Azevedo, J.E. Mapping the cargo protein membrane translocation step into the PEX5 cycling pathway. J. Biol. Chem. 2009, 284, 27243–27251. [Google Scholar] [CrossRef]
- Francisco, T.; Rodrigues, T.A.; Freitas, M.O.; Grou, C.P.; Carvalho, A.F.; Sá-Miranda, C.; Pinto, M.P.; Azevedo, J.E. A cargo-centered perspective on the PEX5 receptor-mediated peroxisomal protein import pathway. J. Biol. Chem. 2013, 288, 29151–29159. [Google Scholar] [CrossRef]
- McCollum, D.; Monosov, E.; Subramani, S. The pas8 mutant of Pichia pastoris exhibits the peroxisomal protein import deficiencies of Zellweger syndrome cells--the PAS8 protein binds to the COOH-terminal tripeptide peroxisomal targeting signal, and is a member of the TPR protein family. J. Cell Biol. 1993, 121, 761–774. [Google Scholar] [CrossRef]
- Fransen, M.; Breesm, C.; Baumgart, E.; Vanhooren, J.C.T.; Baes, M.; Mannaerts, G.P.; Van Veldhoven, P.P. Identification and characterization of the putative human peroxisomal C-terminal targeting signal import receptor. J. Biol. Chem. 1995, 270, 7731–7736. [Google Scholar] [CrossRef]
- Wiemer, E.A.C.; Nuttley, W.M.; Bertolaet, B.L.; Li, X.; Francke, U.; Wheelock, M.J.; Anné, U.K.; Johnson, K.R.; Subramani, S. Human peroxisomal targeting signal-1 receptor restores peroxisomal protein import in cells from patients with fatal peroxisomal disorders. J. Cell Biol. 1995, 130, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Van Der Klei, I.J.; Hilbrands, R.E.; Swaving, G.J.; Waterham, H.R.; Vrieling, E.G.; Titorenko, V.I.; Cregg, J.M.; Harder, W.; Veenhuis, M. The Hansenula polymorpha PER3 gene is essential for the import of PTS1 proteins into the peroxisomal matrix. J. Biol. Chem. 1995, 270, 17229–17236. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, C.; Schmid, M.; Veenhuis, M.; Gietl, C. The plant PTS1 receptor: similarities and differences to its human and yeast counterparts. Plant J. 1998, 16, 453–464. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kragler, F.; Lametschwandtner, G.; Christmann, J.; Hartig, A.; Harada, J.J. Identification and analysis of the plant peroxisomal targeting signal 1 receptor NtPEX5. Proc. Natl. Acad. Sci. USA 1998, 95, 13336–13341. [Google Scholar] [CrossRef]
- Jardim, A.; Liu, W.; Zheleznova, E.; Ullman, B. Peroxisomal targeting signal-1 receptor protein PEX5 from Leishmania donovani. Molecular, biochemical, and immunocytochemical characterization. J. Biol. Chem. 2000, 275, 13637–13644. [Google Scholar] [CrossRef][Green Version]
- Costa-Rodrigues, J.; Carvalho, A.F.; Fransen, M.; Hambruch, E.; Schliebs, W.; Sá-Miranda, C.; Azevedo, J.E. Pex5p, the peroxisomal cycling receptor, is a monomeric non-globular protein. J. Biol. Chem. 2005, 280, 24404–24411. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Costa-Rodrigues, J.; Correia, I.; Faria, T.Q.; Martins, C.L.; Fransen, M.; Sá-Miranda, C.; Azevedo, J.E. The N-terminal half of the peroxisomal cycling receptor Pex5p is a natively unfolded domain. J. Mol. Biol. 2006, 356, 864–875. [Google Scholar] [CrossRef]
- Terlecky, S.R.; Nuttley, W.M.; McCollum, D.; Sock, E.; Subramani, S. The Pichia pastoris peroxisomal protein PAS8p is the receptor for the C-terminal tripeptide peroxisomal targeting signal. EMBO J. 1995, 14, 3627–3634. [Google Scholar] [CrossRef]
- Brocard, C.; Hartig, A. Peroxisome targeting signal 1: is it really a simple tripeptide? Biochim. Biophys. Acta 2006, 1763, 1565–1573. [Google Scholar] [CrossRef]
- Lazarow, P.B. The import receptor Pex7p and the PTS2 targeting sequence. Biochim. Biophys. Acta 2006, 1763, 1599–1604. [Google Scholar] [CrossRef]
- Braverman, N.E.; Dodt, G.; Gould, S.J.; Valle, D. An isoform of pex5p, the human PTS1 receptor, is required for the import of PTS2 proteins into peroxisomes. Hum. Mol. Genet. 1998, 7, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Otera, H.; Okumoto, K.; Tateishi, K.; Ikoma, Y.; Matsuda, E.; Nishimura, M.; Tsukamoto, T.; Osumi, T.; Ohashi, K.; Higuchi, O.; et al. Peroxisome targeting signal type 1 (PTS1) receptor is involved in import of both PTS1 and PTS2: studies with PEX5-defective CHO cell mutants. Mol. Cell. Biol. 1998, 18, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, T.; Otera, H.; Fiyiki, Y. Disruption of the interaction of the longer isoform of Pex5p, Pex5pl, with Pex7p abolishes peroxisome targeting signal type 2 protein import in mammals. Study with a novel Pex5-impaired Chinese hamster ovary cell mutant. J. Biol. Chem. 2000, 275, 21715–21721. [Google Scholar] [CrossRef] [PubMed]
