1. Introduction
Constipation is an acute or chronic gastrointestinal disease characterized by infrequent bowel movements, difficulty during defecation, incomplete bowel evacuation, and hard and dry feces [
1]. Although there are numerous treatments for this disease, constipation is best treated by making simple diet changes by incorporating more fiber into the diet, by drinking plenty of fluids, and by adding exercise in the patient’s daily routine. Chemical drugs (laxatives) such as senna, correctol, exlax, senokot, and gaviscon are frequently administered to patients with constipation [
2]. These drugs act as stimulants to increase the bulkiness and to soften stool or as osmotic agents that enhance water flow into the colon to promote elimination and to trigger bowel movements. However, most laxatives have accompanying undesirable side effects, including artery contraction, coronary spasms, and myocardial infarction [
3,
4,
5]. Hence, the primary focus for constipation therapy remains on the identification of novel laxatives derived from natural products that have no side effects.
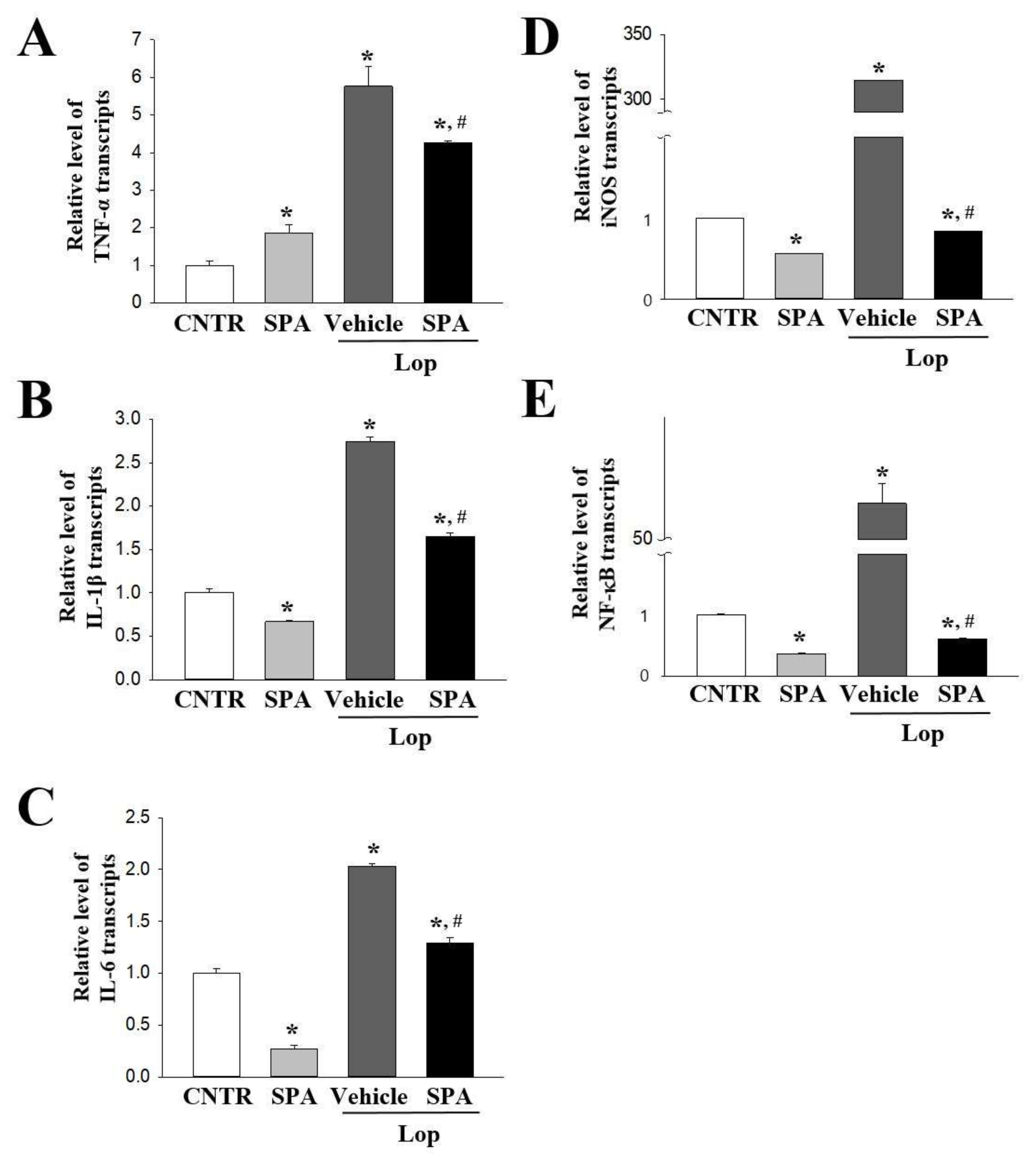
Since very few studies have investigated the inflammatory responses and cholinergic regulation during the laxative effects of natural products, the subjects remain relatively unknown. Atractylodin, a widely used oriental medicine, decreases the expression of proinflammatory cytokines (TNF-α, IL-1β, and IL-6) and inflammatory mediators (iNOS and NF-κB) as well as reduces the number and moisture content of stools in constipation-prominent models [
6]. Also, the downstream signaling pathway of some mAChRs belonging to cholinergic regulation significantly recovered in the Lop-induced constipation models after the treatment with extracts from
Liriope platyphylla [
5], Galla Rhois [
7], or red
L. platyphylla (RLP) [
8]. However, these previous studies did not analyze the anti-inflammatory effects and cholinergic regulation concurrently, although these two responses need to be considered as key factors of chronic constipation.
