The Role of Transient Receptor Potential (TRP) Channels in the Transduction of Dental Pain

 ,
,  , and
, and 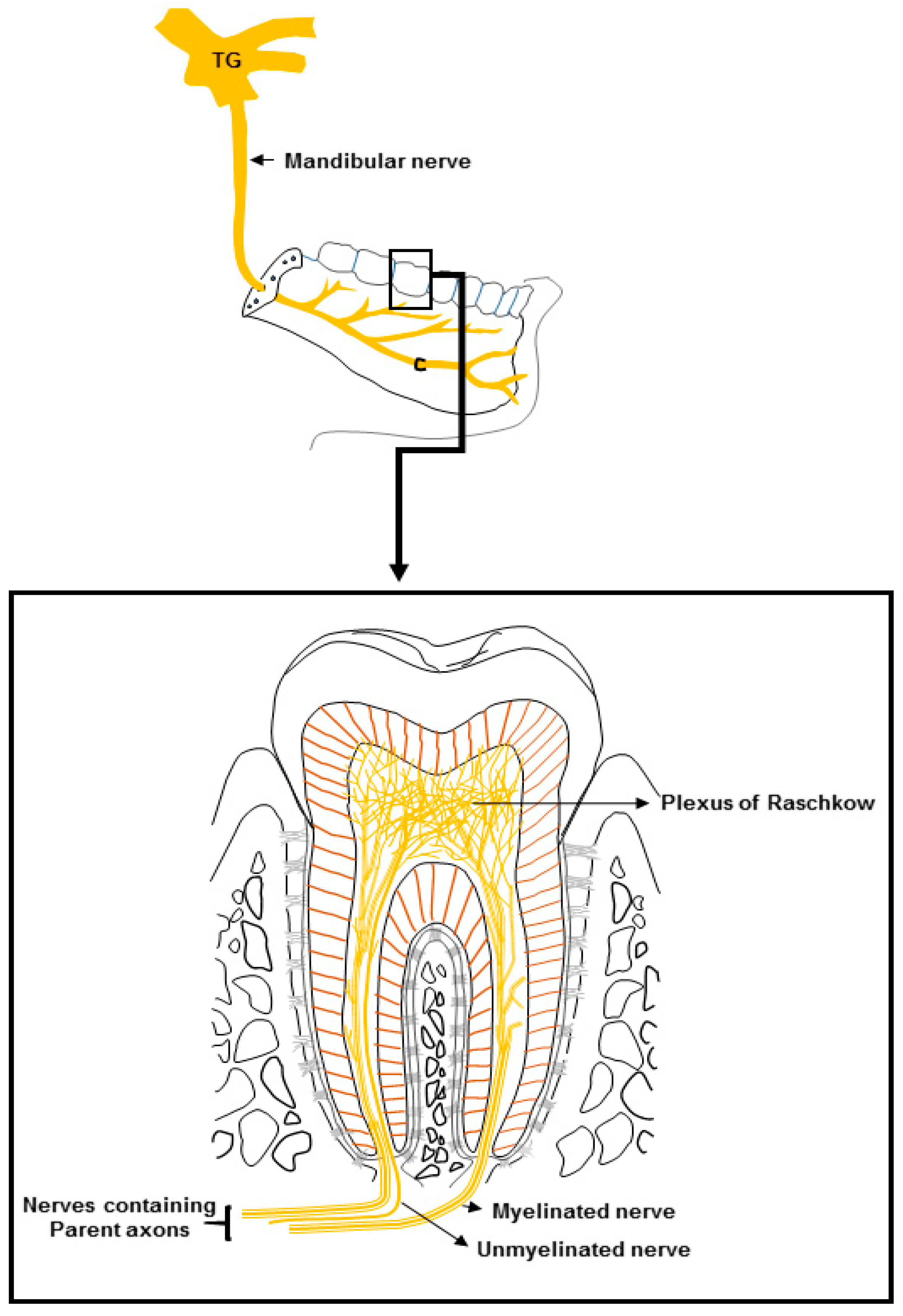{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Dental Innervation
3. TRP Channels and Their Presence in Dental Tissues
3.1. TRPV
3.1.1. TRPV in DPAs
3.1.2. TRPV in Odontoblasts
3.2. TRPM
3.2.1. TRPM in DPAs
3.2.2. TRPM in Odontoblasts
3.3. TRPA
3.3.1. TRPA in DPAs
3.3.2. TRPA in Odontoblasts
3.4. TRPC
TRPC in Odontoblasts
3.5. TRP Channels in Pulpal Fibroblasts and Blood Vessels
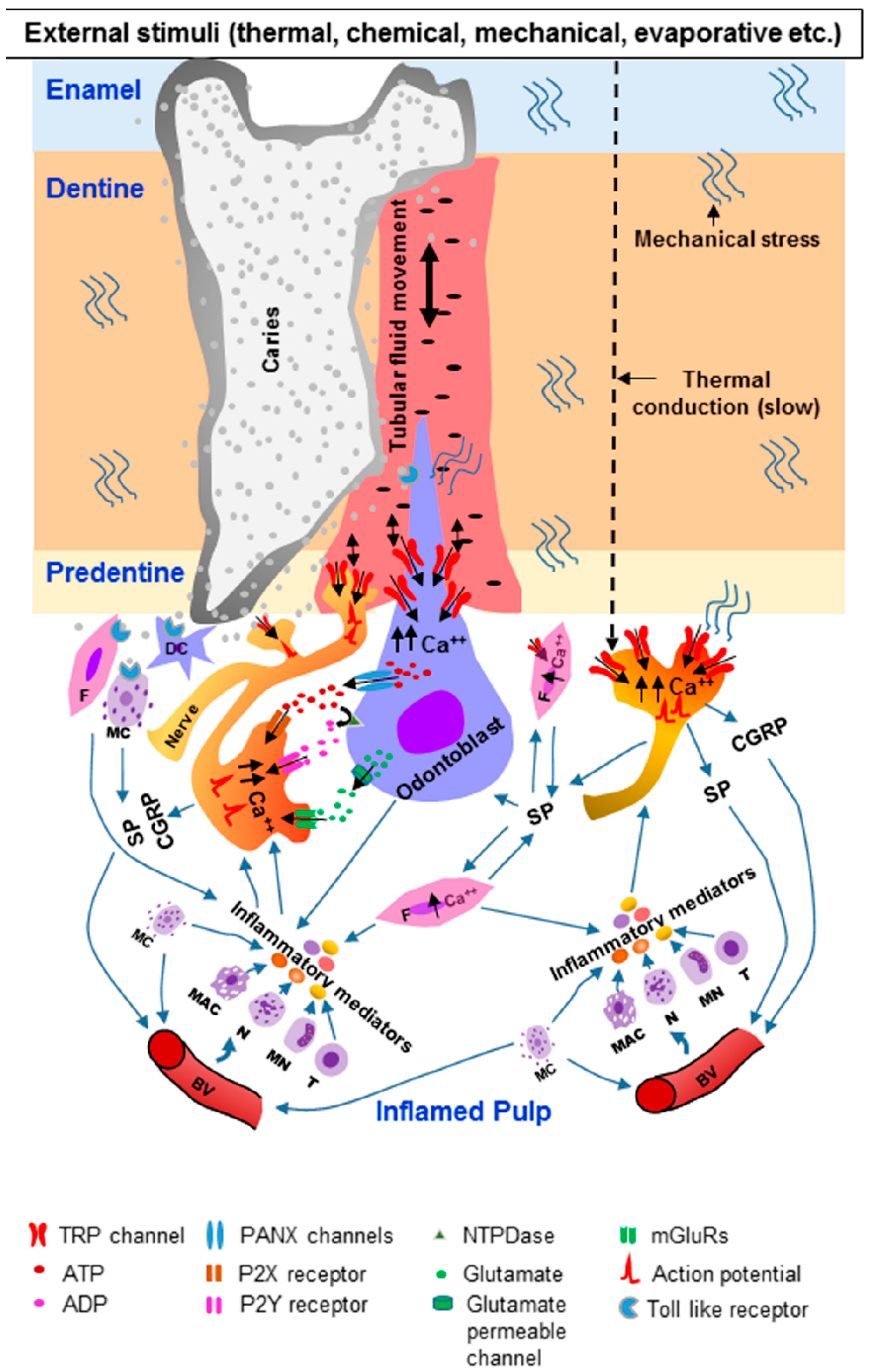
4. Involvement of TRP Channels in the Transduction of Dental Pain
5. Clinical Significance
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kassebaum, N.J.; Bernabe, E.; Dahiya, M.; Bhandari, B.; Murray, C.J.; Marcenes, W. Global burden of untreated caries: A systematic review and metaregression. J. Dent. Res. 2015, 94, 650–658. [Google Scholar] [CrossRef]
- Hahn, C.L.; Falkler, W.A., Jr.; Minah, G.E. Microbiological studies of carious dentine from human teeth with irreversible pulpitis. Arch. Oral Biol. 1991, 36, 147–153. [Google Scholar] [CrossRef]
- Bergenholtz, G. Inflammatory response of the dental pulp to bacterial irritation. J. Endod. 1981, 7, 100–104. [Google Scholar] [CrossRef]
- Dummer, P.M.; Hicks, R.; Huws, D. Clinical signs and symptoms in pulp disease. Int. Endod. J. 1980, 13, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.R.; Holder, M.J.; Smith, A.J. Inflammation and regeneration in the dentin-pulp complex: A double-edged sword. J. Endod. 2014, 40, S46–S51. [Google Scholar] [CrossRef]
- Mitchell, D.A.; Mitchell, L. Oxford Handbook of Clinical Dentistry, 4th ed.; Oxford University Press: Oxford, UK, 2005; p. 260. [Google Scholar]
- McCarthy, P.J.; McClanahan, S.; Hodges, J.; Bowles, W.R. Frequency of localization of the painful tooth by patients presenting for an endodontic emergency. J. Endod. 2010, 36, 801–805. [Google Scholar] [CrossRef]
- Splieth, C.H.; Tachou, A. Epidemiology of dentin hypersensitivity. Clin. Oral Investig. 2013, 17, S3–S8. [Google Scholar] [CrossRef]
- Flynn, J.; Galloway, R.; Orchardson, R. The incidence of ‘hypersensitive’ teeth in the West of Scotland. J. Den. 1985, 13, 230–236. [Google Scholar] [CrossRef]
- Fischer, C.; Fischer, R.G.; Wennberg, A. Prevalence and distribution of cervical dentine hypersensitivity in a population in Rio de Janeiro, Brazil. J. Den. 1992, 20, 272–276. [Google Scholar] [CrossRef]
- Irwin, C.R.; McCusker, P. Prevalence of dentine hypersensitivity in a general dental population. J. Ir. Dent. Assoc. 1997, 43, 7–9. [Google Scholar]
- Liu, H.C.; Lan, W.H.; Hsieh, C.C. Prevalence and distribution of cervical dentin hypersensitivity in a population in Taipei, Taiwan. J. Endod. 1998, 24, 45–47. [Google Scholar] [CrossRef]
- Dowell, P.; Addy, M.; Dummer, P. Dentine hypersensitivity: Aetiology, differential diagnosis and management. Br. Dent. J. 1985, 158, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Dababneh, R.H.; Khouri, A.T.; Addy, M. Dentine hypersensitivity—An enigma? A review of terminology, mechanisms, aetiology and management. Br. Dent. J. 1999, 187, 606–611. [Google Scholar] [PubMed]
- Sole-Magdalena, A.; Martinez-Alonso, M.; Coronado, C.A.; Junquera, L.M.; Cobo, J.; Vega, J.A. Molecular basis of dental sensitivity: The odontoblasts are multisensory cells and express multifunctional ion channels. Ann. Anat. 2018, 215, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Rahim, Z.H.; Bakri, M.M.; Hm, Z.; Ia, A.; Na, Z. High fluoride and low pH level have been detected in popular flavoured beverages in Malaysia. Pak. J. Med. Sci. 2014, 30, 404–408. [Google Scholar] [CrossRef]
- Bakri, M.M.; Hossain, M.Z.; Razak, F.A.; Saqina, Z.H.; Misroni, A.A.; Ab-Murat, N.; Kitagawa, J.; Saub, R.B. Dentinal tubules occluded by bioactive glass-containing toothpaste exhibit high resistance toward acidic soft drink challenge. Aust. Dent. J. 2017, 62, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Brannstrom, M.; Linden, L.A.; Astrom, A. The hydrodynamics of the dental tubule and of pulp fluid. A discussion of its significance in relation to dentinal sensitivity. Caries Res. 1967, 1, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Brannstrom, M.; Astrom, A. The hydrodynamics of the dentine; its possible relationship to dentinal pain. Int. Dent. J. 1972, 22, 219–227. [Google Scholar] [PubMed]
- Sessle, B.J. The neurobiology of facial and dental pain: Present knowledge, future directions. J. Dent. Res. 1987, 66, 962–981. [Google Scholar] [CrossRef]
- Linsuwanont, P.; Versluis, A.; Palamara, J.E.; Messer, H.H. Thermal stimulation causes tooth deformation: A possible alternative to the hydrodynamic theory? Arch. Oral Biol. 2008, 53, 261–272. [Google Scholar] [CrossRef]
- Jacobs, H.R.; Thompson, R.E.; Brown, W.S. Heat transfer in teeth. J. Dent. Res. 1973, 52, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, B.A.; McGinley, M.B.; Brown, W.S. Thermal stress in teeth. J. Dent. Res. 1978, 57, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, H.; Matthews, B. In-vitro observations on fluid flow through human dentine caused by pain-producing stimuli. Arch. Oral Biol. 1973, 18, 275–294. [Google Scholar] [CrossRef]
- Linsuwanont, P.; Palamara, J.E.; Messer, H.H. An investigation of thermal stimulation in intact teeth. Arch. Oral Biol. 2007, 52, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Chidchuangchai, W.; Vongsavan, N.; Matthews, B. Sensory transduction mechanisms responsible for pain caused by cold stimulation of dentine in man. Arch. Oral Biol. 2007, 52, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Shibukawa, Y.; Sato, M.; Kimura, M.; Sobhan, U.; Shimada, M.; Nishiyama, A.; Kawaguchi, A.; Soya, M.; Kuroda, H.; Katakura, A.; et al. Odontoblasts as sensory receptors: Transient receptor potential channels, pannexin-1, and ionotropic ATP receptors mediate intercellular odontoblast-neuron signal transduction. Pflug. Arch. 2015, 467, 843–863. [Google Scholar] [CrossRef] [PubMed]
- Trowbridge, H.O.; Franks, M.; Korostoff, E.; Emling, R. Sensory response to thermal stimulation in human teeth. J. Endod. 1980, 6, 405–412. [Google Scholar] [CrossRef]
- Fuss, Z.; Trowbridge, H.; Bender, I.B.; Rickoff, B.; Sorin, S. Assessment of reliability of electrical and thermal pulp testing agents. J. Endod. 1986, 12, 301–305. [Google Scholar] [CrossRef]
- Närhi, M.; Ngassapa, D.; Shimono, M.M.T.; Suda, H.; Takahashi, K. Function of intradental nociceptors in normal and inflamed teeth. In Dentin/pulp Complex; Shimono, M., Maeda, T., Suda, H., Takahashi, K., Eds.; Quintessence: Tokyo, Japan, 1996; pp. 136–140. [Google Scholar]
- Jyvasjarvi, E.; Kniffki, K.D. Cold stimulation of teeth: A comparison between the responses of cat intradental A delta and C fibres and human sensation. J. Physiol. 1987, 391, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Magloire, H.; Maurin, J.C.; Couble, M.L.; Shibukawa, Y.; Tsumura, M.; Thivichon-Prince, B.; Bleicher, F. Topical review. Dental pain and odontoblasts: Facts and hypotheses. J. Orofac. Pain 2010, 24, 335–349. [Google Scholar]
- Allard, B.; Magloire, H.; Couble, M.L.; Maurin, J.C.; Bleicher, F. Voltage-gated sodium channels confer excitability to human odontoblasts: Possible role in tooth pain transmission. J. Biol. Chem. 2006, 281, 29002–29010. [Google Scholar] [CrossRef] [PubMed]
