The premise of utilizing CK compounds in an attempt to induce ADSCs toward a neural linage was based on several factors pertaining to their chemical properties as well as previous biological observations. Firstly, based on the chemical properties, ketamines are a natural class of sulfur and nitrogen-containing cyclic compounds with reductive activity [
15] which are key features of the previously studied simple chemical neurogenic inducing agents, BME and DMSO [
6,
16]. Furthermore, CK is primarily found in the brain as a natural metabolic by product of the transamination of sulfur-containing amino acids [
17,
18]. LK and AECK have been purified from bovine brain and binding and interaction studies have been successfully completed in the presence other brain specific imine reductases [
19]. LK has also been observed to exhibit neuroprotective, neurotrophic and anti-inflammatory activities [
10]. Additionally, the derivative LKEE displayed a higher efficiency for membrane permeability and was revealed to protect motor neurons from oxidative stress in vitro as well as promote neurite outgrowth at nanomolar concentrations [
10,
20]. The AECK molecule has been noted to react with a similar chemistry to LK and LKEE and is thought to play similar physiological and biochemical roles [
21]. The group has also been shown to prevent ischemic neuronal injury via the innate neuroprotective function [
22]. CKs are now noted as novel neurotrophic small molecules that hold some promise for the treatment of neurodegenerative diseases [
23]. The use of these stable, non-toxic and naturally occurring chemical for promoting the differentiation of a heterogeneous mesenchymal stromal/stem cell population toward a neurogenic lineage was tested and the phenotypic characterization by proteomic quantification was completed.
3.1. Neurogenic Related Roles of Identified Proteins
It is now recognized that protein interactions are not random events, their interfaces being precisely coordinated spatially and temporally according to biological and environmental cues [
24]. In stem cell biology, these signals affect protein-protein interactions, which regulate the molecular processes of proliferation and differentiation. The systems biology approach of visualizing interacting proteins within a proteome presents in silico datasets as graphical a network representation at a global proteome scale and has become a significant tool in understanding the biological context in which proteins function and localize to promote complex cellular events.
The action of CKs have been previously shown to have an effect through primary interactions with thiomorpholine-carboxylate dehydrogenase and μ-crystallin proteins [
9,
10]. These initiating factors are likely responsible for potentiating the downstream crosstalk interaction between the MAP3K and Wnt/β-catenin pathways [
25], which are vital in neural development. Complementary to pathway promotions, numerous correlative interacting protein targets and partners have statistical up regulation of and biologically relevant fold changes that support neurogenic differentiation, neurite migration as well as neuron and axon development. Furthermore, the reductive capacity of the chemicals also regulates oxidative stress and anti-inflammatory conditions. Whereby a substantial number of cellular damaging stress related proteins and cytokines, previously observed in BME treatment in Santos et al. [
6], have been minimized in their expression. Therefore, consolidating the CKs gentle inductive capacity for neural differentiation. The investigation of the roles and interacting partners of up-regulated neural related proteins is important to comprehend the biological context of their expression.
Filamin-B was identified in both chemically treated ADSCs with a statistically significant up regulation in each differentiated cell. Filamin-B is an actin-binding protein that is highly expressed in the CNS, displaying roles in cellular migration and differentiation [
26]. The relevance of Filamin-B in neurogenic differentiating ADSCs stems from its annotated role in neuroblast migration in the developing brain from the ventricular zone to the outer cortical plate [
26]. The migratory roles are linked to the interaction between Filamin-B and its 70% homologous interacting partner Filamin-A, which promotes the development and maintenance of the cell’s bipolar shape [
27]. The up regulation of Filamin-B in treated ADSCs is significant since the morphological shape of the treated cells appear elongated and bipolar prior to the development of neurite outgrowth (
Figure 1G,H). An interesting binding partner was also identified within the statistically relevant proteins of both treatments. The partner; Ras-related C3 botulinum toxin substrate 1 (Rac1) is annotated with the biological process of nerve growth factor receptor signaling pathway and is a neural surface antigen. Rac1 is a pleiotropic regulator of a variety of cellular processes including the proliferation, differentiation, and neuronal maturation during embryonic and adult hippocampal development [
28]. The assistance in axonal migration and dendritic development is directed via its signaling with a dense core of primary interacting proteins of the P21 protein (Cdc42/Rac) activated kinase family. This acts as a GTPase effector that links the action to the JNK/MAPK pathway regulating the spatial reorganization of the cytoskeleton for neural development and dendrite morphogenesis [
29]. The identification of Rac1 in neuronal differentiating ADSCs indicates the morphological changes observed are maintained and directed through a number of control mechanisms that are widely seen in the development of in vivo neural tissue. Rac1 has the potential to be utilized as a novel marker for neurogenesis of ADSCs. The identification of Rac1 in neural differentiating ADSCs has the potential to be utilized as a novel marker for neurogenesis. This finding further supports the CK induction of ADSCs toward a neural phenotype.