To date, there are few reports of major active ingredients, although many studies have reported the laxative effects of various herbal plants. Genkwanin-5-O-beta primeveroside collected from agarwood was first identified as the main constituent contributing to laxative effects. When exposed to 1000 mg/kg of the compound, the frequency and weight of stools in male ddY mice were significantly increased [
9]. Based on the results of previous studies, tannin and saponin have recently been considered potential laxative candidates [
10,
11]. Tannin is distributed in several plant extracts that exhibit laxative activities, based on their ability to increase the number and weight of feces. Tannin is also detected in the leaf aqueous extract of
Mareya micrantha Mull. Arg. [
10]; the methanol extract of
Senna macranthera [
11]; the aqueous extracts of
Aloe ferox Mill. [
12]; the aqueous-methanol extracts of
Fumaria parviflora [
13],
Urginea indica Kunth. [
14],
Phyllanthus emblica [
13], and gallotanin-enriched galla rhois (GEGR) [
7]. Herbal plants containing saponin also exhibit laxative properties including the ability to increase intestinal motility, the frequency and weight of stools, and the ileum tension. Such compounds include extracts of
A. ferox Mill. [
12],
Ficus carica paste [
15], and the aqueous extract of
L. platyphylla (AEtLP) [
5]. Furthermore, saponin as well as extracts containing saponin inhibit the pro-inflammatory responses in cecal ligation, puncture (CLP)-induced sepsis mice, and dextran sulfate sodium (DSS)-induced mouse colitis models [
16,
17,
18]. Ginsenoside Re exerts cholinergic stimulation and inhibitory effects on the contractility of isolated jejunal segments [
19,
20]. The laxative effects, inflammatory response, and cholinergic regulation of saponin-enriched extracts of
Asparagus cochinchinensis (SPA) in the Lop-induced constipation model have not been fully investigated, although seven steroidal saponins were detected in the roots of
A. cochinchinensis [
21,
22].
To evaluate the possibility of developing a new natural medicine, the present study was undertaken to investigate the inflammatory responses and cholinergic regulation during the laxative activity of SPA in a Lop-induced constipation model. Our results provide the first scientific evidence that SPA is a saponin-containing natural product that successfully induces laxative effects in the constipated animal model through the suppression of the inflammatory response and the recovery of the muscarinic cholinergic regulation.
3. Discussion
Attention has recently been focused on several herbal plants and natural products as novel therapeutic drugs for the treatment of gastrointestinal diseases due to their significant potential to alleviate symptoms of such diseases [
5,
7,
9]. In an effort to identify novel drugs for the treatment of constipation, we investigated the inflammatory responses and the cholinergic regulation exerted during the laxative effects of SPA in Lop-induced constipated SD rats.
A. cochinchinensis roots have been implicated in the recovery of neuronal cells and gut injury in studies using animals and Drosophila [
22,
23,
24]. The results of the current study clearly demonstrate that SPA induces laxative effects, including the elevation of stool excretion and the recovery of histological changes induced by Lop injection in the colon. Furthermore, we believe our data to be the first to demonstrate that SPA suppresses proinflammatory cytokines and their mediators, upregulates mucin secretion, as well as downregulates the mAChRs signaling pathway while effectively alleviating constipation.
The main constituents of the tuberous roots of
A. cochinchinesis consist of 19 amino acids, polysaccharides, and more than 20 multifunctional compounds [
25,
26,
27]. These functional compounds include β-sitosterol [
3], daucosterol [
4], n-ethatriacontanoic acid [
5], palmitic acid [
9], 9-heptacosylene [
10], smilagenin [
11], diosgenin [
12], sarsasapogenin-3-O-β-D-glucoside feeding grapes imidacloprid [
13], 5-methoxy methyl furfural, yame sapogenin, diosgenin-3-O-β-D imidacloprid feeding glucose glycosides [
13,
14], aspacochioside D [
7], iso-agatharesinoside [
15], and seven steroidal saponins [
21]. The roots of
A. cochinchinesis have several therapeutic properties, including antiaging and antioxidant effects [
22,
28,
29], antibacterial inflammatory effects [
22,
26,
27], anticancer effects [
23,
24,
30], and hepatoprotective activity [
25]. Moreover, they are administered in combination with other herbs to treat lung diseases, immune system-related diseases, and aging [
9,
10]. Recently, several studies for the medicinal use with
A. cochinchinensis suggest its therapeutic effects for constipation, since constipation is tightly associated with the dysfunction of the neuronal and digestive systems. The root extract with
A. cochinchinensis significantly increases the activity of superoxide dismutase (SOD) and reduces the contents of malondialdehyde (MDA) in the brain [
23]. Moreover, an aqueous extract with
A. cochinchinesis stimulates the nerve growth factor (NGF) expression and secretion in neuronal cells [
24]. Furthermore, changes associated with gut injuries induced by metal ion treatment, such as the survival rate of epithelial cells and morphological changes in the gut, were effectively alleviated by treatment with the aqueous
A. cochinchinesis extract [
25]. However, the laxative effects of
A. cochinchinesis have not been examined in a constipated animal model to date, although the laxative effects of
L. platyphylla are well-studied. This present study provides strong evidence of the novel function of
A. cochinchinesis extract containing high concentrations of saponin, although further studies are required to understand how saponin affects the suppression of the inflammatory response and regulates the motility of the gastrointestinal tract.