- Woodward, A.W.; Bartel, B. The Arabidopsis peroxisomal targeting signal type 2 receptor PEX7 is necessary for peroxisome function and dependent on PEX5. Mol. Biol. Cell 2005, 16, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Galland, N.; Demeure, F.; Hannaert, V.; Verplaetse, E.; Vertommen, D.; Van Der Smissen, P.; Courtoy, P.J.; Michels, P.A.M. Characterization of the role of the receptors PEX5 and PEX7 in the import of proteins into glycosomes of Trypanosoma brucei. Biochim. Biophys. Acta 2007, 1773, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Khan, B.R.; Zolman, B.K. pex5 Mutants that differentially disrupt PTS1 and PTS2 peroxisomal matrix protein import in Arabidopsis. Plant Physiol. 2010, 154, 1602–1615. [Google Scholar] [CrossRef]
- Ramón, N.M.; Bartel, B. Interdependence of the peroxisome-targeting receptors in Arabidopsis thaliana: PEX7 facilitates PEX5 accumulation and import of PTS1 cargo into peroxisomes. Mol. Biol. Cell 2010, 21, 1263–1271. [Google Scholar] [CrossRef]
- Pan, D.; Nakatsu, T.; Kato, H. Crystal structure of peroxisomal targeting signal-2 bound to its receptor complex Pex7p-Pex21p. Nat. Struct. Mol. Biol. 2013, 20, 987–993. [Google Scholar] [CrossRef]
- Rodrigues, T.A.; Grou, C.P.; Azevedo, J.E. Revisiting the intraperoxisomal pathway of mammalian PEX7. Sci. Rep. 2015, 5, 11806. [Google Scholar] [CrossRef]
- Kunze, M.; Malkani, N.; Maurer-Stroh, S.; Wiesinger, C.; Schmid, J.A.; Berger, J. Mechanistic insights into PTS2-mediated peroxisomal protein import: the co-receptor PEX5L drastically increases the interaction strength between the cargo protein and the receptor PEX7. J. Biol. Chem. 2015, 290, 4928–4940. [Google Scholar] [CrossRef]
- Voorn-Brouwer, T.; van der Leij, I.; Hemrika, W.; Distel, B.; Tabak, H.F. Sequence of the PAS8 gene, the product of which is essential for biogenesis of peroxisomes in Saccharomyces cerevisiae. Biochim. Biophys. Acta 1993, 1216, 325–328. [Google Scholar] [CrossRef]
- Purdue, P.E.; Yang, X.; Lazarow, P.B. Pex18p and Pex21p, a novel pair of related peroxins essential for peroxisomal targeting by the PTS2 pathway. J. Cell Biol. 1998, 143, 1859–1869. [Google Scholar]
- Dodt, G.; Warren, D.; Becker, E.; Rehling, P.; Gould, S.J. Domain mapping of human PEX5 reveals functional and structural similarities to Saccharomyces cerevisiae Pex18p and Pex21p. J. Biol. Chem. 2001, 276, 41769–41781. [Google Scholar] [CrossRef] [PubMed]
- Otzen, M.; Wang, D.; Lunenborg, M.G.J.; Van Der Klei, I.J. Hansenula polymorpha Pex20p is an oligomer that binds the peroxisomal targeting signal 2 (PTS2). J. Cell Sci. 2005, 118, 3409–3418. [Google Scholar] [CrossRef]
- Einwächter, H.; Sowinski, S.; Kunau, W.H.; Schliebs, W. Yarrowia lipolytica Pex20p, Saccharomyces cerevisiae Pex18p/Pex21p and mammalian Pex5pL fulfil a common function in the early steps of the peroxisomal PTS2 import pathway. EMBO Rep. 2001, 2, 1035–1039. [Google Scholar]
- Effelsberg, D.; Cruz-Zaragoza, L.D.; Schliebs, W.; Erdmann, R. Pex9p is a new yeast peroxisomal import receptor for PTS1-containing proteins. J. Cell Sci. 2016, 129, 4057–4066. [Google Scholar] [PubMed]
- Yifrach, E.; Chuartzman, S.G.; Dahan, N.; Maskit, S.; Zada, L.; Weill, U.; Yofe, I.; Olender, T.; Schuldiner, M.; Zalckvar, E. Characterization of proteome dynamics during growth in oleate reveals a new peroxisome-targeting receptor. J. Cell Sci. 2016, 129, 4067–4075. [Google Scholar] [PubMed]
- Schäfer, A.; Kerssen, D.; Veenhuis, M.; Kunau, W.H.; Schliebs, W. Functional similarity between the peroxisomal PTS2 receptor binding protein Pex18p and the N-terminal half of the PTS1 receptor Pex5p. Mol. Cell. Biol. 2004, 24, 8895–8906. [Google Scholar] [PubMed]
- Schliebs, W.; Kunau, W.H. PTS2 Co-receptors: Diverse proteins with common features. Biochim. Biophys. Acta-Mol. Cell Res. 2006, 1763, 1605–1612. [Google Scholar]
- Agne, B.; Meindl, N.M.; Niederhoff, K.; Einwächter, H.; Rehling, P.; Sickmann, A.; Meyer, H.E.; Girzalsky, W.; Kunau, W.H. Pex8p: an intraperoxisomal organizer of the peroxisomal import machinery. Mol. Cell 2003, 11, 635–646. [Google Scholar] [CrossRef]
- Reguenga, C.; Oliveira, M.E.; Gouveia, A.M.; Sá-Miranda, C.; Azevedo, J.E. Characterization of the mammalian peroxisomal import machinery: Pex2p, Pex5p, Pex12p, and Pex14p are subunits of the same protein assembly. J. Biol. Chem. 2001, 276, 29935–29942. [Google Scholar] [CrossRef] [PubMed]