Thioredoxin and Peroxiredoxin-1 were abundantly expressed in both chemical treated ADSCs. Each protein is found ubiquitously in mammalian cells [
30] thus they would not be suitable as a neurospecific marker. Their function however is of great importance especially within the context of this study’s focus on chemical induction and role of low oxidative stress. All Peroxiredoxins contain a conserved cysteine residue which undergoes a cycle of peroxide-dependent oxidation and thiol-dependent reduction [
31]. This is particularly relevant since it regulates the intracellular redox state, effectively protecting the cells from oxidative stress. This may occur, to a certain extent, based on the oxidative-stress response proteins seen up regulated with BME treatment in a previous paper [
6]. A study by Simzar et al. [
32] found that the overexpression of peroxiredoxin in PC12 neuronal cells in vitro increased the presence of reactive oxygen species, essentially creating an oxidative stress inducing environment, this does not seem to be the case in this study as the cell population is maintained at sufficiently high numbers. The activity of Peroxiredoxin-1 is regulated by thioredoxin, which reduces the cysteine in peroxiredoxin [
33]. The regulatory mechanisms maintaining the levels of peroxiredoxin and thioredoxin levels are essential in preserving cells in distress. A system of controlled oxidative stress has been shown to increase neurogenesis and oligodendrogenesis in adult neural progenitor cells [
34]. The importance of expression and regulation of these two molecules in sustaining cells for proliferation and guiding neurogenesis is essential; furthermore, it can be used as a marker of the level of stress experienced by neurogenic induced cells.
The network dynamics across both AECK and LK treated cells presented a range of shared and highly interactive neuronal-related proteins, and the identification of Neuroblast differentiation-associated protein (AHNAK) as a hub for many interactions supports the conclusion that a differentiation process is occurring. Furthermore, AHNAK binding partners annexin2 and S100-A10 were also identified with at least a two-fold increase over basal levels. The established GO classification for AHNAK is its involvement in nervous system development. It has also been recognized to have multiple roles in neuronal development dependent on its cellular localization and calcium concentrations which regulates actin cytoskeleton organization, cell membrane architecture and cell-cell junction formation [
35,
36,
37,
38,
39]. Moreover, the non-membrane bound AHNAK promiscuously interacts with several reported partners such as protein kinase C and phospholipase C which are involved in the activation of inositol metabolism [
40,
41,
42]. Protein kinase C and phospholipase C were both found to be statistically significantly up regulated in both the AECK and LK treated ADSCs. Inositol metabolism is vital in regulating the Wnt/β-catenin pathway for nerve guidance, serotonin modulation and the control of intracellular calcium concentration [
40,
42]. There is mounting evidence for the usefulness of AHNAK as a marker for neurogenesis especially in the co-expression and up regulation of neuronal development related binding partners. This is further evidence that the CKs are more suitable for inducing neurogenesis of ADSCs.
An associated multifunctional-actin binding protein uniquely identified in the LK treated ADSCs is Gelsolin. Gelsolin localization and expression has been determined to be highest in the CNS and PNS and it has been found to be involved in a number of growth promoting and neuroprotective functions mediated possibly through the Wnt/β-catenin pathway [
43]. Its role in actin remodeling in the nervous system is calcium dependent, with the protein initiating actin polymerization or disassembly. Since the highest expression has been determined to be in the oligodendrocytes and Schwann cells myelin sheaths, Tanaka et al. [
43] proposed the role of gelsolin to be involved in the maturation of the myelin sheath forming cells. The neuroprotective capability is attributed to its anti-oxidative and anti-apoptotic functions in high oxidative-stress induced environments [
44]. Interestingly the down regulation or proteolytic cleavage of gelsolin has been linked to the development of Alzheimer’s disease [
45]. The up regulation of Gelsolin in the CK treated ADSCs and its value as a neuronal marker is apparent, since the presence of a neuronally related cytoskeletal remodelling protein with neuroprotective capabilities is an appealing find in the array of up regulated proteins of neurogenic differentiating ADSCs. This is further evidence that the CK chemicals are better suited for inducing neurogenesis in ADSCs.
Spectrin alpha and beta chain has been widely studied and are known to be major cytoskeletal components in the brain and distributed in the cytoplasm of neural cells and is modularly downstream regulated by the Wnt signaling pathway [
46]. There is now evidence that spectrin regulates the surface chemistry and morphology of neuronal cells and large modifications or degradation would produce major modifications to synapses [
46]. Spectrin has also been implicated in the calcium regulated release of neurotransmitters between developing synapses [
47]. The regulation of spectrin is derived from the calcium modulated calpain proteolytic enzyme [
46]. The levels of calpain in both chemical treatments in this study are negligible. Here the presence of a heterodimer of the brain isoforms of spectrin alpha and beta chain is well noted in the prospect of utility as a neural marker for further studies.