Some extracts and compounds derived from
A. cochinchinensis roots are reported to induce anti-inflammatory effects in various diseases. The ethanolic extract from
A. cochinchinensis and methyl protodioscin (MP) effectively reduce acute and chronic skin inflammation induced by 12-O-tetradecanoyl-phorbol-13-acetate (TPA)-induced mouse ear edema and inhibits the airway inflammation in a lipopolysaccharide (LPS)-induced acute lung injury model [
31,
32]. A SPA and butanol extract of
A. cochinchinensis roots fermented with
Weissella cibaria (BAW) have been shown to prevent airway inflammation and remodeling in the lung tissue of an ovalbumin (OVA)-induced asthma model [
33,
34]. However, there have been no studies investigating the anti-inflammatory response of
A. cochinchinensis roots in the gastrointestinal tract of animals with inflammatory disease in spite of the above researches providing few evidences. Our results, therefore, present a clue for the novel role of SPA in alleviating the symptoms of constipation accompanied by an inflammatory response in the colon of the Lop-induced model. Furthermore, the ameliorating effects of SPA on the inflammatory mechanism is very similar to the outcomes observed in the atractylodin-treated constipation-prominent model [
6]. Especially, the increased levels of proinflammatory cytokines were remarkably reversed after exposure to SPA and atractylodin, although the decrease rate was greater in the SPA-treated group than in atractylodin-treated group. Moreover, the results of the present study provide clues to the advantages or disadvantages for a single treatment of SPA in the physiological condition of SD rats. This extract promoted the decrease of paneth cells number and some cytokines expression as well as the enhancement of urine volume. At the same time, SPA induced the increase of PKC phosphorylation and the decrease of PI3K phosphorylation. These properties of SPA were showed the possibility that SPA can be considered as a versatile substance to treat human diseases.
Few studies have provided a theoretical basis for microbial treatment in chronic constipation by demonstrating a connection between constipation and intestinal flora disturbance. One treatment includes probiotics, prebiotics, symbiotics, and fecal microbiota transplantation (FMT) and is widely accepted because of the safety, convenience, and curative nature [
35]. The oral administration of
Lactobacillus casei Qian for 9 days induces an increase in the gastrointestinal transit, feces number, and some gastrointestinal hormone levels in carbon-induced constipated mice [
36]. In patients with gastrointestinal symptoms, treatment with EpiCor
® fermentate (consisting of dried yeast extracts) for 2 weeks improves the constipation-related symptoms by modulating the gut microbiome [
37]. Most studies evaluating the therapeutic effects of microbial treatment require administration and observation for a duration exceeding 1 week. However, in the current study, SPA was administered only once and the observation was for 2 days. Hence, we were unable to consider the laxative effects induced by various metabolites of SPA on the disturbance of gut microbiota during our study; future long-term studies are required to confirm whether fermentable saccharides affect the excretion of stool. Meanwhile, some components such as saponin, flavonoids, and polyphenol included in SPA can modulate the composition of gut microbiota through the suppression of pathogen growth and the promotion of beneficial bacteria growth [
38]. These alterations of intestinal microbiota regulated gut sensory and motor functions and then improved chronic constipation. Therefore, the regulation of gut microbiota was considered as one of the new therapeutic tools for the management of chronic constipation [
39]. Also, microbial metabolites short-chain fatty acid (SCFAs) mainly produced in the colon by the bacterial fermentation of carbohydrate can be considered as another regulating factor for laxative effects because SCFAs stimulate the increase of transit time [
40,
41].