- Hermanstyne, T.O.; Markowitz, K.; Fan, L.; Gold, M.S. Mechanotransducers in rat pulpal afferents. J. Dent. Res. 2008, 87, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Chung, G.; Jung, S.J.; Oh, S.B. Cellular and molecular mechanisms of dental nociception. J. Dent. Res. 2013, 92, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Chung, G.; Oh, S.B. TRP channels in dental pain. Open Pain J. 2013, 6, 31–36. [Google Scholar] [CrossRef]
- Narhi, M. The neurophysiology of the teeth. Dent. Clin. N. Am. 1990, 34, 439–448. [Google Scholar]
- Byers, M.R.; Narhi, M.V. Dental injury models: Experimental tools for understanding neuroinflammatory interactions and polymodal nociceptor functions. Crit. Rev. Oral Biol. Med. 1999, 10, 4–39. [Google Scholar] [CrossRef] [PubMed]
- Fried, K.; Sessle, B.J.; Devor, M. The paradox of pain from tooth pulp: Low-threshold “algoneurons”? Pain 2011, 152, 2685–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, P.N. Neural elements in dental pulp and dentin. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 1995, 80, 710–719. [Google Scholar] [Green Version]
- Johnsen, D.; Johns, S. Quantitation of nerve fibres in the primary and permanent canine and incisor teeth in man. Arch. Oral Biol. 1978, 23, 825–829. [Google Scholar] [CrossRef]
- Paik, S.K.; Park, K.P.; Lee, S.K.; Ma, S.K.; Cho, Y.S.; Kim, Y.K.; Rhyu, I.J.; Ahn, D.K.; Yoshida, A.; Bae, Y.C. Light and electron microscopic analysis of the somata and parent axons innervating the rat upper molar and lower incisor pulp. Neuroscience 2009, 162, 1279–1286. [Google Scholar] [CrossRef]
- Fried, K.; Arvidsson, J.; Robertson, B.; Brodin, E.; Theodorsson, E. Combined retrograde tracing and enzyme/immunohistochemistry of trigeminal ganglion cell bodies innervating tooth pulps in the rat. Neuroscience 1989, 33, 101–109. [Google Scholar] [CrossRef]
- Fried, K.; Hildebrand, C. Axon number and size distribution in the developing feline inferior alveolar nerve. J. Neurol. Sci. 1982, 53, 169–180. [Google Scholar] [CrossRef]
- Johansson, C.S.; Hildebrand, C.; Povlsen, B. Anatomy and developmental chronology of the rat inferior alveolar nerve. Anat. Rec. 1992, 234, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Paik, S.K.; Lee, D.S.; Kim, J.Y.; Bae, J.Y.; Cho, Y.S.; Ahn, D.K.; Yoshida, A.; Bae, Y.C. Quantitative ultrastructural analysis of the neurofilament 200-positive axons in the rat dental pulp. J. Endod. 2010, 36, 1638–1642. [Google Scholar] [CrossRef] [PubMed]
- Cadden, S.W.; Lisney, S.J.; Matthews, B. Thresholds to electrical stimulation of nerves in cat canine tooth-pulp with A beta-, A delta- and C-fibre conduction velocities. Brain Res. 1983, 261, 31–41. [Google Scholar] [CrossRef]
- Lisney, S.J. Some anatomical and electrophysiological properties of tooth-pulp afferents in the cat. J. Physiol. 1978, 284, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.A.; Luo, S.; Levinson, S.R. Unmyelinated nerve fibers in the human dental pulp express markers for myelinated fibers and show sodium channel accumulations. BMC Neurosci. 2012, 13, 29. [Google Scholar] [CrossRef]
- Carda, C.; Peydro, A. Ultrastructural patterns of human dentinal tubules, odontoblasts processes and nerve fibres. Tissue Cell 2006, 38, 141–150. [Google Scholar] [CrossRef]
- Byers, M.R. Terminal arborization of individual sensory axons in dentin and pulp of rat molars. Brain Res. 1985, 345, 181–185. [Google Scholar] [CrossRef]
- Aars, H.; Brodin, P.; Andersen, E. A study of cholinergic and β-adrenergic components in the regulation of blood flow in the tooth pulp and gingiva in man. Acta Physiol. Scand. 1993, 148, 441–447. [Google Scholar] [CrossRef]
- Sasano, T.; Shoji, N.; Kuriwada, S.; Sanjo, D.; Izumi, H.; Karita, K. Absence of parasympathetic vasodilatation in cat dental pulp. J. Dent. Res. 1995, 74, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Carr, P.A.; Nagy, J.I. Emerging relationships between cytochemical properties and sensory modality transmission in primary sensory neurons. Brain Res. Bull. 1993, 30, 209–219. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, F.J.; Morris, H.R.; Priestley, J.V. Sub-populations of smaller diameter trigeminal primary afferent neurons defined by expression of calcitonin gene-related peptide and the cell surface oligosaccharide recognized by monoclonal antibody LA4. J. Neurocytol. 1991, 20, 716–731. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.D.; Kruger, L. Lectin and neuropeptide labeling of separate populations of dorsal root ganglion neurons and associated “nociceptor” thin axons in rat testis and cornea whole-mount preparations. Somatosens. Res. 1988, 5, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, E.J.; Burnstock, G.; McMahon, S.B. The expression of P2X3 purinoreceptors in sensory neurons: Effects of axotomy and glial-derived neurotrophic factor. Mol. Cell. Neurosci. 1998, 12, 256–268. [Google Scholar] [CrossRef]
- Chen, C.C.; Akopian, A.N.; Sivilotti, L.; Colquhoun, D.; Burnstock, G.; Wood, J.N. A P2X purinoceptor expressed by a subset of sensory neurons. Nature 1995, 377, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Vulchanova, L.; Riedl, M.S.; Shuster, S.J.; Stone, L.S.; Hargreaves, K.M.; Buell, G.; Surprenant, A.; North, R.A.; Elde, R. P2X3 is expressed by DRG neurons that terminate in inner lamina II. Eur. Neurosci. 1998, 10, 3470–3478. [Google Scholar] [CrossRef]
- Ichikawa, H.; Sugimoto, T. The co-expression of ASIC3 with calcitonin gene-related peptide and parvalbumin in the rat trigeminal ganglion. Brain Res. 2002, 943, 287–291. [Google Scholar] [CrossRef]
- Ichikawa, H.; Deguchi, T.; Nakago, T.; Jacobowitz, D.M.; Sugimoto, T. Parvalbumin- and calretinin-immunoreactive trigeminal neurons innervating the rat molar tooth pulp. Brain Res. 1995, 679, 205–211. [Google Scholar] [CrossRef]
- Yang, H.; Bernanke, J.M.; Naftel, J.P. Immunocytochemical evidence that most sensory neurons of the rat molar pulp express receptors for both glial cell line-derived neurotrophic factor and nerve growth factor. Arch. Oral Biol. 2006, 51, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Fried, K.; Risling, M. Nerve growth factor receptor-like immunoreactivity in primary and permanent canine tooth pulps of the cat. Cell Tissue Res. 1991, 264, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Naftel, J.P.; Wheeler, E.F. Effects of deprivation of neonatal nerve growth factor on the expression of neurotrophin receptors and brain-derived neurotrophic factor by dental pulp afferents of the adult rat. Arch. Oral Biol. 2000, 45, 387–399. [Google Scholar] [CrossRef]
- Ichikawa, H.; Deguchi, T.; Fujiyoshi, Y.; Nakago, T.; Jacobowitz, D.M.; Sugimoto, T. Calbindin-D28k-immunoreactivity in the trigeminal ganglion neurons and molar tooth pulp of the rat. Brain Res. 1996, 715, 71–78. [Google Scholar] [CrossRef]
- Ichikawa, H.; Fukuda, T.; Terayama, R.; Yamaai, T.; Kuboki, T.; Sugimoto, T. Immunohistochemical localization of gamma and β subunits of epithelial Na+ channel in the rat molar tooth pulp. Brain Res. 2005, 1065, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Ishida-Yamamoto, A.; Senba, E.; Ueda, Y.; Tohyama, M. Calcitonin gene-related peptide containing sensory neurons innervating tooth pulp and buccal mucosa of the rat: An immunohistochemical analysis. J. Chem. Neuroanat. 1990, 3, 155–163. [Google Scholar] [PubMed]