- McNew, J.A.; Goodman, J.M. An oligomeric protein is imported into peroxisomes in vivo. J. Cell Biol. 1994, 127, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Glover, J.R.; Andrews, D.W.; Rachubinski, R.A. Saccharomyces cerevisiae peroxisomal thiolase is imported as a dimer. Proc. Natl. Acad. Sci. USA 1994, 91, 10541–10545. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.Q.; Esposito, R.D.; Gowani, J.; Goodman, J.M. Alcohol oxidase and dihydroxyacetone synthase, the abundant peroxisomal proteins of methylotrophic yeasts, assemble in different cellular compartments. J. Cell Sci. 2001, 114, 2863–2868. [Google Scholar] [PubMed]
- Dias, A.F.; Rodrigues, T.A.; Pedrosa, A.G.; Barros-Barbosa, A.; Francisco, T.; Azevedo, J.E. The peroxisomal matrix protein translocon is a large cavity-forming protein assembly into which PEX5 protein enters to release its cargo. J. Biol. Chem. 2017, 292, 15287–15300. [Google Scholar] [CrossRef]
- Dias, A.F.; Francisco, T.; Rodrigues, T.A.; Grou, C.P.; Azevedo, J.E. The first minutes in the life of a peroxisomal matrix protein. Biochim. Biophys. Acta 2016, 1863, 814–820. [Google Scholar] [CrossRef]
- Barros-Barbosa, A.; Rodrigues, T.A.; Ferreira, M.J.; Pedrosa, A.G.; Teixeira, N.R.; Francisco, T.; Azevedo, J.E. The intrinsically disordered nature of the peroxisomal protein translocation machinery. FEBS J. 2019, 286, 24–38. [Google Scholar] [CrossRef]
- Gouveia, A.M.; Guimarães, C.P.; Oliveira, M.E.; Sá-Miranda, C.; Azevedo, J.E. Insertion of Pex5p into the peroxisomal membrane is cargo protein-dependent. J. Biol. Chem. 2003, 278, 4389–4392. [Google Scholar] [CrossRef]
- Costa-Rodrigues, J.; Carvalho, A.F.; Gouveia, A.M.; Fransen, M.; Sá-Miranda, C.; Azevedo, J.E. The N terminus of the peroxisomal cycling receptor, Pex5p, is required for redirecting the peroxisome-associated peroxin back to the cytosol. J. Biol. Chem. 2004, 279, 46573–46579. [Google Scholar] [CrossRef]
- Gouveia, A.M.; Guimarães, C.P.; Oliveira, M.E.; Reguenga, C.; Sá-Miranda, C.; Azevedo, J.E. Characterization of the peroxisomal cycling receptor, Pex5p, using a cell-free in vitro import system. J. Biol. Chem. 2003, 278, 226–232. [Google Scholar] [CrossRef]
- Rodrigues, T.A.; Alencastre, I.S.; Francisco, T.; Brites, P.; Fransen, M.; Grou, C.P.; Azevedo, J.E. A PEX7-centered perspective on the peroxisomal targeting signal type 2-mediated protein import pathway. Mol. Cell. Biol. 2014, 34, 2917–2928. [Google Scholar] [CrossRef] [PubMed]
- Van Der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of intrinsically disordered regions and proteins. Chem. Rev. 2014, 114, 6589–6631. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, A.; Kooshapur, H.; Wolf, J.; Meyer, N.H.; Madl, T.; Saidowsky, J.; Hambruch, E.; Lazam, A.; Jung, M.; Sattler, M.; et al. A novel Pex14 protein-interacting site of human Pex5 is critical for matrix protein import into peroxisomes. J. Biol. Chem. 2014, 289, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Saidowsky, J.; Dodt, G.; Kirchberg, K.; Wegner, A.; Nastainezyk, W.; Kunau, W.H.; Schliebs, W. The di-aromatic pentapeptide repeats of the human peroxisome import receptor PEX5 are separate high affinity binding sites for the peroxisomal membrane protein PEX14. J. Biol. Chem. 2001, 276, 34524–34529. [Google Scholar] [CrossRef]
- Waterham, H.R.; Titorenko, V.I.; Haima, P.; Cregg, J.M.; Harder, W.; Veenhuis, M. The Hansenula polymorpha PER1 gene is essential for peroxisome biogenesis and encodes a peroxisomal matrix protein with both carboxy- and amino-terminal targeting signals. J. Cell Biol. 1994, 127, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Elgersma, Y.; Kwast, L.; Klein, A.; Voorn-Brouwer, T.; van den Berg, M.; Metzig, B.; America, T.; Tabak, H.F.; Distel, B. The SH3 domain of the Saccharomyces cerevisiae peroxisomal membrane protein Pex13p functions as a docking site for Pex5p, a mobile receptor for the import PTS1-containing proteins. J. Cell Biol. 1996, 135, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Warren, D.S.; Sacksteder, K.A.; Gould, S.J. PEX12 interacts with PEX5 and PEX10 and acts downstream of receptor docking in peroxisomal matrix protein import. J. Cell Biol. 1999, 147, 761–774. [Google Scholar] [CrossRef]
- Rehling, P.; Skaletz-Rorowski, A.; Girzalsky, W.; Voorn-Brouwer, T.; Franse, M.M.; Distel, B.; Veenhuis, M.; Kunau, W.H.; Erdmann, R. Pex8p, an intraperoxisomal peroxin of Saccharomyces cerevisiae required for protein transport into peroxisomes binds the PTS1 receptor pex5p. J. Biol. Chem. 2000, 275, 3593–3602. [Google Scholar] [CrossRef]
- Mano, S.; Nakamori, C.; Nito, K.; Kondo, M.; Nishimura, M. The Arabidopsis pex12 and pex13 mutants are defective in both PTS1- and PTS2-dependent protein transport to peroxisomes. Plant J. 2006, 47, 604–618. [Google Scholar] [CrossRef]