A chemically synthesized drug, Lop decreases the risk of dehydration and systemic or metabolic diseases such as diarrhea. Due to these properties, it is widely applied to induce constipation by administering 1.5–3 mg/kg body weight of Lop for 3–7 days in laboratory animals [
35,
36,
37,
42,
43]. Lop stimulates the extension of the stool evacuation time and the delay of intestinal luminal transit by inhibiting water secretion [
44] and smooth muscle movement of the intestinal wall [
45,
46]. However, the efficacy of Lop differs depending on the specific target of their mechanism of action. Diarrhea was controlled for 2 or 8 h following the oral administration of Lop in the castor oil diarrhea test using rats [
47]. In patients with irritable bowel syndrome treated with 4–12 mg of Lop, the prolongation of the whole gut transit lasted from 42 to 56 h [
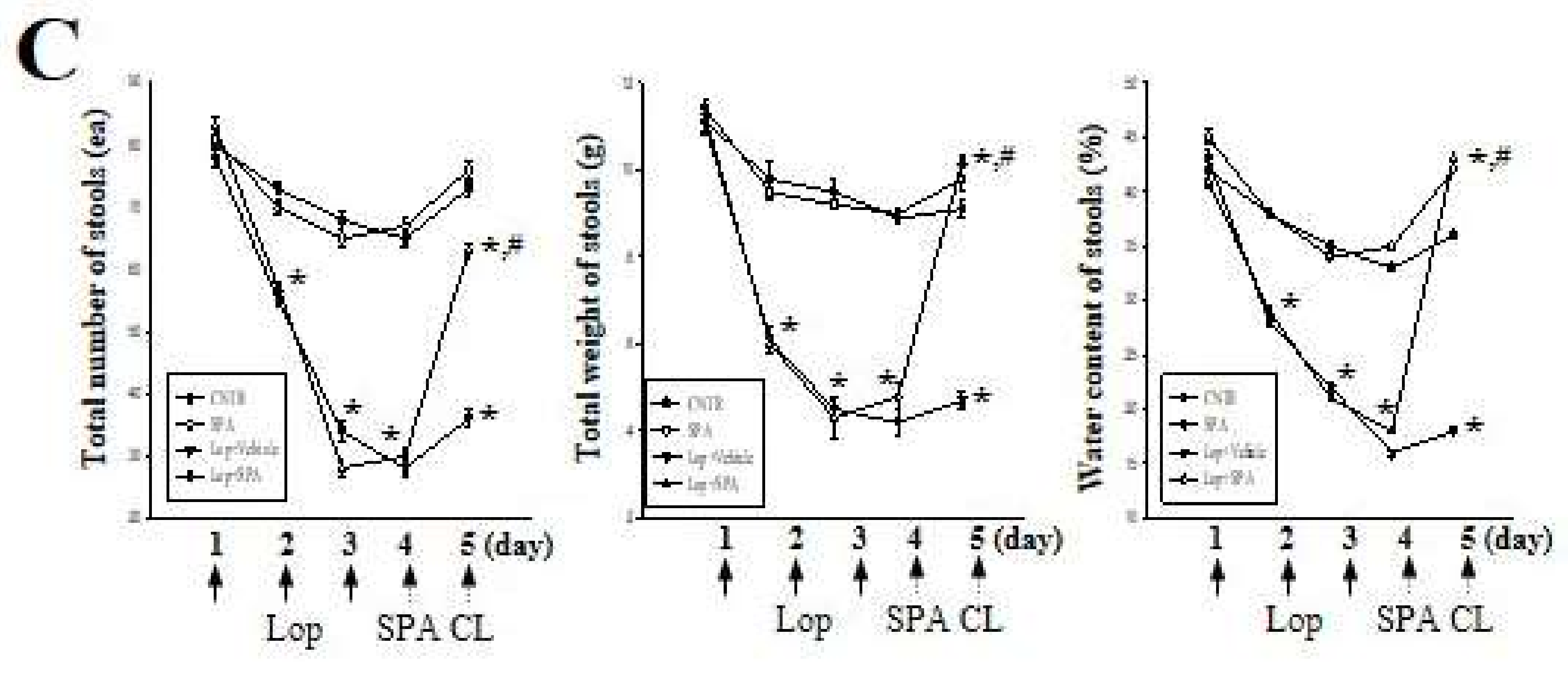
48]. In our study, an alteration of the laxative parameters, including stool excretion, histological structure, signaling pathway of mAChR and mucin secretion, were evaluated in Vehicle- or SPA-treated SD rats at 48 h after the final Lop treatment. The constipation induced by Lop was consistently maintained for 48 h in the Lop + Vehicle-treated group, while the Lop + SPA-treated group showed significant laxative effects. Hence, we believe that the constipation effect of Lop administration on the histological structure of the colon can be completely excluded in SD rats at 48 h after SPA administration.
Food intake and water consumption are important factors to be considered when investigating the process leading to symptoms of constipation and the subsequent therapeutic effects of laxative drugs. In the constipation model, several herbal medicines and foods induce a variety of responses with regards to food intake and water consumption. Aqueous leaf extract of
A. ferox Mill. induces enhanced water consumption without any significant alteration of food intake [
12], while the reverse was observed in a constipation model treated with an aqueous extract of
L. platyphylla (AEtLP) [
5]. Moreover, food intake and water consumption were significantly decreased in the constipation model treated with a
Ficus carica paste [
15]. However, a previous study of GEGR [
7] and the current investigation of SPA reveal no significant alterations in food intake or water consumption. These differences might be associated to unknown factors such as the innate taste of the herbal medicines and foods used in each study.