Lastly Galectin-3 increase in expression has been detected in the LK treated ADSCs. Recent studies have shown that galectin-3 is expressed in a variety of neuronal tissues, especially glial cells in the CNS in which it directs oligodendrocyte differentiation to control myelin sheath formation [
48]. Furthermore, the control of neuroblast migration in brain development was proposed by Comte et al. [
49]. A function that is more suited to the development of neuronal like cells was found by Pesheva et al., in which galectin-3 was found to stimulate neural cell adhesion and moreover, neurite outgrowth in developing cells [
50]. This also provides further evidence that the ADSCs are responding favorably to the CK treatment, expressing neuronal-related proteins known to be functionally and structurally useful.
3.2. Neurogenic Roles of Cytokines
Cytokines are pleotropic proteins that coordinate signaling across varied tissues and cell types including during neural development [
51]. Cytokines display functional roles in a variety of developmental stages, acting as neurotrophic factors initiating the repair and regeneration of cells [
51]. Furthermore, certain groups of cytokines, such as chemokines, regulate the directed growth and communication between radially migrating neuronal cells which give rise to mature neurons, glial, astrocytes and oligodendrocytes [
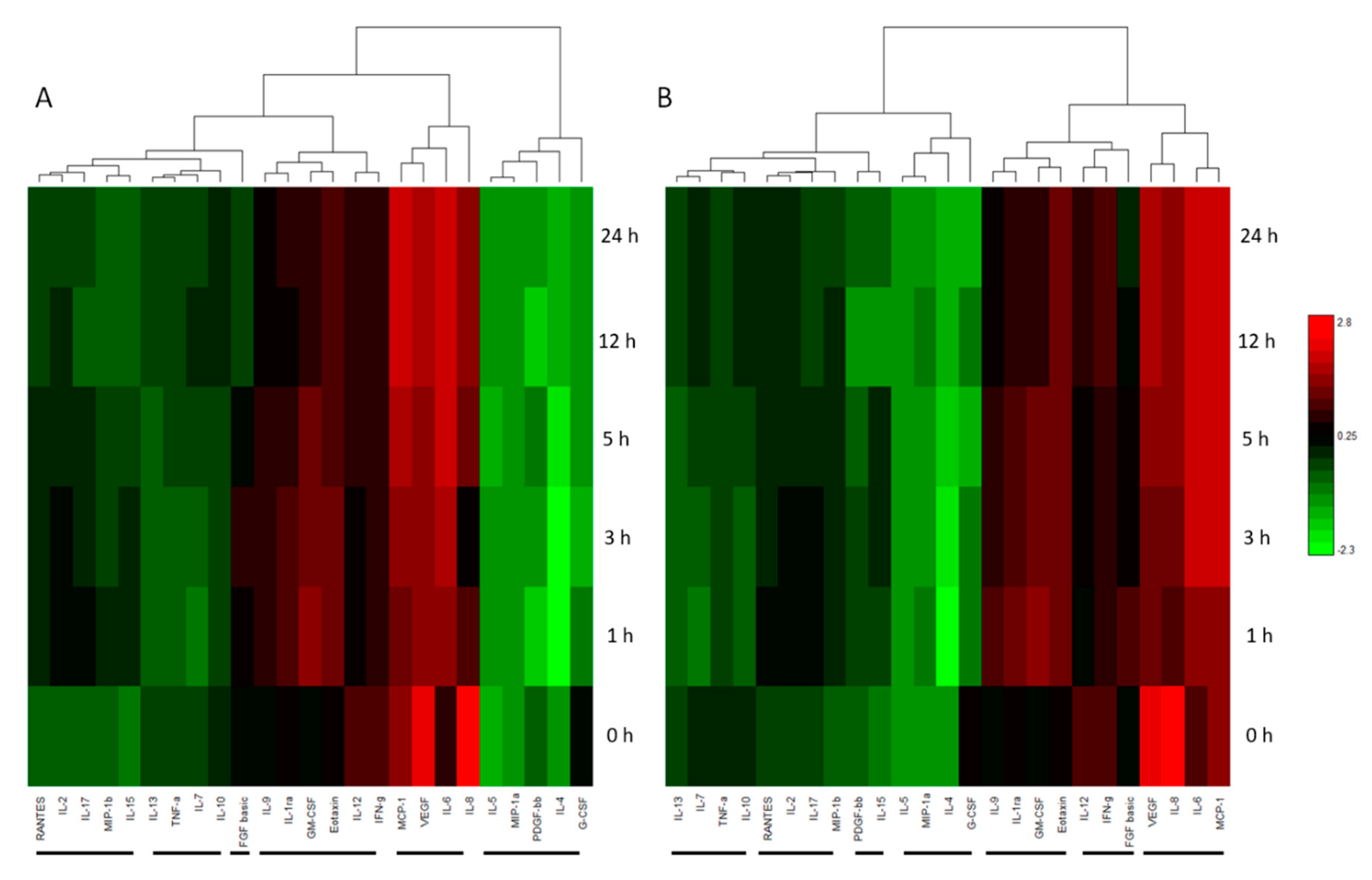
51]. Due to their relatively low abundance and the dynamic range of a proteome, these molecules are extremely difficult to detect by MS and alternative detection methods, such as the Bioplex cytokine, chemokine, and growth factor assay system, allows the relative quantitation and comparison of 27 secreted cytokines. Hierarchical group clustering was carried out to identify the cytokines that responded with similar trends during the differentiation process and possible roles within the CK treated cells relative to neurogenesis.