- Freitas, M.O.; Francisco, T.; Rodrigues, T.A.; Alencastre, I.S.; Pinto, M.P.; Grou, C.P.; Carvalho, A.F.; Fransen, M.; Sá-Miranda, C.; Azevedo, J.E. PEX5 protein binds monomeric catalase blocking its tetramerization and releases it upon binding the N-terminal domain of PEX14. J. Biol. Chem. 2011, 286, 40509–40519. [Google Scholar] [CrossRef]
- Lanyon-Hogg, T.; Hooper, J.; Gunn, S.; Warriner, S.L.; Baker, A. PEX14 binding to Arabidopsis PEX5 has differential effects on PTS1 and PTS2 cargo occupancy of the receptor. FEBS Lett. 2014, 588, 2223–2229. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.F.; Pinto, M.P.; Grou, C.P.; Alencastre, I.S.; Fransen, M.; Sá-Miranda, C.; Azevedo, J.E. Ubiquitination of mammalian Pex5p, the peroxisomal import receptor. J. Biol. Chem. 2007, 282, 31267–31272. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.P.; Van Den Berg, M.; Sprenger, R.R.; Distel, B. A conserved cysteine is essential for Pex4p-dependent ubiquitination of the peroxisomal import receptor Pex5p. J. Biol. Chem. 2007, 282, 22534–22543. [Google Scholar] [CrossRef] [PubMed]
- Francisco, T.; Rodrigues, T.A.; Pinto, M.P.; Carvalho, A.F.; Azevedo, J.E.; Grou, C.P. Ubiquitin in the peroxisomal protein import pathway. Biochimie 2014, 98, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Tamura, S.; Fujiki, Y. The pathogenic peroxin Pex26p recruits the Pex1p-Pex6p AAA ATPase complexes to peroxisomes. Nat. Cell Biol. 2003, 5, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Birschmann, I.; Stroobants, A.K.; van den Berg, M.; Schäfer, A.; Rosenkranz, K.; Kunau, W.H.; Tabak, H.F. Pex15p of Saccharomyces cerevisiae provides a molecular basis for recruitment of the AAA peroxin Pex6p to peroxisomal membranes. Mol. Biol. Cell 2003, 14, 2226–2236. [Google Scholar] [CrossRef]
- Miyata, N.; Fujiki, Y. Shuttling mechanism of peroxisome targeting signal type 1 receptor Pex5: ATP-independent import and ATP-dependent export. Mol. Cell. Biol. 2005, 25, 10822–10832. [Google Scholar] [CrossRef]
- Platta, H.W.; Grunau, S.; Rosenkranz, K.; Girzalsky, W.; Erdmann, R. Functional role of the AAA peroxins in dislocation of the cycling PTS1 receptor back to the cytosol. Nat. Cell Biol. 2005, 7, 817–822. [Google Scholar] [CrossRef]
- Goto, S.; Mano, S.; Nakamori, C.; Nishimura, M. Arabidopsis ABERRANT PEROXISOME MORPHOLOGY9 is a peroxin that recruits the PEX1-PEX6 complex to peroxisomes. Plant Cell 2011, 23, 1573–1587. [Google Scholar]
- Pedrosa, A.G.; Francisco, T.; Bicho, D.; Dias, A.F.; Barros-Barbosa, A.; Hagmann, V.; Dodt, G.; Rodrigues, T.A.; Azevedo, J.E. Peroxisomal monoubiquitinated PEX5 interacts with the AAA ATPases PEX1 and PEX6 and is unfolded during its dislocation into the cytosol. J. Biol. Chem. 2018, 293, 11553–11563. [Google Scholar] [CrossRef]
- Grou, C.P.; Carvalho, A.F.; Pinto, M.P.; Alencastre, I.S.; Rodrigues, T.A.; Freitas, M.O.; Francisco, T.; Sá-Miranda, C.; Azevedo, J.E. The peroxisomal protein import machinery--a case report of transient ubiquitination with a new flavor. Cell. Mol. Life Sci. 2009, 66, 254–262. [Google Scholar] [PubMed]
- Erdmann, R.; Wiebel, F.F.; Flessau, A.; Rytka, J.; Beyer, A.; Fröhlich, K.U.; Kunau, W.H. PAS1, a yeast gene required for peroxisome biogenesis, encodes a member of a novel family of putative ATPases. Cell 1991, 64, 499–510. [Google Scholar] [CrossRef]
- Portsteffen, H.; Beyer, A.; Becker, E.; Epplen, C.; Pawlak, A.; Kunau, W.H.; Dodt, G. Human PEX1 is mutated in complementation group 1 of the peroxisome biogenesis disorders. Nat. Genet. 1997, 17, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Reuber, B.E.; Germain-Lee, E.; Collins, C.S.; Morrell, J.C.; Ameritunga, R.; Moser, H.W.; Valle, D.; Gould, S.J. Mutations in PEX1 are the most common cause of peroxisome biogenesis disorders. Nat. Genet. 1997, 17, 445–448. [Google Scholar] [CrossRef]
- Tamura, S.; Okumoto, K.; Toyama, R.; Shimozawa, N.; Tsukamoto, T.; Suzuki, Y.; Osumi, T.; Kondo, N.; Fujiki, Y. Human PEX1 cloned by functional complementation on a CHO cell mutant is responsible for peroxisome-deficient Zellweger syndrome of complementation group I. Proc. Natl. Acad. Sci. USA 1998, 95, 4350–4355. [Google Scholar] [CrossRef]
- Lopez-Huertas, E.; Charlton, W.L.; Johnson, B.; Graham, I.A.; Baker, A. Stress induces peroxisome biogenesis genes. EMBO J. 2000, 19, 6770–6777. [Google Scholar] [CrossRef]
- Zolman, B.K.; Yoder, A.; Bartel, B. Genetic analysis of indole-3-butyric acid responses in Arabidopsis thaliana reveals four mutant classes. Genetics 2000, 156, 1323–1337. [Google Scholar]