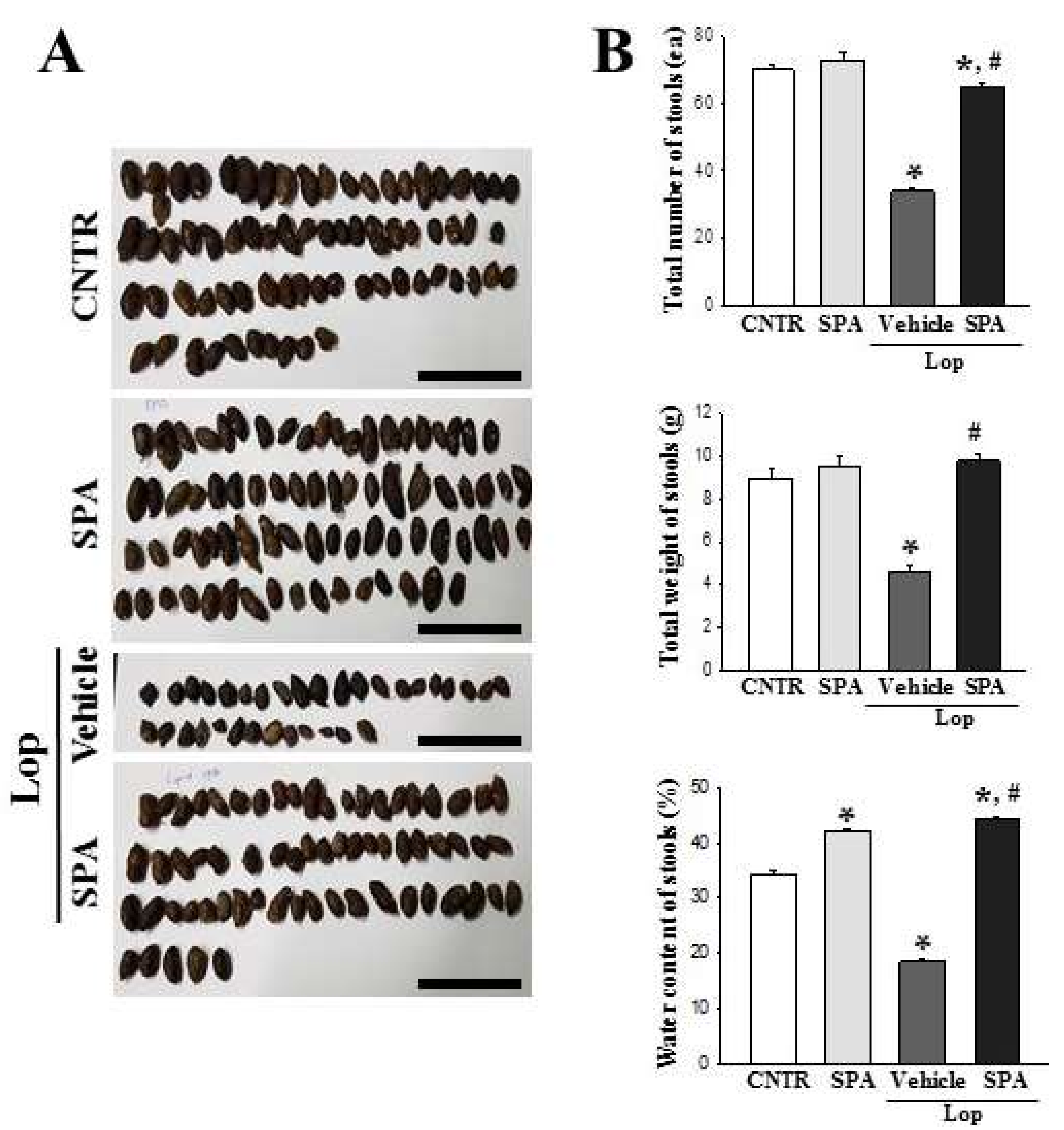
In most constipation studies, a key marker of constipation is the significant reduction in stool excretion in the Lop-induced animals. Previously, stool-related factors including stool number, weight, and water content dramatically decreased in the Lop-induced constipation model [
5,
7,
10,
12,
22]. However, several plant extracts and foods with laxative effects induce the recovery of these alterations. Leaf extracts of
Mareya micrantha and
A. ferox Mill., a widely used medicinal plant with healing properties, are known to improve the number, water content, and weight of stools in Lop-induced rats [
10,
12]. Furthermore, relative to a control group, AEtLP and GEGR increase the stool output of rats exposed to 1000 mg/kg or 250–1000 mg/kg, respectively [
5,
7]. In the current study, a similar effect on stool-related factors was observed in the Lop-induced constipation model treated with 1000 mg/kg of SPA, although some differences were observed in the concentration of the treated extracts.