The cytokine expression changes in the CK treated ADSCs closely support the proteomic data displaying a similar broad change in concentrations and the thus cellular phenotype in response to treatment. The intimate view of the cytokines is vital in understanding their corresponding roles in neural development and growth. The concentration change of molecules from group 1 (IL-1ra, IL-2, MIP-1b, RANTES and MIP-1a) was apparent with the out of trend large fluctuation of MIP-1b and RANTES in the LK treated ADSCs (See
supplementary material for detailed grouping). MIP-1b is a pro-inflammatory protein which displayed a substantial increase at 20 h in LK, indicating a significant pro-inflammatory affect. Interestingly, the increase in MIP-1b is mirrored by RANTES with a proceeding concentration increase at the same time point. The secretion of MIP-1b has been noted in a number of neuronal cells [
52]. The developmental organization of neuronal cells and CNS development have been linked to a synchronous regulation of MIP-1b alongside other chemokines [
53]. Studies have shown that MIP-1b knockout mice soon die after birth due to brain formation abnormalities [
53]. Thus, the role of MIP-1b in ADSCs has gained some interest especially with a number of co-regulations at the same time point in other cytokine groups. RANTES, also a pro-inflammatory cytokine, was found to initiate neuro-protective roles assisting in the survival of stressed hippocampal cell lines in the presence of a toxin [
54]. Interestingly a study conducted into investigating the effect of certain pain management drugs in patients with HIV found that morphine treatment deregulated and decreased the expression of RANTES in the neuronal tissue. This progressive effect was implicated in the decrease in microglial cell migration due to the suppression of the chemotactic cytokine [
55]. Furthermore, another study into the effect of HIV glycoprotein120 on neuronal tissue presented a number of up regulated secreted pro-inflammatory cytokines including RANTES which offered a neuroprotective role [
56]. These responses all lead to the stimulation of the CX3CL1, CCL4 and CCL5 receptors of the above cytokines [
53] which may also be regulated in the differentiating ADSCs.
Examining beyond group 1 with the consideration of the 20 h time point, a number of similar variations in trends were identified amongst several cytokines in other groups. The decrease of IL-15 is a significant find as its expression and regulation, through an indirect neuroprotective mechanism, in astroglial cells has been found to be linked to RANTES, MIP-1a, MIP-1b and GM-CSF [
57]. Interestingly IL-15, IL-17 and GM-CSF in LK’s group 5 present a massive decrease in concentration per cell from the 5 to 20 h time point. Elevated concentrations of IL-15 in neural stem cell (NSC) cultures have proven to reduce maturation and neurite out growth in differentiating neurons but not affect proliferation [
58]. Furthermore, IL-15-deficient mice exhibited defective JAK/STAT and ERK pathways which are key in the regulation of differentiation in NSCs. This substantiates the possible role for the decrease of IL-15 in LK treated ADSCs which presented greater morphological differentiation than the AECK treated cells. The down regulation of IL-17 is also considered a beneficial decrease for cytokines in brain tissue, since elevated levels are usually present in a number of traumatic brain injuries or infarcts in which a large proportion of cells are damaged [
59].
The cytokine (Bioplex) data complement the proteomic findings that the AECK and LK treatments of ADSCs initiate the differentiation toward a neural lineage and potentially neurons, evidenced by the proteomic and microscopic analyses. The results also indicate the ADSCs favor the treatment with LK over AECK for neuronal-like differentiation within the 24 h treatment time. Notwithstanding the AECK did produce a slightly higher cell population at the end of the differentiation time and similar neurogenic-related proteins were identified. The cytokine data also are consistent with a lack of cytotoxic stress of the cells over the period examined, thus removing cytotoxicity as a cause for the neuronal-like morphological changes [
7].










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}