- Jacobsen, E.B.; Fristad, I.; Heyeraas, K.J. Nerve fibers immunoreactive to calcitonin gene-related peptide, substance P, neuropeptide Y, and dopamine β-hydroxylase in innervated and denervated oral tissues in ferrets. Acta Odontol. Scand. 1998, 56, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damann, N.; Voets, T.; Nilius, B. TRPs in our senses. Curr. Biol. 2008, 18, R880–889. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Szallasi, A. Transient receptor potential channels as drug targets: From the science of basic research to the art of medicine. Pharmacol. Rev. 2014, 66, 676–814. [Google Scholar] [CrossRef] [PubMed]
- Jardin, I.; Lopez, J.J.; Diez, R.; Sanchez-Collado, J.; Cantonero, C.; Albarran, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front. Physiol. 2017, 8, 392. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Jones, K.; Hafen, E.; Rubin, G. Rescue of the Drosophila phototransduction mutation trp by germline transformation. Science 1985, 230, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Rubin, G.M. Molecular characterization of the Drosophila trp locus: A putative integral membrane protein required for phototransduction. Neuron 1989, 2, 1313–1323. [Google Scholar] [CrossRef]
- Minke, B.; Selinger, Z. The roles of trp and calcium in regulating photoreceptor function in Drosophila. Curr. Opin. Neurobiol. 1996, 6, 459–466. [Google Scholar] [CrossRef]
- Li, H. TRP Channel Classification. Adv. Exp. Med. Biol. 2017, 976, 1–8. [Google Scholar] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [PubMed]
- Vandewauw, I.; Owsianik, G.; Voets, T. Systematic and quantitative mRNA expression analysis of TRP channel genes at the single trigeminal and dorsal root ganglion level in mouse. BMC Neurosci. 2013, 14, 21. [Google Scholar] [CrossRef]
- Flegel, C.; Schobel, N.; Altmuller, J.; Becker, C.; Tannapfel, A.; Hatt, H.; Gisselmann, G. RNA-Seq Analysis of Human Trigeminal and Dorsal Root Ganglia with a Focus on Chemoreceptors. PLoS ONE 2015, 10, e0128951. [Google Scholar] [CrossRef]
- Vay, L.; Gu, C.; McNaughton, P.A. The thermo-TRP ion channel family: Properties and therapeutic implications. Br. J. Pharm. 2012, 165, 787–801. [Google Scholar] [CrossRef]
- Voets, T.; Talavera, K.; Owsianik, G.; Nilius, B. Sensing with TRP channels. Nat. Chem. Biol. 2005, 1, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Voets, T.; Droogmans, G.; Wissenbach, U.; Janssens, A.; Flockerzi, V.; Nilius, B. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature 2004, 430, 748–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vriens, J.; Nilius, B.; Voets, T. Peripheral thermosensation in mammals. Nat. Rev. 2014, 15, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef]
- Jordt, S.E.; Tominaga, M.; Julius, D. Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc. Natl. Acad. Sci. USA 2000, 97, 8134–8139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. 2007, 6, 357–372. [Google Scholar]
- Hwang, S.W.; Cho, H.; Kwak, J.; Lee, S.Y.; Kang, C.J.; Jung, J.; Cho, S.; Min, K.H.; Suh, Y.G.; Kim, D.; et al. Direct activation of capsaicin receptors by products of lipoxygenases: Endogenous capsaicin-like substances. Proc. Natl. Acad. Sci. USA 2000, 97, 6155–6160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriyama, T.; Higashi, T.; Togashi, K.; Iida, T.; Segi, E.; Sugimoto, Y.; Tominaga, T.; Narumiya, S.; Tominaga, M. Sensitization of TRPV1 by EP1 and IP reveals peripheral nociceptive mechanism of prostaglandins. Mol. Pain 2005, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Wada, M.; Masu, M. Potentiation of capsaicin receptor activity by metabotropic ATP receptors as a possible mechanism for ATP-evoked pain and hyperalgesia. Proc. Natl. Acad. Sci. USA 2001, 98, 6951–6956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julius, D. TRP channels and pain. Ann. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.Z.; Ando, H.; Unno, S.; Masuda, Y.; Kitagawa, J. Activation of TRPV1 and TRPM8 Channels in the Larynx and Associated Laryngopharyngeal Regions Facilitates the Swallowing Reflex. Int. J. Mol. Sci. 2018, 19, 4113. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef] [PubMed]
- White, J.P.; Urban, L.; Nagy, I. TRPV1 function in health and disease. Curr. Pharm. Biotechnol. 2011, 12, 130–144. [Google Scholar] [CrossRef] [PubMed]
- Christie, S.; Wittert, G.A.; Li, H.; Page, A.J. Involvement of TRPV1 Channels in Energy Homeostasis. Front. Endocrinol. 2018, 9, 420. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Rosen, T.A.; Tominaga, M.; Brake, A.J.; Julius, D. A capsaicin-receptor homologue with a high threshold for noxious heat. Nature 1999, 398, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, W. Transient receptor potential vanilloid channels functioning in transduction of osmotic stimuli. J. Endocrinol. 2006, 191, 515–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahluwalia, J.; Rang, H.; Nagy, I. The putative role of vanilloid receptor-like protein-1 in mediating high threshold noxious heat-sensitivity in rat cultured primary sensory neurons. Eur. Neurosci. 2002, 16, 1483–1489. [Google Scholar] [CrossRef]
- Story, G.M. The emerging role of TRP channels in mechanisms of temperature and pain sensation. Curr. Neuropharmacol. 2006, 4, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Park, U.; Vastani, N.; Guan, Y.; Raja, S.N.; Koltzenburg, M.; Caterina, M.J. TRP vanilloid 2 knock-out mice are susceptible to perinatal lethality but display normal thermal and mechanical nociception. J. Neurosci. 2011, 31, 11425–11436. [Google Scholar] [CrossRef]
- Nedungadi, T.P.; Dutta, M.; Bathina, C.S.; Caterina, M.J.; Cunningham, J.T. Expression and distribution of TRPV2 in rat brain. Exp. Neurol. 2012, 237, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Neeper, M.P.; Liu, Y.; Hutchinson, T.L.; Lubin, M.L.; Flores, C.M. TRPV2 is activated by cannabidiol and mediates CGRP release in cultured rat dorsal root ganglion neurons. J. Neurosci. 2008, 28, 6231–6238. [Google Scholar] [CrossRef]
- Ichikawa, H.; Sugimoto, T. Vanilloid receptor 1-like receptor-immunoreactive primary sensory neurons in the rat trigeminal nervous system. Neuroscience 2000, 101, 719–725. [Google Scholar] [CrossRef]
- Gibbs, J.L.; Melnyk, J.L.; Basbaum, A.I. Differential TRPV1 and TRPV2 channel expression in dental pulp. J. Dent. Res. 2011, 90, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ramsey, I.S.; Kotecha, S.A.; Moran, M.M.; Chong, J.A.; Lawson, D.; Ge, P.; Lilly, J.; Silos-Santiago, I.; Xie, Y.; et al. TRPV3 is a calcium-permeable temperature-sensitive cation channel. Nature 2002, 418, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D.; Gunthorpe, M.J.; Kelsell, R.E.; Hayes, P.D.; Reilly, P.; Facer, P.; Wright, J.E.; Jerman, J.C.; Walhin, J.P.; Ooi, L.; et al. TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 2002, 418, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Vogt-Eisele, A.K.; Weber, K.; Sherkheli, M.A.; Vielhaber, G.; Panten, J.; Gisselmann, G.; Hatt, H. Monoterpenoid agonists of TRPV3. Br. J. Pharm. 2007, 151, 530–540. [Google Scholar] [CrossRef]
- Xu, H.; Delling, M.; Jun, J.C.; Clapham, D.E. Oregano, thyme and clove-derived flavors and skin sensitizers activate specific TRP channels. Nat. Neurosci. 2006, 9, 628–635. [Google Scholar] [CrossRef]
- Peier, A.M.; Reeve, A.J.; Andersson, D.A.; Moqrich, A.; Earley, T.J.; Hergarden, A.C.; Story, G.M.; Colley, S.; Hogenesch, J.B.; McIntyre, P.; et al. A heat-sensitive TRP channel expressed in keratinocytes. Science 2046–2049. [CrossRef]