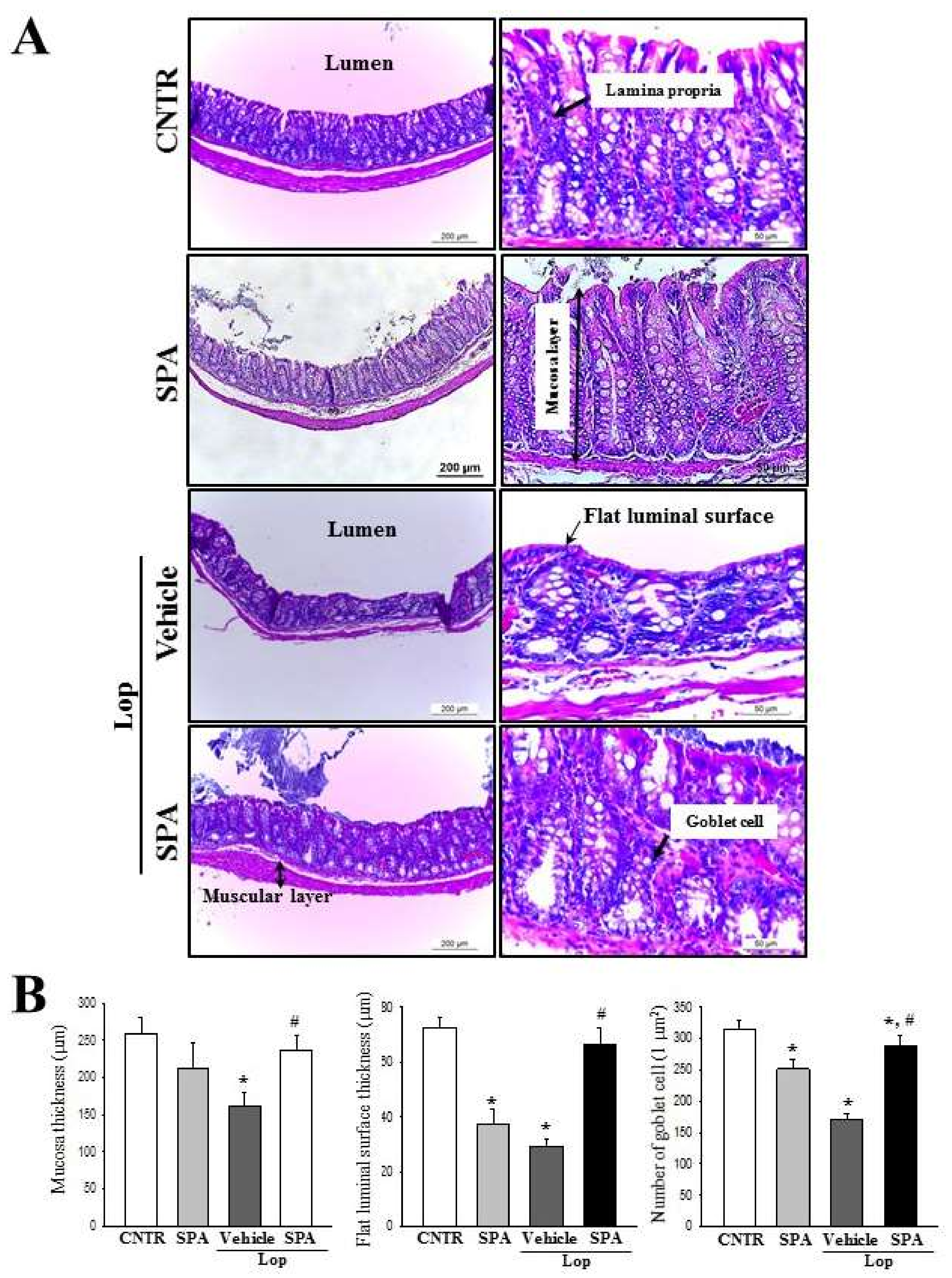
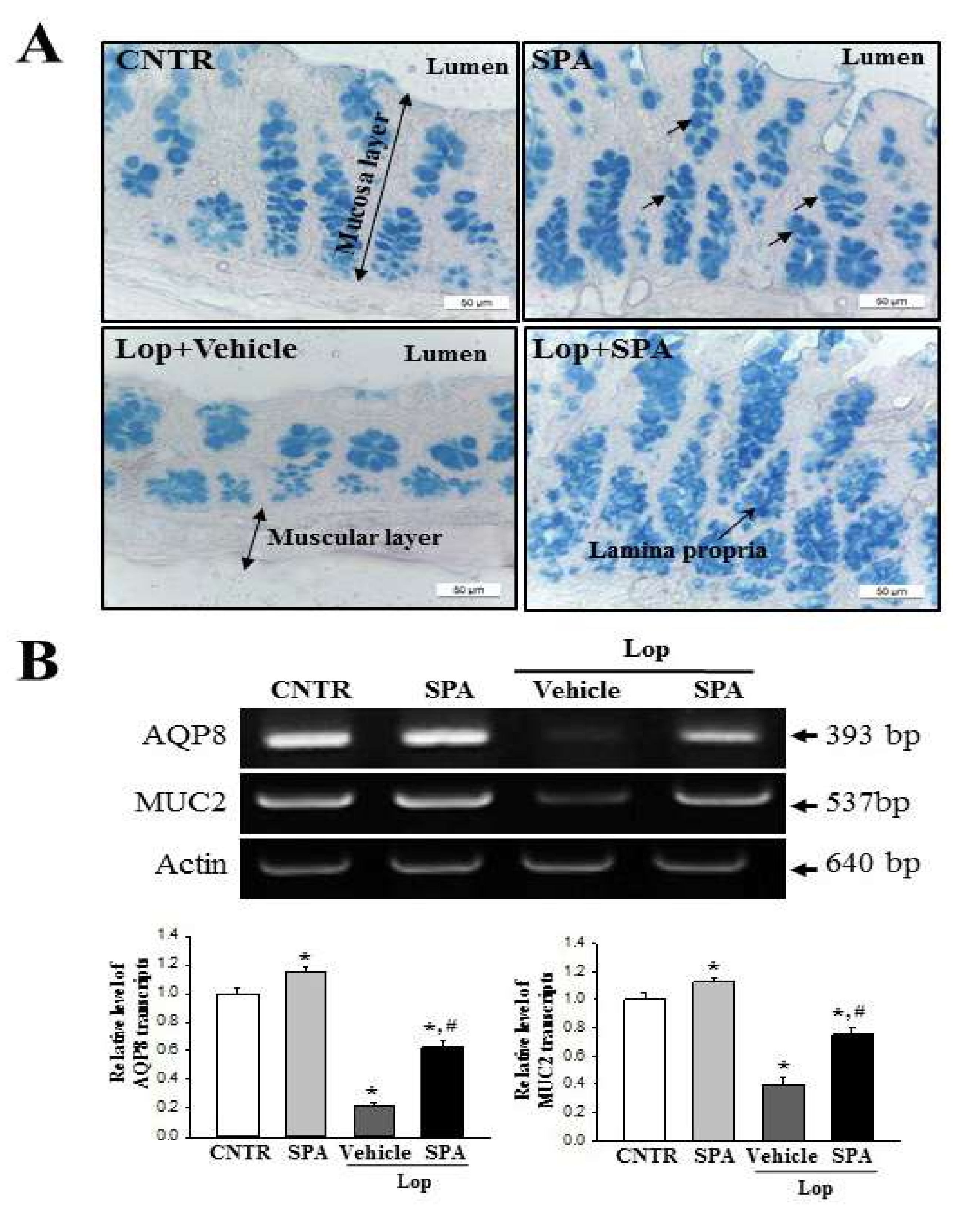
Several significant alterations in histological and cytological structures are detected in the colons of animal models with constipation. The thickness of the colon layer significantly decreases after Lop treatment [
5], and the average thickness of the mucus layer, muscle, and flat luminal surface is thinner in the Lop-treated groups than the vehicle-treated groups [
7]. However, these alterations recovered completely after treatment with several natural products, including AEtLP and GEGR [
5,
7]. As presented in
Figure 3, the histological alterations of the colons observed in the present study were very similar to those of previous studies reported by Kim et al. [
5,
7]. Especially, the flat luminal surface including the outer mucus layer of the colon acts as the first defense line to protect the epithelium from bacteria, inhibiting inflammation and infection [
49]. Their thickness can be altered under various pathological conditions such as ulcerative colitis, Crohn’s disease, and cancer [
