Importance of the Interaction between Heading Date Genes Hd1 and Ghd7 for Controlling Yield Traits in Rice



Abstract
:1. Introduction
2. Results
2.1. Effects of Hd1 and Ghd7 on Heading Date
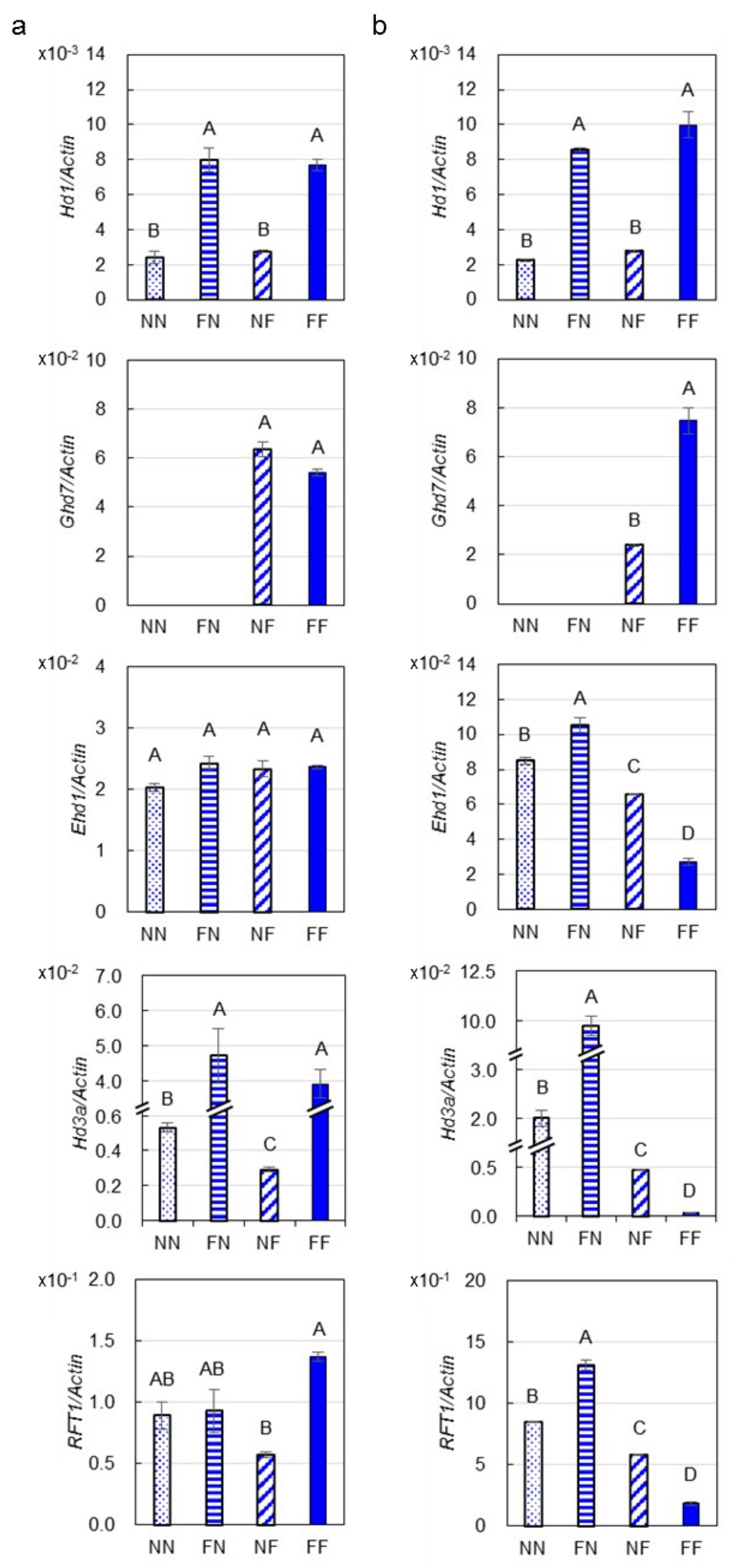
2.2. Expressions of Genes Involved in the Photoperiod Pathway
2.3. Influence of Hd1 and Ghd7 on Yield Traits and Its Relationship with HD
2.4. Validation of the Influences of Hd1 and Ghd7 on HD and Yield Traits under NLD Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Field Experiments and Phenotyping
4.3. DNA Marker Genotyping and Quantitative Real-time PCR Analysis
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| NSDs | Natural short-day conditions |
| NLDs | Natural long-day conditions |
| SD | Short-day conditions |
| LD | Long-day conditions |
| NIL | Near isogenic lines |
| ZS97 | Zhenshan 97 |
| MY46 | Milyang 46 |
| HD | Heading date |
| QTL | Quantitative trait locus |
| R2 | The proportion of phenotypic variance explained |
| GY | Grain yield per plant |
| NP | Number of panicles per plant |
| NSP | Number of spikelets per panicle |
| NGP | Number of grains per panicle |
| SF | Spikelet fertility |
| TGW | 1000-grain weight |
| r | Correlation coefficient |
References
- Tamaki, S.; Matsuo, S.; Wong, H.L.; Yokoi, S.; Shimamoto, K. Hd3a protein is a mobile flowering signal in rice. Science 2007, 316, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Komiya, R.; Yokoi, S.; Shimamoto, K. A gene network for long-day flowering activates RFT1 encoding a mobile flowering signal in rice. Development 2009, 136, 3443–3450. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Matsubara, K.; Yano, M. Genetic control of flowering time in rice: Integration of Mendelian genetics and genomics. Theor. Appl. Genet. 2016, 129, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Izawa, T.; Oikawa, T.; Sugiyama, N.; Tanisaka, T.; Yano, M.; Shimamoto, K. Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice. Genes Dev. 2002, 16, 2006–2020. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Aoki, M.; Kurotani, K.; Yokoi, S.; Shinomura, T.; Takano, M.; Shimamoto, K. Phytochrome B regulates Heading date 1 (Hd1)-mediated expression of rice florigen Hd3a and critical day length in rice. Mol. Genet. Genomics 2011, 285, 461–470. [Google Scholar] [CrossRef]
- Nemoto, Y.; Nonoue, Y.; Yano, M.; Izawa, T. Hd1, a CONSTANS ortholog in rice, functions as an Ehd1 repressor through interaction with monocot-specific CCT-domain protein Ghd7. Plant J. 2016, 86, 221–233. [Google Scholar] [CrossRef]