- Mandadi, S.; Sokabe, T.; Shibasaki, K.; Katanosaka, K.; Mizuno, A.; Moqrich, A.; Patapoutian, A.; Fukumi-Tominaga, T.; Mizumura, K.; Tominaga, M. TRPV3 in keratinocytes transmits temperature information to sensory neurons via ATP. Pflug. Arch. 2009, 458, 1093–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, T.; Ueda, T.; Ugawa, S.; Ishida, Y.; Imayasu, M.; Koyama, S.; Shimada, S. Functional expression of transient receptor potential vanilloid 3 (TRPV3) in corneal epithelial cells: Involvement in thermosensation and wound healing. Exp. Eye Res. 2010, 90, 121–129. [Google Scholar] [CrossRef]
- Aijima, R.; Wang, B.; Takao, T.; Mihara, H.; Kashio, M.; Ohsaki, Y.; Zhang, J.Q.; Mizuno, A.; Suzuki, M.; Yamashita, Y.; et al. The thermosensitive TRPV3 channel contributes to rapid wound healing in oral epithelia. FASEB J. 2015, 29, 182–192. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Lee, H.; Chung, M.K.; Park, U.; Yu, Y.Y.; Bradshaw, H.B.; Coulombe, P.A.; Walker, J.M.; Caterina, M.J. Overexpressed transient receptor potential vanilloid 3 ion channels in skin keratinocytes modulate pain sensitivity via prostaglandin E2. J. Neurosci. 2008, 28, 13727–13737. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, W.; Choe, Y.; Marti-Renom, M.A.; Bell, A.M.; Denis, C.S.; Sali, A.; Hudspeth, A.J.; Friedman, J.M.; Heller, S. Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell 2000, 103, 525–535. [Google Scholar] [CrossRef]
- Strotmann, R.; Harteneck, C.; Nunnenmacher, K.; Schultz, G.; Plant, T.D. OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat. Cell Biol. 2000, 2, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, W.; Friedman, J.M. Abnormal osmotic regulation in trpv4-/- mice. Proc. Natl. Acad. Sci. USA 2003, 100, 13698–13703. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, W.; Tobin, D.M.; Bargmann, C.I.; Friedman, J.M. Mammalian TRPV4 (VR-OAC) directs behavioral responses to osmotic and mechanical stimuli in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2003, 100, 14531–14536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandri-Haber, N.; Yeh, J.J.; Boyd, A.E.; Parada, C.A.; Chen, X.; Reichling, D.B.; Levine, J.D. Hypotonicity induces TRPV4-mediated nociception in rat. Neuron 2003, 39, 497–511. [Google Scholar] [CrossRef]
- Alessandri-Haber, N.; Joseph, E.; Dina, O.A.; Liedtke, W.; Levine, J.D. TRPV4 mediates pain-related behavior induced by mild hypertonic stimuli in the presence of inflammatory mediator. Pain 2005, 118, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Kohler, R.; Heyken, W.T.; Heinau, P.; Schubert, R.; Si, H.; Kacik, M.; Busch, C.; Grgic, I.; Maier, T.; Hoyer, J. Evidence for a functional role of endothelial transient receptor potential V4 in shear stress-induced vasodilatation. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1495–1502. [Google Scholar] [CrossRef]
- Suzuki, M.; Mizuno, A.; Kodaira, K.; Imai, M. Impaired pressure sensation in mice lacking TRPV4. J. Biol. Chem. 2003, 278, 22664–22668. [Google Scholar] [CrossRef]
- Watanabe, H.; Vriens, J.; Prenen, J.; Droogmans, G.; Voets, T.; Nilius, B. Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature 2003, 424, 434–438. [Google Scholar] [CrossRef]
- Grant, A.D.; Cottrell, G.S.; Amadesi, S.; Trevisani, M.; Nicoletti, P.; Materazzi, S.; Altier, C.; Cenac, N.; Zamponi, G.W.; Bautista-Cruz, F.; et al. Protease-activated receptor 2 sensitizes the transient receptor potential vanilloid 4 ion channel to cause mechanical hyperalgesia in mice. J. Physiol. 2007, 578, 715–733. [Google Scholar] [CrossRef] [PubMed]
- Todaka, H.; Taniguchi, J.; Satoh, J.; Mizuno, A.; Suzuki, M. Warm temperature-sensitive transient receptor potential vanilloid 4 (TRPV4) plays an essential role in thermal hyperalgesia. J. Biol. Chem. 2004, 279, 35133–35138. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Gupta, R.; Jordt, S.E.; Chen, Y.; Liedtke, W.B. Regulation of Pain and Itch by TRP Channels. Neurosci. Bull. 2018, 34, 120–142. [Google Scholar] [CrossRef] [PubMed]
- Cenac, N.; Altier, C.; Chapman, K.; Liedtke, W.; Zamponi, G.; Vergnolle, N. Transient receptor potential vanilloid-4 has a major role in visceral hypersensitivity symptoms. Gastroenterology 2008, 135, 937–946.e2. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.H.; Ge, H.Y.; Arendt-Nielsen, L.; Wang, R.; Yue, S.W. A transient receptor potential vanilloid 4 contributes to mechanical allodynia following chronic compression of dorsal root ganglion in rats. Neurosci. Lett. 2008, 432, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, C.; Liu, L. Changes in osmolality modulate voltage-gated calcium channels in trigeminal ganglion neurons. Brain Res. 2008, 1208, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Williams, S.H.; McNulty, A.L.; Hong, J.H.; Lee, S.H.; Rothfusz, N.E.; Parekh, P.K.; Moore, C.; Gereau, R.W.T.; Taylor, A.B.; et al. Temporomandibular joint pain: A critical role for Trpv4 in the trigeminal ganglion. Pain 2013, 154, 1295–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Kanju, P.; Fang, Q.; Lee, S.H.; Parekh, P.K.; Lee, W.; Moore, C.; Brenner, D.; Gereau, R.W.T.; Wang, F.; et al. TRPV4 is necessary for trigeminal irritant pain and functions as a cellular formalin receptor. Pain 2014, 155, 2662–2672. [Google Scholar] [CrossRef] [Green Version]
- Rajasekhar, P.; Poole, D.P.; Liedtke, W.; Bunnett, N.W.; Veldhuis, N.A. P2Y1 Receptor Activation of the TRPV4 Ion Channel Enhances Purinergic Signaling in Satellite Glial Cells. J. Biol. Chem. 2015, 290, 29051–29062. [Google Scholar] [CrossRef]
- Alessandri-Haber, N.; Dina, O.A.; Joseph, E.K.; Reichling, D.; Levine, J.D. A transient receptor potential vanilloid 4-dependent mechanism of hyperalgesia is engaged by concerted action of inflammatory mediators. J. Neurosci. 2006, 26, 3864–3874. [Google Scholar] [CrossRef]
- Ichikawa, H.; Sugimoto, T. VR1-immunoreactive primary sensory neurons in the rat trigeminal ganglion. Brain Res. 2001, 890, 184–188. [Google Scholar] [CrossRef]
- Chaudhary, P.; Martenson, M.E.; Baumann, T.K. Vanilloid receptor expression and capsaicin excitation of rat dental primary afferent neurons. J. Dent. Res. 2001, 80, 1518–1523. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.K.; Lee, J.; Duraes, G.; Ro, J.Y. Lipopolysaccharide-induced pulpitis up-regulates TRPV1 in trigeminal ganglia. J. Dent. Res. 2011, 90, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Stenholm, E.; Bongenhielm, U.; Ahlquist, M.; Fried, K. VRl- and VRL-l-like immunoreactivity in normal and injured trigeminal dental primary sensory neurons of the rat. Acta Odontol. Scand. 2002, 60, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Chung, G.; Jo, H.J.; Kim, Y.S.; Bae, Y.C.; Jung, S.J.; Kim, J.S.; Oh, S.B. Characterization of dental nociceptive neurons. J. Dent. Res. 2011, 90, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Kim, M.S.; Fang, Z.; Li, H.Y.; Jung, S.J.; Choi, S.Y.; Lee, S.J.; Park, K.; Kim, J.S.; Oh, S.B. Functional expression of thermo-transient receptor potential channels in dental primary afferent neurons: Implication for tooth pain. J. Biol. Chem. 2006, 281, 17304–17311. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, Y.J.; Paik, S.K.; Cho, Y.S.; Kwon, T.G.; Ahn, D.K.; Kim, S.K.; Yoshida, A.; Bae, Y.C. Expression of metabotropic glutamate receptor mGluR5 in human dental pulp. J. Endod. 2009, 35, 690–694. [Google Scholar] [CrossRef]