- Zolman, B.K.; Bartel, B. An Arabidopsis indole-3-butyric acid-response mutant defective in PEROXIN6, an apparent ATPase implicated in peroxisomal function. Proc. Natl. Acad. Sci. USA 2004, 101, 1786–1791. [Google Scholar]
- Schlüter, A.; Fourcade, S.; Ripp, R.; Mandel, J.L.; Poch, O.; Pujol, A. The evolutionary origin of peroxisomes: An ER-peroxisome connection. Mol. Biol. Evol. 2006, 23, 838–845. [Google Scholar] [CrossRef]
- Gabaldón, T.; Snel, B.; van Zimmeren, F.; Hemrika, W.; Tabak, H.F.; Huynen, M.A. Origin and evolution of the peroxisomal proteome. Biol. Direct 2006, 1, 8. [Google Scholar]
- Waterham, H.R.; Ebberink, M.S. Genetics and molecular basis of human peroxisome biogenesis disorders. Biochim. Biophys. Acta 2012, 1822, 1430–1441. [Google Scholar] [PubMed]
- Argyriou, C.; D’Agostino, M.D.; Braverman, N.E. Peroxisome biogenesis disorders. Transl. Sci. Rare Dis. 2016, 1, 111–144. [Google Scholar] [CrossRef] [PubMed]
- Schieferdecker, A.; Wendler, P. Structural Mapping of Missense Mutations in the Pex1/Pex6 Complex. Int. J. Mol. Sci. 2019, 20, 3756. [Google Scholar] [CrossRef] [PubMed]
- Elgersma, Y.; Snyder, W.B.; Subramani, S.; Kwast, L.; Van Den Berg, M.; Distel, B.; Tabak, H.F. Overexpression of Pex15p, a phosphorylated peroxisomal integral membrane protein required for peroxisome assembly in S.cerevisiae, causes proliferation of the endoplasmic reticulum membrane. EMBO J. 1997, 16, 7326–7341. [Google Scholar]
- Collins, C.S.; Kalish, J.E.; Morrell, J.C.; McCaffery, J.M.; Gould, S.J. The peroxisome biogenesis factors pex4p, pex22p, pex1p, and pex6p act in the terminal steps of peroxisomal matrix protein import. Mol. Cell. Biol. 2000, 20, 7516–7526. [Google Scholar]
- Hanson, P.I.; Whiteheart, S.W. AAA+ proteins: have engine, will work. Nat. Rev. Mol. Cell Biol. 2005, 6, 519–529. [Google Scholar] [CrossRef]
- Imanaka, T.; Small, G.M.; Lazarow, P.B. Translocation of acyl-CoA oxidase into peroxisomes required ATP hydrolysis but not a membrane potential. J. Cell Biol. 1987, 105, 2915–2922. [Google Scholar] [CrossRef]
- Behari, R.; Baker, A. The carboxyl terminus of isocitrate lyase is not essential for import into glyoxysomes in an in vitro system. J. Biol. Chem. 1993, 268, 7315–7322. [Google Scholar]
- Rapp, S.; Soto, U.; Just, W.W. Import of firefly luciferase into peroxisomes of permeabilized Chinese hamster ovary cells: A model system to study peroxisomal protein import in vitro. Exp. Cell Res. 1993, 205, 59–65. [Google Scholar] [CrossRef]
- Wendland, M.; Subramani, S. Cytosol-dependent peroxisomal protein import in a permeabilized cell system. J. Cell Biol. 1993, 120, 675–685. [Google Scholar] [CrossRef]
- Brickner, D.G.; Harada, J.J.; Olsen, L.J. Protein transport into higher plant peroxisomes: In vitro import assay provides evidence for receptor involvement. Plant Physiol. 1997, 113, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.J.; Collins, C.S. Opinion: peroxisomal-protein import: is it really that complex? Nat. Rev. Mol. Cell Biol. 2002, 3, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Titorenko, V.I.; Rachubinski, R.A. Peroxisomal membrane fusion requires two AAA family ATPases, Pex1p and Pex6p. J. Cell Biol. 2000, 150, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Van Der Zand, A.; Gent, J.; Braakman, I.; Tabak, H.F. Biochemically distinct vesicles from the endoplasmic reticulum fuse to form peroxisomes. Cell 2012, 149, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Knoops, K.; De Boer, R.; Kram, A.; Van Der Klei, I.J. Yeast pex1 cells contain peroxisomal ghosts that import matrix proteins upon reintroduction of Pex1. J. Cell Biol. 2015, 211, 955–962. [Google Scholar] [CrossRef]
- Motley, A.M.; Galvin, P.C.; Ekal, L.; Nuttall, J.M.; Hettema, E.H. Reevaluation of the role of Pex1 and dynamin-related proteins in peroxisome membrane biogenesis. J. Cell Biol. 2015, 211, 1041–1056. [Google Scholar] [CrossRef] [PubMed]
- Ogura, T.; Wilkinson, A.J. AAA+ superfamily ATPases: common structure—Diverse function. Genes to Cells 2001, 6, 575–597. [Google Scholar] [CrossRef]
- Sysoeva, T.A. Assessing heterogeneity in oligomeric AAA+ machines. Cell. Mol. Life Sci. 2017, 74, 1001–1018. [Google Scholar] [CrossRef]
- Saffert, P.; Enenkel, C.; Wendler, P. Structure and Function of p97 and Pex1/6 Type II AAA+ Complexes. Front. Mol. Biosci. 2017, 4, 1–13. [Google Scholar] [CrossRef]
- Bard, J.A.; Goodall, E.A.; Greene, E.R.; Jonsson, E.; Dong, K.C.; Martin, A. Structure and Function of the 26S Proteasome. Annu. Rev. Biochem. 2018, 87, 697–724. [Google Scholar] [CrossRef]