- Du, A.; Tian, W.; Wei, M.; Yan, W.; He, H.; Zhou, D.; Huang, X.; Li, S.; Ouyang, X. The DTH8-Hd1 module mediates day-length-dependent regulation of rice flowering. Mol. Plant 2017, 10, 948–961. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, W.; Shen, G.; Liu, H.; Hu, Y.; Zhou, X.; Liu, T.; Xing, Y. Alternative functions of Hd1 in repressing or promoting heading are determined by Ghd7 status under long-day conditions. Sci. Rep. 2017, 7, 5388. [Google Scholar] [CrossRef]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef]
- Wei, X.; Xu, J.; Guo, H.; Jiang, L.; Chen, S.; Yu, C.; Zhou, Z.; Hu, P.; Zhai, H.; Wan, J. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 2010, 153, 1747–1758. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.H.; Wang, P.; Chen, H.X.; Zhou, H.J.; Li, Q.P.; Wang, C.R.; Ding, Z.H.; Zhang, Y.S.; Yu, S.B.; Xing, Y.Z.; et al. A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol. Plant 2011, 4, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Endo-Higashi, N.; Izawa, T. Flowering time genes Heading date 1 and Early heading date 1 together control panicle development in rice. Plant Cell Physiol. 2011, 52, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-H.; Wang, K.; Guo, L.; Zhu, Y.-J.; Fan, Y.-Y.; Cheng, S.-H.; Zhuang, J.-Y. Pleiotropism of the photoperiod-insensitive allele of Hd1 on heading date, plant height and yield traits in rice. PLoS ONE 2012, 7, e52538. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Liu, H.; Zhou, X.; Li, Q.; Zhang, J.; Lu, L.; Liu, T.; Liu, H.; Zhang, C.; Zhang, Z.; et al. Natural variation in Ghd7.1 plays an important role in grain yield and adaptation in rice. Cell Res. 2013, 23, 969–971. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Jin, M.; Zheng, X.-M.; Chen, J.; Yuan, D.; Xin, Y.; Wang, M.; Huang, D.; Zhang, Z.; Zhou, K.; et al. Days to heading 7, a major quantitative locus determining photoperiod sensitivity and regional adaptation in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 16337–16342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-H.; Cao, L.-Y.; Chen, J.-Y.; Zhang, Y.-X.; Zhuang, J.-Y.; Cheng, S.-H. Effects of Hd2 in the presence of the photoperiod-insensitive functional allele of Hd1 in rice. Biol. Open 2016, 5, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-J.; Fan, Y.-Y.; Wang, K.; Huang, D.-R.; Liu, W.-Z.; Ying, J.-Z.; Zhuang, J.-Y. Rice Flowering Locus T 1 plays an important role in heading date influencing yield traits in rice. Sci. Rep. 2017, 7, 4918. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Guo, L.; Ma, H.; Chen, Y.-Y.; Zhang, H.-W.; Ying, J.-Z.; Zhuang, J.-Y. Fine mapping of qHd1, a minor heading date QTL with pleiotropism for yield traits in rice (Oryza sativa L.). Theor. Appl. Genet. 2014, 127, 2515–2524. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Zhang, H.-W.; Zhang, H.-L.; Ying, J.-Z.; Ma, L.-Y.; Zhuang, J.-Y. Natural variation at qHd1 affects heading date acceleration at high temperatures with pleiotropism for yield traits in rice. BMC Plant Biol. 2018, 18, 112. [Google Scholar] [CrossRef]