50]. Similar patterns were observed in the colon of Lop-induced constipated SD rats. Furthermore, an analysis of the cytological structures revealed similar responses in the crypt of Lieberkühn, including enterocytes, goblet cells, and Paneth cells [
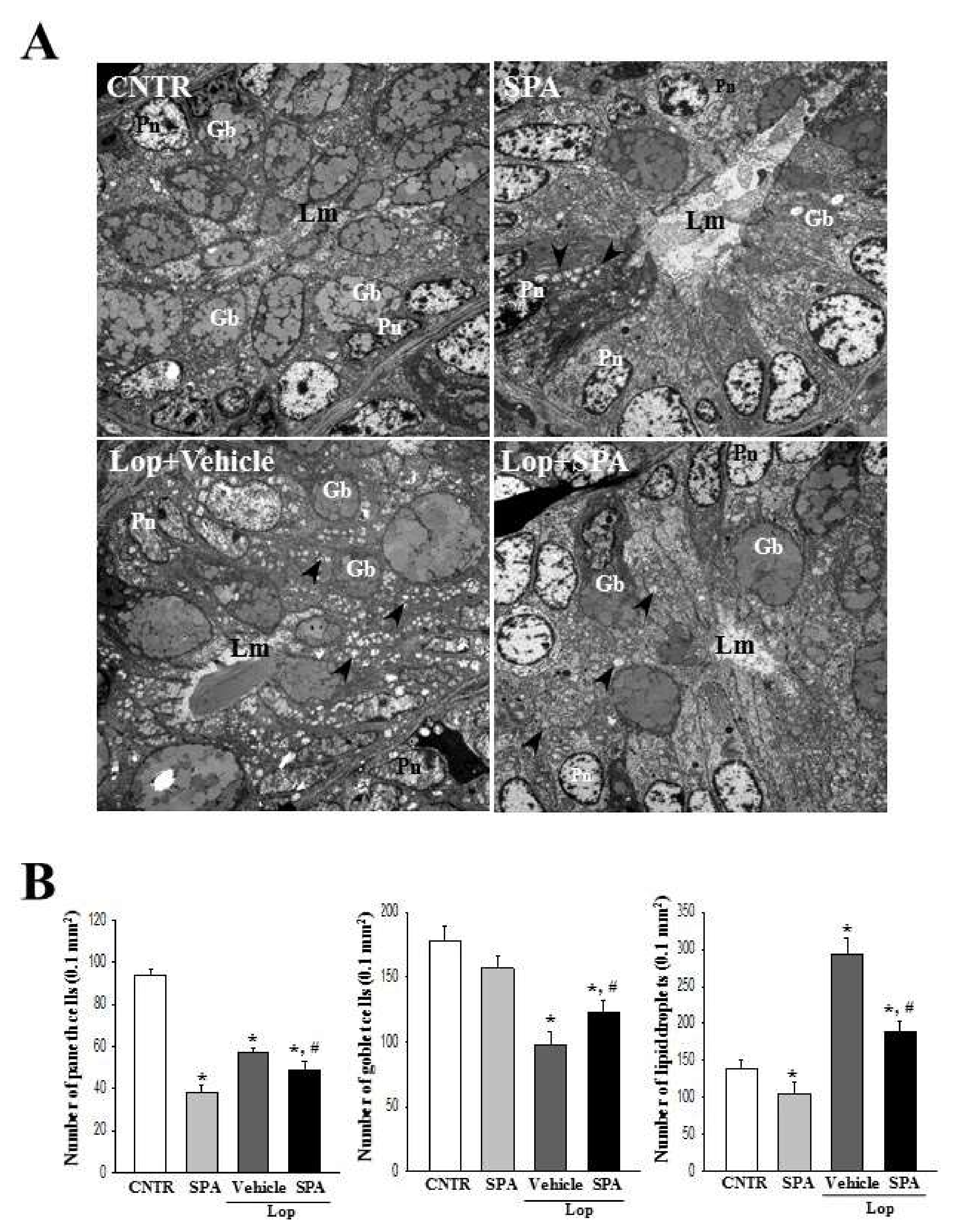
5,
7]. Lipid droplets containing mucin accumulate in the cytoplasm of goblet cells and enterocytes following the induction of constipation, whereas abundant granules were present in paneth cells. However, Lop + SPA administration stimulated the secretion of lipid droplets into the crypt lumen in both goblet cells and enterocytes. These alterations were similarly detected in the Lop-induced constipation model treated with AEtLP and GEGR [
5,
7]. Therefore, our results provide additional evidence that ultrastructural changes in all three types of crypt cells are tightly correlated with the progression of Lop-induced constipation and the recovery with laxative drugs. However, some additional studies including Ussing-like chamber analysis are needed to justify the recovery of mucus thickness in the colon of a Lop-induced constipation model after SPA treatment.