- Olivares, A.O.; Baker, T.A.; Sauer, R.T. Mechanistic insights into bacterial AAA+ proteases and protein-remodelling machines. Nat. Rev. Microbiol. 2016, 14, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Enemark, E.J. Fundamental Characteristics of AAA+ Protein Family Structure and Function. Archaea 2016, 9294307. [Google Scholar] [CrossRef] [PubMed]
- Deville, C.; Franke, K.; Mogk, A.; Bukau, B.; Saibil, H.R. Two-Step Activation Mechanism of the ClpB Disaggregase for Sequential Substrate Threading by the Main ATPase Motor. Cell Rep. 2019, 27, 3433–3446.e4. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.L.; Vieux, E.F.; Kalastavadi, T.; Kim, S.; Chen, J.Z.; Baker, T.A. N domain of the Lon AAA+ protease controls assembly and substrate choice. Protein Sci. 2019, 28, 1239–1251. [Google Scholar] [CrossRef]
- Dougan, D.A.; Mogk, A.; Zeth, K.; Turgay, K.; Bukau, B. AAA+ proteins and substrate recognition, it all depends on their partner in crime. FEBS Lett. 2002, 529, 6–10. [Google Scholar] [CrossRef]
- Stach, L.; Freemont, P.S. The AAA+ ATPase p97, a cellular multitool. Biochem. J. 2017, 474, 2953–2976. [Google Scholar] [CrossRef]
- Zhao, C.; Smith, E.C.; Whiteheart, S.W. Requirements for the catalytic cycle of the N-ethylmaleimide-Sensitive Factor (NSF). Biochim. Biophys. Acta 2012, 1823, 159–171. [Google Scholar] [CrossRef]
- Sweeny, E.A.; Shorter, J. Mechanistic and Structural Insights into the Prion-Disaggregase Activity of Hsp104. J. Mol. Biol. 2016, 428, 1870–1885. [Google Scholar] [CrossRef]
- Wendler, P.; Ciniawsky, S.; Kock, M.; Kube, S. Structure and function of the AAA+ nucleotide binding pocket. Biochim. Biophys. Acta 2012, 1823, 2–14. [Google Scholar] [CrossRef]
- Hinnerwisch, J.; Fenton, W.A.; Furtak, K.J.; Farr, G.W.; Horwich, A.L. Loops in the central channel of ClpA chaperone mediate protein binding, unfolding, and translocation. Cell 2005, 121, 1029–1041. [Google Scholar] [CrossRef]
- DeLaBarre, B.; Christianson, J.C.; Kopito, R.R.; Brunger, A.T. Central pore residues mediate the p97/VCP activity required for ERAD. Mol. Cell 2006, 22, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Roll-Mecak, A.; Vale, R.D. Structural basis of microtubule severing by the hereditary spastic paraplegia protein spastin. Nature 2008, 451, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Matveeva, E.A.; Ren, Q.; Whiteheart, S.W. Dissecting the N-ethylmaleimide-sensitive factor: required elements of the N and D1 domains. J. Biol. Chem. 2010, 285, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Cipriano, D.J.; Jung, J.; Vivona, S.; Fenn, T.D.; Brunger, A.T.; Bryant, Z. Processive ATP-driven substrate disassembly by the N-ethylmaleimide- sensitive factor (NSF) molecular machine. J. Biol. Chem. 2013, 288, 23436–23445. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Wu, S.; Zhou, Q.; Vivona, S.; Cipriano, D.J.; Cheng, Y.; Brunger, A.T. Mechanistic insights into the recycling machine of the SNARE complex. Nature 2015, 518, 61–67. [Google Scholar] [CrossRef]
- Zhao, M.; Brunger, A.T. Recent Advances in Deciphering the Structure and Molecular Mechanism of the AAA+ ATPase N-Ethylmaleimide-Sensitive Factor (NSF). J. Mol. Biol. 2016, 428, 1912–1926. [Google Scholar] [CrossRef] [PubMed]
- White, K.I.; Zhao, M.; Choi, U.B.; Pfuetzner, R.A.; Brunger, A.T. Structural principles of SNARE complex recognition by the AAA+ protein NSF. Elife 2018, 7, e38888. [Google Scholar] [CrossRef]
- van den Boom, J.; Meyer, H. VCP/p97-Mediated Unfolding as a Principle in Protein Homeostasis and Signaling. Mol. Cell 2018, 69, 182–194. [Google Scholar] [CrossRef]
- Twomey, E.C.; Ji, Z.; Wales, T.E.; Bodnar, N.O.; Ficarro, S.B.; Marto, J.A.; Engen, J.R.; Rapoport, T.A. Substrate processing by the Cdc48 ATPase complex is initiated by ubiquitin unfolding. Science 2019, 365, eaax1033. [Google Scholar] [CrossRef]
- Cooney, I.; Han, H.; Stewart, M.G.; Carson, R.H.; Hansen, D.T.; Iwasa, J.H.; Price, J.C.; Hill, C.P.; Shen, P.S. Structure of the Cdc48 segregase in the act of unfolding an authentic substrate. Science 2019, 365, 502–505. [Google Scholar] [CrossRef]
- Shiozawa, K.; Maita, N.; Tomii, K.; Seto, A.; Goda, N.; Akiyama, Y.; Shimizu, T.; Shirakawa, M.; Hiroaki, H. Structure of the N-terminal domain of PEX1 AAA-ATPase. Characterization of a putative adaptor-binding domain. J. Biol. Chem. 2004, 279, 50060–50068. [Google Scholar] [CrossRef] [PubMed]
- Ciniawsky, S.; Grimm, I.; Saffian, D.; Girzalsky, W.; Erdmann, R.; Wendler, P. Molecular snapshots of the Pex1/6 AAA+ complex in action. Nat. Commun. 2015, 6, 7331. [Google Scholar] [CrossRef] [PubMed]