- Weng, X.; Wang, L.; Wang, J.; Hu, Y.; Du, H.; Xu, C.; Xing, Y.; Li, X.; Xiao, J.; Zhang, Q. Grain number, plant height, and heading date7 is a central regulator of growth, development, and stress response. Plant Physiol. 2014, 164, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, J.; Zhao, T.; Gomez, A.; Li, C.; Yu, C.; Li, H.; Lin, J.; Yang, Y.; Liu, B.; et al. A drought-inducible transcription factor delays reproductive timing in rice. Plant Physiol. 2016, 171, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shen, J.; Xu, Y.; Li, X.; Xiao, J.; Xiong, L. Ghd2, a CONSTANS-like gene, confers drought sensitivity through regulation of senescence in rice. J. Exp. Bot. 2016, 67, 5785–5798. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Shan, J.X.; Gao, J.P.; Lin, H.X. OsHAL3, a blue light-responsive protein, interacts with the floral regulator Hd1 to activate flowering in rice. Mol. Plant 2016, 9, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Minh-Thu, P.T.; Kim, J.S.; Chae, S.; Jun, K.M.; Lee, G.S.; Kim, D.E.; Cheong, J.J.; Song, S.I.; Nahm, B.H.; Kim, Y.K. A WUSCHEL homeobox transcription factor, OsWOX13, enhances drought tolerance and triggers early flowering in rice. Mol. Cells 2018, 41, 781–798. [Google Scholar] [PubMed]
- Song, T.; Zhang, Q.; Wang, H.; Han, J.; Xu, Z.; Yan, S.; Zhu, Z. OsJMJ703, a rice histone demethylase gene, plays key roles in plant development and responds to drought stress. Plant Physiol. Biochem. 2018, 132, 183–188. [Google Scholar] [CrossRef]
- Yano, M.; Katayose, Y.; Ashikari, M.; Yamanouchi, U.; Monna, L.; Fuse, T.; Baba, T.; Yamamoto, K.; Umehara, Y.; Nagamura, Y.; et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 2000, 12, 2473–2484. [Google Scholar] [CrossRef]
- Wei, X.-J.; Xu, J.-F.; Jiang, L.; Wang, J.-J.; Zhou, Z.-L.; Zhai, H.-Q.; Wan, J.-M. Genetic analysis for the diversity of heading date of cultivated rice in China. Acta Agron. Sin. 2012, 38, 10–12. [Google Scholar] [CrossRef]
- Ye, J.; Niu, X.; Yang, Y.; Wang, S.; Xu, Q.; Yuan, X.; Yu, H.; Wang, Y.; Wang, S.; Feng, Y.; et al. Divergent Hd1, Ghd7, and DTH7 alleles control heading date and yield potential of japonica rice in northeast China. Front. Plant Sci. 2018, 9, 35. [Google Scholar] [CrossRef]
- Hu, Y.; Li, S.; Xing, Y. Lessons from natural variations: Artificially induced heading date variations for improvement of regional adaptation in rice. Theor. Appl. Genet. 2018. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, W.; Yin, Z.; Wen, C.K. Rice CONSTITUTIVE TRIPLE-RESPONSE2 is involved in the ethylene-receptor signalling and regulation of various aspects of rice growth and development. J. Exp. Bot. 2013, 64, 4863–4875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Saito, H.; Hirose, I.; Katsura, K.; Yoshitake, Y.; Yokoo, T.; Tsukiyama, T.; Teraishi, M.; Tanisaka, T.; Okumoto, Y. The effects of the photoperiod-insensitive alleles, se13, hd1 and ghd7, on yield components in rice. Mol. Breed. 