- Renton, T.; Yiangou, Y.; Baecker, P.A.; Ford, A.P.; Anand, P. Capsaicin receptor VR1 and ATP purinoceptor P2X3 in painful and nonpainful human tooth pulp. J. Orofac. Pain 2003, 17, 245–250. [Google Scholar]
- Morgan, C.R.; Rodd, H.D.; Clayton, N.; Davis, J.B.; Boissonade, F.M. Vanilloid receptor 1 expression in human tooth pulp in relation to caries and pain. J. Orofac. Pain 2005, 19, 248–260. [Google Scholar]
- Bakri, M.M.; Yahya, F.; Munawar, K.M.M.; Kitagawa, J.; Hossain, M.Z. Transient receptor potential vanilloid 4 (TRPV4) expression on the nerve fibers of human dental pulp is upregulated under inflammatory condition. Arch. Oral Biol. 2018, 89, 94–98. [Google Scholar] [CrossRef]
- Okumura, R.; Shima, K.; Muramatsu, T.; Nakagawa, K.; Shimono, M.; Suzuki, T.; Magloire, H.; Shibukawa, Y. The odontoblast as a sensory receptor cell? The expression of TRPV1 (VR-1) channels. Arch. Histol. Cytol. 2005, 68, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeon, K.Y.; Chung, G.; Shin, M.S.; Jung, S.J.; Kim, J.S.; Oh, S.B. Adult rat odontoblasts lack noxious thermal sensitivity. J. Dent. Res. 2009, 88, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Sobhan, U.; Muramatsu, T.; Sato, M.; Ichikawa, H.; Sahara, Y.; Tazaki, M.; Shibukawa, Y. TRPV1-mediated calcium signal couples with cannabinoid receptors and sodium-calcium exchangers in rat odontoblasts. Cell Calcium 2012, 52, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Son, A.R.; Yang, Y.M.; Hong, J.H.; Lee, S.I.; Shibukawa, Y.; Shin, D.M. Odontoblast TRP channels and thermo/mechanical transmission. J. Dent. Res. 2009, 88, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Sobhan, U.; Tsumura, M.; Kuroda, H.; Soya, M.; Masamura, A.; Nishiyama, A.; Katakura, A.; Ichinohe, T.; Tazaki, M.; et al. Hypotonic-induced stretching of plasma membrane activates transient receptor potential vanilloid channels and sodium-calcium exchangers in mouse odontoblasts. J. Endod. 2013, 39, 779–787. [Google Scholar] [CrossRef]
- El Karim, I.A.; Linden, G.J.; Curtis, T.M.; About, I.; McGahon, M.K.; Irwin, C.R.; Lundy, F.T. Human odontoblasts express functional thermo-sensitive TRP channels: Implications for dentin sensitivity. Pain 2011, 152, 2211–2223. [Google Scholar] [CrossRef]
- El Karim, I.; McCrudden, M.T.; Linden, G.J.; Abdullah, H.; Curtis, T.M.; McGahon, M.; About, I.; Irwin, C.; Lundy, F.T. TNF-α-induced p38MAPK activation regulates TRPA1 and TRPV4 activity in odontoblast-like cells. Am. J. Pathol. 2015, 185, 2994–3002. [Google Scholar] [CrossRef]
- Wen, W.; Que, K.; Zang, C.; Wen, J.; Sun, G.; Zhao, Z.; Li, Y. Expression and distribution of three transient receptor potential vanilloid (TRPV) channel proteins in human odontoblast-like cells. J. Mol. Histol. 2017, 48, 367–377. [Google Scholar] [CrossRef]
- Egbuniwe, O.; Grover, S.; Duggal, A.K.; Mavroudis, A.; Yazdi, M.; Renton, T.; Di Silvio, L.; Grant, A.D. TRPA1 and TRPV4 activation in human odontoblasts stimulates ATP release. J. Dent. Res. 2014, 93, 911–917. [Google Scholar] [CrossRef]
- Sumoza-Toledo, A.; Penner, R. TRPM2: A multifunctional ion channel for calcium signalling. J. Physiol. 2011, 589, 1515–1525. [Google Scholar] [CrossRef]
- Aarts, M.M.; Tymianski, M. TRPMs and neuronal cell death. Pflug. Arch. 2005, 451, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Togashi, K.; Hara, Y.; Tominaga, T.; Higashi, T.; Konishi, Y.; Mori, Y.; Tominaga, M. TRPM2 activation by cyclic ADP-ribose at body temperature is involved in insulin secretion. EMBO J. 2006, 25, 1804–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perraud, A.L.; Fleig, A.; Dunn, C.A.; Bagley, L.A.; Launay, P.; Schmitz, C.; Stokes, A.J.; Zhu, Q.; Bessman, M.J.; Penner, R.; et al. ADP-ribose gating of the calcium-permeable LTRPC2 channel revealed by Nudix motif homology. Nature 2001, 411, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Cho, P.S.; Yang, Y.D.; Hwang, S.W. Nociceptive Roles of TRPM2 Ion Channel in Pathologic Pain. Mol. Neurobiol. 2018, 55, 6589–6600. [Google Scholar] [CrossRef] [PubMed]
- Grimm, C.; Kraft, R.; Sauerbruch, S.; Schultz, G.; Harteneck, C. Molecular and functional characterization of the melastatin-related cation channel TRPM3. J. Biol. Chem. 2003, 278, 21493–21501. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Chen, J.; Sun, L.; Wu, S.; Gray, K.R.; Rich, A.; Huang, M.; Lin, J.H.; Feder, J.N.; Janovitz, E.B.; et al. Expression and characterization of human transient receptor potential melastatin 3 (hTRPM3). J. Biol. Chem. 2003, 278, 20890–20897. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.F.; Loch, S.; Lambert, S.; Straub, I.; Mannebach, S.; Mathar, I.; Dufer, M.; Lis, A.; Flockerzi, V.; Philipp, S.E.; et al. Transient receptor potential M3 channels are ionotropic steroid receptors in pancreatic β cells. Nat. Cell Biol. 2008, 10, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S. TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Voets, T.; Vriens, J. TRPM3 in temperature sensing and beyond. Temperature 2015, 2, 201–213. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yao, G.; Savoia, C.; Touyz, R.M. Transient receptor potential melastatin 7 ion channels regulate magnesium homeostasis in vascular smooth muscle cells: Role of angiotensin II. Circ. Res. 2005, 96, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, C.; Perraud, A.L.; Johnson, C.O.; Inabe, K.; Smith, M.K.; Penner, R.; Kurosaki, T.; Fleig, A.; Scharenberg, A.M. Regulation of vertebrate cellular Mg2+ homeostasis by TRPM7. Cell 2003, 114, 191–200. [Google Scholar] [CrossRef]
- Krapivinsky, G.; Mochida, S.; Krapivinsky, L.; Cibulsky, S.M.; Clapham, D.E. The TRPM7 ion channel functions in cholinergic synaptic vesicles and affects transmitter release. Neuron 2006, 52, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Desai, B.N.; Navarro, B.; Donovan, A.; Andrews, N.C.; Clapham, D.E. Deletion of Trpm7 disrupts embryonic development and thymopoiesis without altering Mg2+ homeostasis. Science 2008, 322, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Turlova, E.; Bae, C.Y.J.; Deurloo, M.; Chen, W.; Barszczyk, A.; Horgen, F.D.; Fleig, A.; Feng, Z.P.; Sun, H.S. TRPM7 Regulates Axonal Outgrowth and Maturation of Primary Hippocampal Neurons. Mol. Neurobiol. 2016, 53, 595–610. [Google Scholar] [CrossRef] [PubMed]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- Reid, G.; Flonta, M.L. Ion channels activated by cold and menthol in cultured rat dorsal root ganglion neurones. Neurosci. Lett. 2002, 324, 164–168. [Google Scholar] [CrossRef]
- Thut, P.D.; Wrigley, D.; Gold, M.S. Cold transduction in rat trigeminal ganglia neurons in vitro. Neuroscience 2003, 119, 1071–1083. [Google Scholar] [CrossRef]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef]
- Abe, J.; Hosokawa, H.; Okazawa, M.; Kandachi, M.; Sawada, Y.; Yamanaka, K.; Matsumura, K.; Kobayashi, S. TRPM8 protein localization in trigeminal ganglion and taste papillae. Brain Res. 2005, 136, 91–98. [Google Scholar] [CrossRef]