- Gardner, B.M.; Chowdhury, S.; Lander, G.C.; Martin, A. The Pex1/Pex6 complex is a heterohexameric AAA+ motor with alternating and highly coordinated subunits. J. Mol. Biol. 2015, 427, 1375–1388. [Google Scholar] [CrossRef]
- Tsukamoto, T.; Miura, S.; Nakai, T.; Yokota, S.; Shimozawa, N.; Suzuki, Y.; Orii, T.; Fujiki, Y.; Sakai, F.; Bogaki, A.; et al. Peroxisome assembly factor-2, a putative ATPase cloned by functional complementation on a peroxisome-deficient mammalian cell mutant. Nat. Genet. 1995, 11, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Tamura, S.; Shimozawa, N.; Suzuki, Y.; Tsukamoto, T.; Osumi, T.; Fujiki, Y. A cytoplasmic AAA family peroxin, Pex1p, interacts with Pex6p. Biochem. Biophys. Res. Commun. 1998, 245, 883–886. [Google Scholar] [CrossRef]
- Yahraus, T.; Braverman, N.E.; Dodt, G.; Kalish, J.E.; Morrell, J.C.; Moser, H.W.; Valle, D.; Gould, S.J. The peroxisome biogenesis disorder group 4 gene, PXAAA1, encodes a cytoplasmic ATPase required for stability of the PTS1 receptor. EMBO J. 1996, 15, 2914–2923. [Google Scholar] [CrossRef]
- Blok, N.B.; Tan, D.; Wang, R.Y.-R.; Penczek, P.A.; Baker, D.; DiMaio, F.; Rapoport, T.A.; Walz, T. Unique double-ring structure of the peroxisomal Pex1/Pex6 ATPase complex revealed by cryo-electron microscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E4017–E4025. [Google Scholar] [CrossRef]
- Hänzelmann, P.; Schindelin, H. Structural Basis of ATP Hydrolysis and Intersubunit Signaling in the AAA+ ATPase p97. Structure 2016, 24, 127–139. [Google Scholar] [CrossRef]
- Gardner, B.M.; Castanzo, D.T.; Chowdhury, S.; Stjepanovic, G.; Stefely, M.S.; Hurley, J.H.; Lander, G.C.; Martin, A. The peroxisomal AAA-ATPase Pex1/Pex6 unfolds substrates by processive threading. Nat. Commun. 2018, 9, 135. [Google Scholar] [CrossRef]
- Birschmann, I.; Rosenkranz, K.; Erdmann, R.; Kunau, W.H. Structural and functional analysis of the interaction of the AAA-peroxins Pex1p and Pex6p. FEBS J. 2005, 272, 47–58. [Google Scholar] [CrossRef]
- Tamura, S.; Yasutake, S.; Matsumoto, N.; Fujiki, Y. Dynamic and functional assembly of the AAA peroxins, Pex1p and Pex6p, and their membrane receptor Pex26p. J. Biol. Chem. 2006, 281, 27693–27704. [Google Scholar] [CrossRef] [PubMed]
- Tamura, S.; Matsumoto, N.; Takebas, R.; Fujiki, Y. AAA peroxins and their recruiter Pex26p modulate the interactions of peroxins involved in peroxisomal protein import. J. Biol. Chem. 2014, 289, 24336–24346. [Google Scholar] [CrossRef] [PubMed]
- Grimm, I.; Saffian, D.; Girzalsky, W.; Erdmann, R. Nucleotide-dependent assembly of the peroxisomal receptor export complex. Sci. Rep. 2016, 6, 19838. [Google Scholar] [CrossRef] [PubMed]
- Erzberger, J.P.; Berger, J.M. Evolutionary relationships and structural mechanisms of AAA+ proteins. Annu. Rev. Biophys. Biomol. Struct. 2006, 35, 93–114. [Google Scholar] [CrossRef] [PubMed]
- Hagmann, V.; Sommer, S.; Fabian, P.; Bierlmeier, J.; van Treel, N.; Mootz, H.D.; Schwarzer, D.; Azevedo, J.E.; Dodt, G. Chemically monoubiquitinated PEX5 binds to the components of the peroxisomal docking and export machinery. Sci. Rep. 2018, 8, 16014. [Google Scholar] [CrossRef]
- Schwerter, D.P.; Grimm, I.; Girzalsky, W.; Erdmann, R. Receptor recognition by the peroxisomal AAA complex depends on the presence of the ubiquitin moiety and is mediated by Pex1p. J. Biol. Chem. 2018, 293, 15458–15470. [Google Scholar] [CrossRef]
- Urquhart, A.J.; Kennedy, D.; Gould, S.J.; Crane, D.I. Interaction of Pex5p, the type 1 peroxisome targeting signal receptor, with the peroxisomal membrane proteins Pex14p and Pex13p. J. Biol. Chem. 2000, 275, 4127–4136. [Google Scholar] [CrossRef]
- Fransen, M.; Brees, C.; Ghys, K.; Amery, L.; Mannaerts, G.P.; Ladant, D.; Van Veldhoven, P.P. Analysis of mammalian peroxin interactions using a non-transcription-based bacterial two-hybrid assay. Mol. Cell. Proteomics 2002, 1, 243–252. [Google Scholar] [CrossRef]
- Barros-Barbosa, A.; Ferreira, M.J.M.J.; Rodrigues, T.A.; Pedrosa, A.G.; Grou, C.P.; Pinto, M.P.M.P.; Fransen, M.; Francisco, T.; Azevedo, J.E. Membrane topologies of PEX13 and PEX14 provide new insights on the mechanism of protein import into peroxisomes. FEBS J. 2019, 286, 205–222. [Google Scholar] [CrossRef]
- Nordgren, M.; Francisco, T.; Lismont, C.; Hennebel, L.; Brees, C.; Wang, B.; Van Veldhoven, P.P.; Azevedo, J.E.; Fransen, M. Export-deficient monoubiquitinated PEX5 triggers peroxisome removal in SV40 large T antigen-transformed mouse embryonic fibroblasts. Autophagy 2015, 11, 1326–1340. [Google Scholar] [CrossRef]