2014, 33, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-W.; Fan, Y.-Y.; Zhu, Y.-J.; Chen, J.-Y.; Yu, S.-B.; Zhuang, J.-Y. Dissection of the qTGW1.1 region into two tightly-linked minor QTLs having stable effects for grain weight in rice. BMC Genet. 2016, 17, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.L.; Huang, N.; Bennett, J.; Khush, G.S. PCR-Based Marker-Assisted Selection in Rice Breeding; IRRI Discussion Paper Series No. 12; International Rice Research Institute: Los Banos, CA, USA, 1995. [Google Scholar]
- Chen, X.; Temnykh, S.; Xu, Y.; Cho, Y.G.; McCouch, S.R. Development of a microsatellite framework map providing genome-wide coverage in rice. Theor. Appl. Genet. 1997, 95, 553–567. [Google Scholar] [CrossRef]
- Dong, Q.; Zhang, Z.-H.; Wang, L.-L.; Zhu, Y.-J.; Fan, Y.-Y.; Mou, T.-M.; Ma, L.-Y.; Zhuang, J.-Y. Dissection and fine-mapping of two QTL for grain size linked in a 460-kb region on chromosome 1 of rice. Rice 2018, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Basten, C.J.; Zeng, Z.-B. Windows QTL Cartographer 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2012. [Google Scholar]
- SAS Institute Inc. SAS/STAT User’s Guide; SAS Institute: Cary, NC, USA, 1999. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Population | Trial | Trait | Hd1 | Ghd7 | Hd1 × Ghd7 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P | A | R2% | P | A | R2% | P | I-effect | R2% | |||
| R1-NIL | 16LS | HD | <0.0001 | 10.09 | 80.74 | <0.0001 | 0.51 | 5.79 | <0.0001 | −1.30 | 1.30 |
| 17LS | HD | <0.0001 | 7.95 | 75.69 | <0.0001 | 0.77 | 6.50 | 0.2586 | |||
| NP | 0.5940 | 0.0014 | −0.44 | 7.51 | 0.0103 | ||||||
| NSP | <0.0001 | 7.57 | 35.99 | <0.0001 | 4.35 | 20.36 | 0.0438 | ||||
| NGP | <0.0001 | 7.60 | 41.05 | <0.0001 | 3.62 | 17.38 | 0.1490 | ||||
| SF | 0.0015 | 0.94 | 10.18 | 0.7543 | 0.1061 | ||||||
| TGW | <0.0001 | 0.99 | 51.36 | <0.0001 | 0.32 | 11.31 | 0.0479 | ||||
| GY | <0.0001 | 3.26 | 45.02 | 0.0122 | 0.0575 | ||||||
| 17HZ | HD | <0.0001 | −3.30 | 3.03 | <0.0001 | 6.08 | 56.54 | <0.0001 | 3.06 | 16.43 | |
| NP | <0.0001 | 0.72 | 4.93 | <0.0001 | −1.08 | 20.84 | 0.3167 | ||||
| NSP | 0.0442 | <0.0001 | 5.45 | 13.87 | 0.2970 | ||||||
| NGP | 0.1371 | 0.0677 | 0.3892 | ||||||||
| SF | 0.7773 | 0.0198 | 0.8471 | ||||||||
| TGW | 0.9515 | <0.0001 | 1.03 | 38.44 | 0.0002 | 0.46 | 3.49 | ||||
| GY | 0.4677 | 0.8271 | 0.5233 | ||||||||
| R2-NIL | 18HZ | HD | <0.0001 | −2.34 | 6.60 | <0.0001 | 6.15 | 62.57 | <0.0001 | 4.20 | 28.27 |
| NP | 0.8453 | 0.0304 | <0.0001 | −0.45 | 9.97 | ||||||
| NSP | 0.0253 | <0.0001 | 9.50 | 43.27 | <0.0001 | 4.07 | 7.81 | ||||