- Dhaka, A.; Murray, A.N.; Mathur, J.; Earley, T.J.; Petrus, M.J.; Patapoutian, A. TRPM8 is required for cold sensation in mice. Neuron 2007, 54, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Siemens, J.; Glazer, J.M.; Tsuruda, P.R.; Basbaum, A.I.; Stucky, C.L.; Jordt, S.E.; Julius, D. The menthol receptor TRPM8 is the principal detector of environmental cold. Nature 2007, 448, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Colburn, R.W.; Lubin, M.L.; Stone, D.J., Jr.; Wang, Y.; Lawrence, D.; D’Andrea, M.R.; Brandt, M.R.; Liu, Y.; Flores, C.M.; Qin, N. Attenuated cold sensitivity in TRPM8 null mice. Neuron 2007, 54, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Michot, B.; Lee, C.S.; Gibbs, J.L. TRPM8 and TRPA1 do not contribute to dental pulp sensitivity to cold. Sci. Rep. 2018, 8, 13198. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Sobhan, U.; Sato, M.; Shimada, M.; Nishiyama, A.; Kawaguchi, A.; Soya, M.; Kuroda, H.; Tazaki, M.; Shibukawa, Y. Functional expression of TRPM8 and TRPA1 channels in rat odontoblasts. PLoS ONE 2013, 8, e82233. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, K.; Ikeda, H.; Kawashima, N.; Okiji, T. Transient receptor potential melastatin (TRPM) 8 is expressed in freshly isolated native human odontoblasts. Arch. Oral Biol. 2017, 75, 55–61. [Google Scholar] [CrossRef]
- Kwon, M.; Baek, S.H.; Park, C.K.; Chung, G.; Oh, S.B. Single-cell RT-PCR and immunocytochemical detection of mechanosensitive transient receptor potential channels in acutely isolated rat odontoblasts. Arch. Oral Biol. 2014, 59, 1266–1271. [Google Scholar] [CrossRef]
- Won, J.; Vang, H.; Kim, J.H.; Lee, P.R.; Kang, Y.; Oh, S.B. TRPM7 Mediates Mechanosensitivity in Adult Rat Odontoblasts. J. Dent. Res. 2018, 97, 1039–1046. [Google Scholar] [CrossRef]
- Nakano, Y.; Le, M.H.; Abduweli, D.; Ho, S.P.; Ryazanova, L.V.; Hu, Z.; Ryazanov, A.G.; Den Besten, P.K.; Zhang, Y. A Critical Role of TRPM7 As an Ion Channel Protein in Mediating the Mineralization of the Craniofacial Hard Tissues. Front. Physiol. 2016, 7, 258. [Google Scholar] [CrossRef]
- Won, J.; Kim, J.H.; Oh, S.B. Molecular expression of Mg2+ regulator TRPM7 and CNNM4 in rat odontoblasts. Arch. Oral Biol. 2018, 96, 182–188. [Google Scholar] [CrossRef]
- Nilius, B.; Appendino, G.; Owsianik, G. The transient receptor potential channel TRPA1: From gene to pathophysiology. Pflug. Arch. 2012, 464, 425–458. [Google Scholar] [CrossRef] [PubMed]
- Nassenstein, C.; Kwong, K.; Taylor-Clark, T.; Kollarik, M.; Macglashan, D.M.; Braun, A.; Undem, B.J. Expression and function of the ion channel TRPA1 in vagal afferent nerves innervating mouse lungs. J. Physiol. 2008, 586, 1595–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozawa, K.; Kawabata-Shoda, E.; Doihara, H.; Kojima, R.; Okada, H.; Mochizuki, S.; Sano, Y.; Inamura, K.; Matsushime, H.; Koizumi, T.; et al. TRPA1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3408–3413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ro, J.Y.; Lee, J.S.; Zhang, Y. Activation of TRPV1 and TRPA1 leads to muscle nociception and mechanical hyperalgesia. Pain 2009, 144, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Investing. 2005, 115, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, D.M.; Jordt, S.E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.S.; Woolf, C.J.; Corey, D.P. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Jordt, S.E.; Bautista, D.M.; Chuang, H.H.; McKemy, D.D.; Zygmunt, P.M.; Hogestatt, E.D.; Meng, I.D.; Julius, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef]
- Bautista, D.M.; Movahed, P.; Hinman, A.; Axelsson, H.E.; Sterner, O.; Hogestatt, E.D.; Julius, D.; Jordt, S.E.; Zygmunt, P.M. Pungent products from garlic activate the sensory ion channel TRPA1. Proc. Natl. Acad. Sci. USA 2005, 102, 12248–12252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, E.T.; Rowland, K.; Gautam, M. Tooth injury increases expression of the cold sensitive TRP channel TRPA1 in trigeminal neurons. Arch. Oral Biol. 2011, 56, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Jung, H.K.; Kwon, T.K.; Kim, C.S.; Cho, J.H.; Ahn, D.K.; Bae, Y.C. Expression of transient receptor potential ankyrin 1 in human dental pulp. J. Endod. 2012, 38, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- El Karim, I.A.; McCrudden, M.T.; McGahon, M.K.; Curtis, T.M.; Jeanneau, C.; Giraud, T.; Irwin, C.R.; Linden, G.J.; Lundy, F.T.; About, I. Biodentine Reduces Tumor Necrosis Factor α-induced TRPA1 Expression in Odontoblastlike Cells. J. Endod. 2016, 42, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Song, Z.; Chen, L.; Wang, R.; Huang, S.; Qin, W.; Guo, J.; Lin, Z. Role of transient receptor potential channel 6 in the odontogenic differentiation of human dental pulp cells. Exp. Ther. Med. 2017, 14, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Chen, L.; Guo, J.; Qin, W.; Wang, R.; Huang, S.; Yang, X.; Tian, Y.; Lin, Z. The Role of Transient Receptor Potential Cation Channel, Subfamily C, Member 1 in the Odontoblast-like Differentiation of Human Dental Pulp Cells. J. Endod. 2017, 43, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Fahlbusch, M.; Gudermann, T. Classical Transient Receptor Potential 1 (TRPC1): Channel or Channel Regulator? Cells 2014, 3, 939–962. [Google Scholar] [CrossRef] [Green Version]
- Ambudkar, I.S. TRPC1: A core component of store-operated calcium channels. Biochem. Soc. Trans. 2007, 35, 96–100. [Google Scholar] [CrossRef]
- Strubing, C.; Krapivinsky, G.; Krapivinsky, L.; Clapham, D.E. TRPC1 and TRPC5 form a novel cation channel in mammalian brain. Neuron 2001, 29, 645–655. [Google Scholar] [CrossRef]
- Valero, M.L.; Caminos, E.; Juiz, J.M.; Martinez-Galan, J.R. TRPC1 and metabotropic glutamate receptor expression in rat auditory midbrain neurons. J. Neurosci. Res. 2015, 93, 964–972. [Google Scholar] [CrossRef]
- Broker-Lai, J.; Kollewe, A.; Schindeldecker, B.; Pohle, J.; Nguyen Chi, V.; Mathar, I.; Guzman, R.; Schwarz, Y.; Lai, A.; Weissgerber, P.; et al. Heteromeric channels formed by TRPC1, TRPC4 and TRPC5 define hippocampal synaptic transmission and working memory. EMBO J. 2017, 36, 2770–2789. [Google Scholar] [CrossRef] [PubMed]
- Elg, S.; Marmigere, F.; Mattsson, J.P.; Ernfors, P. Cellular subtype distribution and developmental regulation of TRPC channel members in the mouse dorsal root ganglion. J. Comp. Neurol. 2007, 503, 35–46. [Google Scholar] [CrossRef]
- Garrison, S.R.; Dietrich, A.; Stucky, C.L. TRPC1 contributes to light-touch sensation and mechanical responses in low-threshold cutaneous sensory neurons. J. Neurophysiol. 2012, 107, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spassova, M.A.; Hewavitharana, T.; Xu, W.; Soboloff, J.; Gill, D.L. A common mechanism underlies stretch activation and receptor activation of TRPC6 channels. Proc. Natl. Acad. Sci. USA 2006, 103, 16586–16591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroto, R.; Raso, A.; Wood, T.G.; Kurosky, A.; Martinac, B.; Hamill, O.P. TRPC1 forms the stretch-activated cation channel in vertebrate cells. Nat. Cell Biol. 2005, 7, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Dina, O.A.; Chen, X.; Levine, J.D. TRPC1 and TRPC6 channels cooperate with TRPV4 to mediate mechanical hyperalgesia and nociceptor sensitization. J. Neurosci. 2009, 29, 6217–6228. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, R.; Tokuda, M.; Sakuta, T.; Nagaoka, S.; Torii, M. Expression and characterization of vanilloid receptor subtype 1 in human dental pulp cell cultures. J. Endod. 2005, 31, 652–658. [Google Scholar] [CrossRef]
- El Karim, I.A.; Linden, G.J.; Curtis, T.M.; About, I.; McGahon, M.K.; Irwin, C.R.; Killough, S.A.; Lundy, F.T. Human dental pulp fibroblasts express the “cold-sensing” transient receptor potential channels TRPA1 and TRPM8. J. Endod. 2011, 37, 473–478. [Google Scholar] [CrossRef]