- Johnston, J.A.; Johnson, E.S.; Waller, P.R.; Varshavsky, A. Methotrexate inhibits proteolysis of dihydrofolate reductase by the N-end rule pathway. J. Biol. Chem. 1995, 270, 8172–8178. [Google Scholar] [CrossRef]
- Yedidi, R.S.; Wendler, P.; Enenkel, C. AAA-ATPases in Protein Degradation. Front. Mol. Biosci. 2017, 4, 1–14. [Google Scholar] [CrossRef]
- Johjima, A.; Noi, K.; Nishikori, S.; Ogi, H.; Esaki, M.; Ogura, T. Microtubule severing by katanin p60 AAA+ATPase requires the C-terminal acidic tails of both α-and β-tubulins and basic amino acid residues in the AAA+ring pore. J. Biol. Chem. 2015, 290, 11762–11770. [Google Scholar] [CrossRef]
- Zehr, E.A.; Szyk, A.; Piszczek, G.; Szczesna, E.; Zuo, X.; Roll-Mecak, A. Katanin spiral and ring structures shed light on power stroke for microtubule severing. Nat. Struct. Mol. Biol. 2017, 24, 717–725. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Yu, H.; Mim, C.; Matouschek, A. Regulated protein turnover: snapshots of the proteasome in action. Nat. Rev. Mol. Cell Biol. 2014, 15, 122–133. [Google Scholar] [CrossRef]
- Han, H.; Fulcher, J.M.; Dandey, V.P.; Iwasa, J.H.; Sundquist, W.I.; Kay, M.S.; Shen, P.S.; Hill, C.P. Structure of Vps4 with circular peptides and implications for translocation of two polypeptide chains by AAA+ ATPases. Elife 2019, 8, e44071. [Google Scholar] [CrossRef]
- Blythe, E.E.; Olson, K.C.; Chau, V.; Deshaies, R.J. Ubiquitin- and ATP-dependent unfoldase activity of P97/VCP•NPLOC4•UFD1L is enhanced by a mutation that causes multisystem proteinopathy. Proc. Natl. Acad. Sci. USA 2017, 114, E4380–E4388. [Google Scholar] [CrossRef]
- Bodnar, N.O.; Rapoport, T.A. Molecular Mechanism of Substrate Processing by the Cdc48 ATPase Complex. Cell 2017, 169, 722–735. [Google Scholar] [CrossRef]
- Monroe, N.; Hill, C.P. Meiotic Clade AAA ATPases: Protein Polymer Disassembly Machines. J. Mol. Biol. 2016, 428, 1897–1911. [Google Scholar] [CrossRef]
- Saffian, D.; Grimm, I.; Girzalsky, W.; Erdmann, R. ATP-dependent assembly of the heteromeric Pex1p-Pex6p-complex of the peroxisomal matrix protein import machinery. J. Struct. Biol. 2012, 179, 126–132. [Google Scholar] [CrossRef]
- Bailey, M.E.; Jiang, N.; Dima, R.I.; Ross, J.L. Invited review: Microtubule severing enzymes couple atpase activity with tubulin GTPase spring loading. Biopolymers 2016, 105, 547–556. [Google Scholar] [CrossRef]
- Lu, C.; Turley, S.; Marionni, S.T.; Park, Y.J.; Lee, K.K.; Patrick, M.; Shah, R.; Sandkvist, M.; Bush, M.F.; Hol, W.G.J. Hexamers of the type II secretion ATPase GspE from Vibrio cholerae with increased ATPase activity. Structure 2013, 21, 1707–1717. [Google Scholar] [CrossRef]
- Hartman, J.J.; Vale, R.D. Microtubule disassembly by ATP-dependent oligomerization of the AAA enzyme katanin. Science 1999, 286, 782–785. [Google Scholar] [CrossRef]
- Olivares, A.O.; Nager, A.R.; Iosefson, O.; Sauer, R.T.; Baker, T.A. Mechanochemical basis of protein degradation by a double-ring AAA+ machine. Nat. Struct. Mol. Biol. 2014, 21, 871–875. [Google Scholar] [CrossRef]
- Driessen, A.J. Precursor protein translocation by the Escherichia coli translocase is directed by the protonmotive force. EMBO J. 1992, 11, 847–853. [Google Scholar] [CrossRef]
- Alder, N.N.; Theg, S.M. Energetics of protein transport across biological membranes. a study of the thylakoid DeltapH-dependent/cpTat pathway. Cell 2003, 112, 231–242. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedrosa, A.G.; Francisco, T.; Ferreira, M.J.; Rodrigues, T.A.; Barros-Barbosa, A.; Azevedo, J.E. A Mechanistic Perspective on PEX1 and PEX6, Two AAA+ Proteins of the Peroxisomal Protein Import Machinery. Int. J. Mol. Sci. 2019, 20, 5246. https://doi.org/10.3390/ijms20215246
Pedrosa AG, Francisco T, Ferreira MJ, Rodrigues TA, Barros-Barbosa A, Azevedo JE. A Mechanistic Perspective on PEX1 and PEX6, Two AAA+ Proteins of the Peroxisomal Protein Import Machinery. International Journal of Molecular Sciences. 2019; 20(21):5246. https://doi.org/10.3390/ijms20215246
Chicago/Turabian StylePedrosa, Ana G., Tânia Francisco, Maria J. Ferreira, Tony A. Rodrigues, Aurora Barros-Barbosa, and Jorge E. Azevedo. 2019. "A Mechanistic Perspective on PEX1 and PEX6, Two AAA+ Proteins of the Peroxisomal Protein Import Machinery" International Journal of Molecular Sciences 20, no. 21: 5246. https://doi.org/10.3390/ijms20215246
APA StylePedrosa, A. G., Francisco, T., Ferreira, M. J., Rodrigues, T. A., Barros-Barbosa, A., & Azevedo, J. E. (2019). A Mechanistic Perspective on PEX1 and PEX6, Two AAA+ Proteins of the Peroxisomal Protein Import Machinery. International Journal of Molecular Sciences, 20(21), 5246. https://doi.org/10.3390/ijms20215246