| NGP | 0.8697 | <0.0001 | 4.05 | 16.34 | 0.0610 | ||||||
| SF | <0.0001 | 0.99 | 3.10 | <0.0001 | −3.25 | 46.28 | <0.0001 | −1.80 | 14.12 | ||
| TGW | 0.5604 | <0.0001 | 0.33 | 24.38 | <0.0001 | 0.29 | 18.28 | ||||
| GY | 0.0097 | 0.66 | 2.68 | 0.6200 | <0.0001 | −1.14 | 7.33 | ||||
| Population | Trial | Group | HD | NP | NSP | NGP | SF | TGW | GY |
|---|---|---|---|---|---|---|---|---|---|
| R1-NIL | 17LS | FN | 87.2 ± 2.6 Dd | 11.8 ± 1.1 ABb | 79.3 ± 4.4 Cc | 69.3 ± 4.7 Cd | 87.4 ± 3.1 Bb | 25.4 ± 1.1 Cd | 20.7 ± 2.8 Cc |
| FF | 91.2 ± 3.8 Cc | 11.6 ± 1.1 ABb | 87.9 ± 6.9 Bb | 77.6 ± 5.9 Bc | 88.3 ± 2.6 ABb | 27.0 ± 1.0 Bc | 24.0 ± 2.9 Bb | ||
| NN | 103.0 ± 3.3 Bb | 12.6 ± 0.9 Aa | 92.0 ± 5.8 Bb | 83.3 ± 5.5 Bb | 90.6 ± 2.4 Aa | 28.2 ± 0.9 Ab | 29.4 ± 2.6 Aa | ||
| NF | 108.6 ± 2.5 Aa | 11.0 ± 0.8 Bb | 108.2 ± 7.2 Aa | 96.7 ± 7.0 Aa | 89.3 ± 2.4 ABab | 28.9 ± 1.0 Aa | 29.9 ± 3.0 Aa | ||
| 17HZ | FN | 75.0 ± 1.6 Dd | 15.5 ± 1.2 ABa | 107.6 ± 8.6 ABb | 89.0 ± 5.7 Aab | 82.8 ± 3.9 Aa | 22.8 ± 0.8 Cb | 30.4 ± 1.5 Aa | |
| FF | 78.5 ± 2.1 Cc | 16.1 ± 1.2 Aa | 100.1 ± 6.8 Bc | 83.5 ± 4.0 Ab | 83.6 ± 3.9 Aa | 23.8 ± 0.8 BCb | 30.5 ± 1.8 Aa | ||
| NN | 83.7 ± 1.5 Bb | 14.5 ± 1.2 BCb | 112.6 ± 9.1 Aab | 89.8 ± 9.1 Aab | 79.7 ± 5.7 Aa | 24.9 ± 1.1 ABa | 30.9 ± 3.1 Aa | ||
| NF | 92.5 ± 1.6 Aa | 13.1 ± 1.4 Cc | 115.0 ± 10.7 Aa | 91.3 ± 11.8 Aa | 79.6 ± 8.5 Aa | 25.8 ± 1.2 Aa | 29.9 ± 4.5 Aa | ||
| R2-NIL | 18HZ | FN | 81.6 ± 1.2 Dd | 12.6 ± 1.0 Aa | 115.4 ± 5.1 Dd | 105.3 ± 4.9 Bb | 91.3 ± 1.6 Aa | 25.0 ± 0.3 Cc | 31.8 ± 2.9 ABb |
| FF | 86.0 ± 1.4 Cc | 11.7 ± 0.9 BCbc | 120.4 ± 6.7 Cc | 107.6 ± 5.9 Bb | 89.3 ± 2.1 Bb | 25.6 ± 0.4 Bb | 30.8 ± 2.9 Bbc | ||
| NN | 90.0 ± 1.0 Bb | 12.2 ± 1.2 ABab | 131.5 ± 8.8 Bb | 113.5 ± 7.9 Aa | 86.4 ± 2.1 Cc | 25.7 ± 0.6 Bb | 33.3 ± 2.7 Aa | ||
| NF | 102.5 ± 0.7 Aa | 11.3 ± 1.2 Cc | 142.7 ± 7.5 Aa | 115.7 ± 6.7 Aa | 81.1 ± 2.7 Dd | 26.3 ± 0.5 Aa | 29.8 ± 3.2 Bc |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-H.; Zhu, Y.-J.; Wang, S.-L.; Fan, Y.-Y.; Zhuang, J.-Y. Importance of the Interaction between Heading Date Genes Hd1 and Ghd7 for Controlling Yield Traits in Rice. Int. J. Mol. Sci. 2019, 20, 516. https://doi.org/10.3390/ijms20030516
Zhang Z-H, Zhu Y-J, Wang S-L, Fan Y-Y, Zhuang J-Y. Importance of the Interaction between Heading Date Genes Hd1 and Ghd7 for Controlling Yield Traits in Rice. International Journal of Molecular Sciences. 2019; 20(3):516. https://doi.org/10.3390/ijms20030516
Chicago/Turabian StyleZhang, Zhen-Hua, Yu-Jun Zhu, Shi-Lin Wang, Ye-Yang Fan, and Jie-Yun Zhuang. 2019. "Importance of the Interaction between Heading Date Genes Hd1 and Ghd7 for Controlling Yield Traits in Rice" International Journal of Molecular Sciences 20, no. 3: 516. https://doi.org/10.3390/ijms20030516