- Rowland, K.C.; Kanive, C.B.; Wells, J.E.; Hatton, J.F. TRPM2 immunoreactivity is increased in fibroblasts, but not nerves, of symptomatic human dental pulp. J. Endod. 2007, 33, 245–248. [Google Scholar] [CrossRef]
- Hashimoto, M.; Ito, S.; Tay, F.R.; Svizero, N.R.; Sano, H.; Kaga, M.; Pashley, D.H. Fluid movement across the resin-dentin interface during and after bonding. J. Dent. Res. 2004, 83, 843–848. [Google Scholar] [CrossRef]
- Kim, S. Thermal stimuli in dentinal sensitivity. Endod. Dent. Traumatol. 1986, 2, 138–140. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Schuessler, G.; Chien, S. Measurement of blood flow in the dental pulp of dogs with the 133xenon washout method. Arch. Oral Biol. 1983, 28, 501–505. [Google Scholar] [CrossRef]
- Narhi, M. Activation of dental pulp nerves of the cat and the dog with hydrostatic pressure. Proc. Finn. Dent. Soc. 1978, 74, 1–63. [Google Scholar] [PubMed]
- Pashley, D.H. Sensitivity of dentin to chemical stimuli. Endod. Dent. Traumatol. 1986, 2, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Roudaut, Y.; Lonigro, A.; Coste, B.; Hao, J.; Delmas, P.; Crest, M. Touch sense: Functional organization and molecular determinants of mechanosensitive receptors. Channels 2012, 6, 234–245. [Google Scholar] [CrossRef]
- Christensen, A.P.; Corey, D.P. TRP channels in mechanosensation: Direct or indirect activation? Nat. Rev. 2007, 8, 510–521. [Google Scholar] [CrossRef]
- Narhi, M.; Jyvasjarvi, E.; Virtanen, A.; Huopaniemi, T.; Ngassapa, D.; Hirvonen, T. Role of intradental A- and C-type nerve fibres in dental pain mechanisms. Proc. Finn. Dent. Soc. 1992, 88, 507–516. [Google Scholar]
- Narhi, M.; Jyvasjarvi, E.; Hirvonen, T.; Huopaniemi, T. Activation of heat-sensitive nerve fibres in the dental pulp of the cat. Pain 1982, 14, 317–326. [Google Scholar] [CrossRef]
- Hamilton, S.G. ATP and pain. Pain Pract. 2002, 2, 289–294. [Google Scholar] [CrossRef]
- Wirkner, K.; Sperlagh, B.; Illes, P. P2X3 receptor involvement in pain states. Mol. Neurobiol. 2007, 36, 165–183. [Google Scholar] [CrossRef]
- Sato, M.; Ogura, K.; Kimura, M.; Nishi, K.; Ando, M.; Tazaki, M.; Shibukawa, Y. Activation of Mechanosensitive Transient Receptor Potential/Piezo Channels in Odontoblasts Generates Action Potentials in Cocultured Isolectin B4-negative Medium-sized Trigeminal Ganglion Neurons. J. Endod. 2018, 44, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, C.; Fujita, T.; Malmstrom, H.S.; Nedergaard, M.; Ren, Y.F.; Dirksen, R.T. External Dentin Stimulation Induces ATP Release in Human Teeth. J. Dent. Res. 2015, 94, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, H.; Braun, N. Extracellular metabolism of nucleotides in the nervous system. J. Auton. Pharmacol. 1996, 16, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, L.; Wang, Q.; Pelletier, J.; Fausther, M.; Sevigny, J.; Malmstrom, H.S.; Dirksen, R.T.; Ren, Y.F. Expression of ecto-ATPase NTPDase2 in human dental pulp. J. Dent. Res. 2012, 91, 261–267. [Google Scholar] [CrossRef]
- Burnstock, G. Physiology and pathophysiology of purinergic neurotransmission. Physiol. Rev. 2007, 87, 659–797. [Google Scholar] [CrossRef]
- Kuroda, H.; Shibukawa, Y.; Soya, M.; Masamura, A.; Kasahara, M.; Tazaki, M.; Ichinohe, T. Expression of P2X(1) and P2X(4) receptors in rat trigeminal ganglion neurons. Neuroreport 2012, 23, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Bo, X.; Burnstock, G. Localization of ATP-gated P2X receptor immunoreactivity in rat sensory and sympathetic ganglia. Neurosci. Lett. 1998, 256, 105–108. [Google Scholar] [CrossRef]
- Jiang, J.; Gu, J. Expression of adenosine triphosphate P2X3 receptors in rat molar pulp and trigeminal ganglia. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2002, 94, 622–626. [Google Scholar] [CrossRef]
- Alavi, A.M.; Dubyak, G.R.; Burnstock, G. Immunohistochemical evidence for ATP receptors in human dental pulp. J. Dent. Res. 2001, 80, 476–483. [Google Scholar] [CrossRef]
- Cook, S.P.; Vulchanova, L.; Hargreaves, K.M.; Elde, R.; McCleskey, E.W. Distinct ATP receptors on pain-sensing and stretch-sensing neurons. Nature 1997, 387, 505–508. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Sato, M.; Kimura, M.; Ichinohe, T.; Tazaki, M.; Shibukawa, Y. Expression and function of purinergic P2Y12 receptors in rat trigeminal ganglion neurons. Neurosci. Res. 2015, 98, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, A.; Sato, M.; Kimura, M.; Katakura, A.; Tazaki, M.; Shibukawa, Y. Intercellular signal communication among odontoblasts and trigeminal ganglion neurons via glutamate. Cell Calcium 2016, 60, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.S.; Ryu, C.H.; Won, J.H.; Vang, H.; Oh, S.B.; Ro, J.Y.; Bae, Y.C. Rat odontoblasts may use glutamate to signal dentin injury. Neuroscience 2016, 335, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Horst, O.V.; Tompkins, K.A.; Coats, S.R.; Braham, P.H.; Darveau, R.P.; Dale, B.A. TGF-β1 Inhibits TLR-mediated odontoblast responses to oral bacteria. J. Dent. Res. 2009, 88, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.W.; Zhang, W.; Ren, B.P.; Zeng, J.F.; Ling, J.Q. Expression of toll like receptor 4 in normal human odontoblasts and dental pulp tissue. J. Endod. 2006, 32, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Mutoh, N.; Tani-Ishii, N.; Tsukinoki, K.; Chieda, K.; Watanabe, K. Expression of toll-like receptor 2 and 4 in dental pulp. J. Endod. 2007, 33, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Staquet, M.J.; Carrouel, F.; Keller, J.F.; Baudouin, C.; Msika, P.; Bleicher, F.; Kufer, T.A.; Farges, J.C. Pattern-recognition receptors in pulp defense. Adv. Dent. Res. 2011, 23, 296–301. [Google Scholar] [CrossRef]
- Rechenberg, D.K.; Galicia, J.C.; Peters, O.A. Biological Markers for Pulpal Inflammation: A Systematic Review. PLoS ONE 2016, 11, e0167289. [Google Scholar] [CrossRef]
- Hahn, C.L.; Liewehr, F.R. Innate immune responses of the dental pulp to caries. J. Endod. 2007, 33, 643–651. [Google Scholar] [CrossRef]
- Lundy, F.T.; Linden, G.J. Neuropeptides and Neurogenic Mechanisms in Oral and Periodontal Inflammation. Crit. Rev. Oral Biol. Med. 2004, 15, 82–98. [Google Scholar] [CrossRef]
- Caviedes-Bucheli, J.; Munoz, H.R.; Azuero-Holguin, M.M.; Ulate, E. Neuropeptides in dental pulp: The silent protagonists. J. Endod. 2008, 34, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Sensory TRP channels: The key transducers of nociception and pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 73–118. [Google Scholar] [PubMed]
- Hossain, M.Z.; Unno, S.; Ando, H.; Masuda, Y.; Kitagawa, J. Neuron-Glia Crosstalk and Neuropathic Pain: Involvement in the Modulation of Motor Activity in the Orofacial Region. Int. J. Mol. Sci. 2017, 18, 2051. [Google Scholar] [CrossRef] [PubMed]
- Patapoutian, A.; Tate, S.; Woolf, C.J. Transient receptor potential channels: Targeting pain at the source. Nat. Rev. Drug Discov. 2009, 8, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Rodd, H.D.; Boissonade, F.M. Substance P expression in human tooth pulp in relation to caries and pain experience. Eur. J. Oral Sci. 2000, 108, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Killough, S.A.; Lundy, F.T.; Irwin, C.R. Substance P expression by human dental pulp fibroblasts: A potential role in neurogenic inflammation. J. Endod. 2009, 35, 73–77. [Google Scholar] [CrossRef]
- Sacerdote, P.; Levrini, L. Peripheral mechanisms of dental pain: The role of substance P. Mediat. Inflamm. 2012, 2012, 951920. [Google Scholar] [CrossRef]
- Nakanishi, T.; Takegawa, D.; Hirao, K.; Takahashi, K.; Yumoto, H.; Matsuo, T. Roles of dental pulp fibroblasts in the recognition of bacterium-related factors and subsequent development of pulpitis. Jpn. Dent. Sci. Rev. 2011, 47, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Suvas, S. Role of Substance P Neuropeptide in Inflammation, Wound Healing, and Tissue Homeostasis. J. Immunol. 2017, 199, 1543–1552. [Google Scholar] [CrossRef]
- Goehler, L.E.; Gaykema, R.P.; Opitz, N.; Reddaway, R.; Badr, N.; Lyte, M. Activation in vagal afferents and central autonomic pathways: Early responses to intestinal infection with Campylobacter jejuni. Brain Behav. Immun. 2005, 19, 334–344. [Google Scholar] [CrossRef]
- Chiu, I.M.; Heesters, B.A.; Ghasemlou, N.; Von Hehn, C.A.; Zhao, F.; Tran, J.; Wainger, B.; Strominger, A.; Muralidharan, S.; Horswill, A.R.; et al. Bacteria activate sensory neurons that modulate pain and inflammation. Nature 2013, 501, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonen, B.; Alpizar, Y.A.; Meseguer, V.M.; Talavera, K. TRP Channels as Sensors of Bacterial Endotoxins. Toxins 2018, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Diogenes, A.; Ferraz, C.C.; Akopian, A.N.; Henry, M.A.; Hargreaves, K.M. LPS sensitizes TRPV1 via activation of TLR4 in trigeminal sensory neurons. J. Dent. Res. 2011, 90, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.C.; Henry, M.A.; Hargreaves, K.M.; Diogenes, A. Lipopolysaccharide from Porphyromonas gingivalis sensitizes capsaicin-sensitive nociceptors. J. Endod. 2011, 37, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Du, Y.; Cai, W.K.; Kuang, R.; Chang, T.; Zhang, Z.; Yang, Y.X.; Sun, C.; Li, Z.Y.; Kuang, F. Toll-like receptor 4 signaling in neurons of trigeminal ganglion contributes to nociception induced by acute pulpitis in rats. Sci. Rep. 2015, 5, 12549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meseguer, V.; Alpizar, Y.A.; Luis, E.; Tajada, S.; Denlinger, B.; Fajardo, O.; Manenschijn, J.A.; Fernandez-Pena, C.; Talavera, A.; Kichko, T.; et al. TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nat. Commun. 2014, 5, 3125. [Google Scholar] [CrossRef] [PubMed]
- Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Lopez-Requena, A.; Voets, T.; Talavera, K. Differential effects of lipopolysaccharide on mouse sensory TRP channels. Cell Calcium 2018, 73, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Alpizar, Y.A.; Boonen, B.; Sanchez, A.; Jung, C.; Lopez-Requena, A.; Naert, R.; Steelant, B.; Luyts, K.; Plata, C.; De Vooght, V.; et al. TRPV4 activation triggers protective responses to bacterial lipopolysaccharides in airway epithelial cells. Nat. Commun. 2017, 8, 1059. [Google Scholar] [CrossRef] [Green Version]
- Pashley, D.H. How can sensitive dentine become hypersensitive and can it be reversed? J. Den. 2013, 41, S49–S55. [Google Scholar] [CrossRef] [Green Version]
- Stenvik, A.; Iversen, J.; Mjor, I.A. Tissue pressure and histology of normal and inflamed tooth pulps in macaque monkeys. Arch. Oral Biol. 1972, 17, 1501–1511. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Daud, S.; Nambiar, P.; Razak, F.A.; Ab-Murat, N.; Saub, R.; Bakri, M.M. Correlation between numbers of cells in human dental pulp and age: Implications for age estimation. Arch. Oral Biol. 2017, 80, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Daud, S.; Nambiar, P.; Hossain, M.Z.; Rahman, M.R.; Bakri, M.M. Changes in cell density and morphology of selected cells of the ageing human dental pulp. Gerodontology 2016, 33, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.C.; Huang, F.M.; Lin, C.S.; Liu, C.M.; Lai, C.C.; Chang, Y.C. Induction of interleukin-8 gene expression by black-pigmented Bacteroides in human pulp fibroblasts and osteoblasts. Int. Endod. J. 2003, 36, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Coil, J.; Tam, E.; Waterfield, J.D. Proinflammatory cytokine profiles in pulp fibroblasts stimulated with lipopolysaccharide and methyl mercaptan. J. Endod. 2004, 30, 88–91. [Google Scholar] [CrossRef]
- Patel, T.; Park, S.H.; Lin, L.M.; Chiappelli, F.; Huang, G.T. Substance P induces interleukin-8 secretion from human dental pulp cells. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2003, 96, 478–485. [Google Scholar] [CrossRef]
- Park, S.H.; Hsiao, G.Y.; Huang, G.T. Role of substance P and calcitonin gene-related peptide in the regulation of interleukin-8 and monocyte chemotactic protein-1 expression in human dental pulp. Int. Endod. J. 2004, 37, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Kojima, T.; Kanekawa, M.; Aihara, N.; Nogimura, A.; Kasai, K. Neuropeptides stimulate production of interleukin-1β, interleukin-6, and tumor necrosis factor-α in human dental pulp cells. Inflamm. Res. 2004, 53, 199–204. [Google Scholar] [CrossRef]
- Binshtok, A.M.; Bean, B.P.; Woolf, C.J. Inhibition of nociceptors by TRPV1-mediated entry of impermeant sodium channel blockers. Nature 2007, 449, 607–610. [Google Scholar] [CrossRef]
- Zakir, H.M.; Mostafeezur, R.M.; Suzuki, A.; Hitomi, S.; Suzuki, I.; Maeda, T.; Seo, K.; Yamada, Y.; Yamamura, K.; Lev, S.; et al. Expression of TRPV1 channels after nerve injury provides an essential delivery tool for neuropathic pain attenuation. PLoS ONE 2012, 7, e44023. [Google Scholar] [CrossRef]
- Kim, H.Y.; Kim, K.; Li, H.Y.; Chung, G.; Park, C.K.; Kim, J.S.; Jung, S.J.; Lee, M.K.; Ahn, D.K.; Hwang, S.J.; et al. Selectively targeting pain in the trigeminal system. Pain 2010, 150, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Badral, B.; Davies, A.J.; Kim, Y.H.; Ahn, J.S.; Hong, S.D.; Chung, G.; Kim, J.S.; Oh, S.B. Pain fiber anesthetic reduces brainstem Fos after tooth extraction. J. Dent. Res. 2013, 92, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.H.; Piao, Z.G.; Kim, Y.B.; Lee, C.H.; Lee, J.K.; Park, K.; Kim, J.S.; Oh, S.B. Activation of vanilloid receptor 1 (VR1) by eugenol. J. Dent. Res. 2003, 82, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies. Pharmaceuticals 2016, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Treanor, J.J.; Garami, A.; Fang, L.; Surapaneni, S.; Akrami, A.; Alvarez, F.; Bak, A.; Darling, M.; Gore, A.; et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. Pain 2008, 136, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Quiding, H.; Jonzon, B.; Svensson, O.; Webster, L.; Reimfelt, A.; Karin, A.; Karlsten, R.; Segerdahl, M. TRPV1 antagonistic analgesic effect: A randomized study of AZD1386 in pain after third molar extraction. Pain 2013, 154, 808–812. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.Z.; Bakri, M.M.; Yahya, F.; Ando, H.; Unno, S.; Kitagawa, J. The Role of Transient Receptor Potential (TRP) Channels in the Transduction of Dental Pain. Int. J. Mol. Sci. 2019, 20, 526. https://doi.org/10.3390/ijms20030526
Hossain MZ, Bakri MM, Yahya F, Ando H, Unno S, Kitagawa J. The Role of Transient Receptor Potential (TRP) Channels in the Transduction of Dental Pain. International Journal of Molecular Sciences. 2019; 20(3):526. https://doi.org/10.3390/ijms20030526
Chicago/Turabian StyleHossain, Mohammad Zakir, Marina Mohd Bakri, Farhana Yahya, Hiroshi Ando, Shumpei Unno, and Junichi Kitagawa. 2019. "The Role of Transient Receptor Potential (TRP) Channels in the Transduction of Dental Pain" International Journal of Molecular Sciences 20, no. 3: 526. https://doi.org/10.3390/ijms20030526