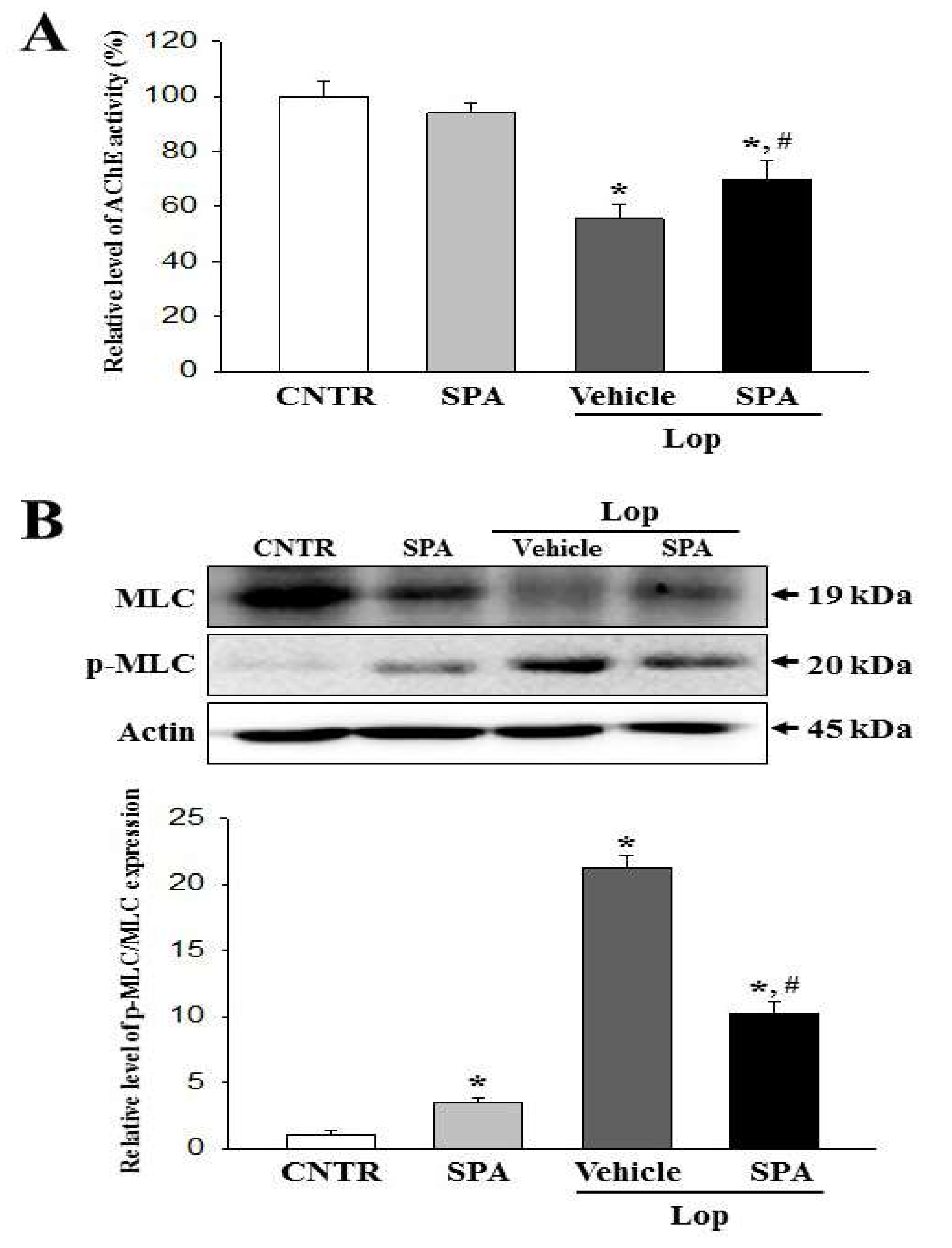
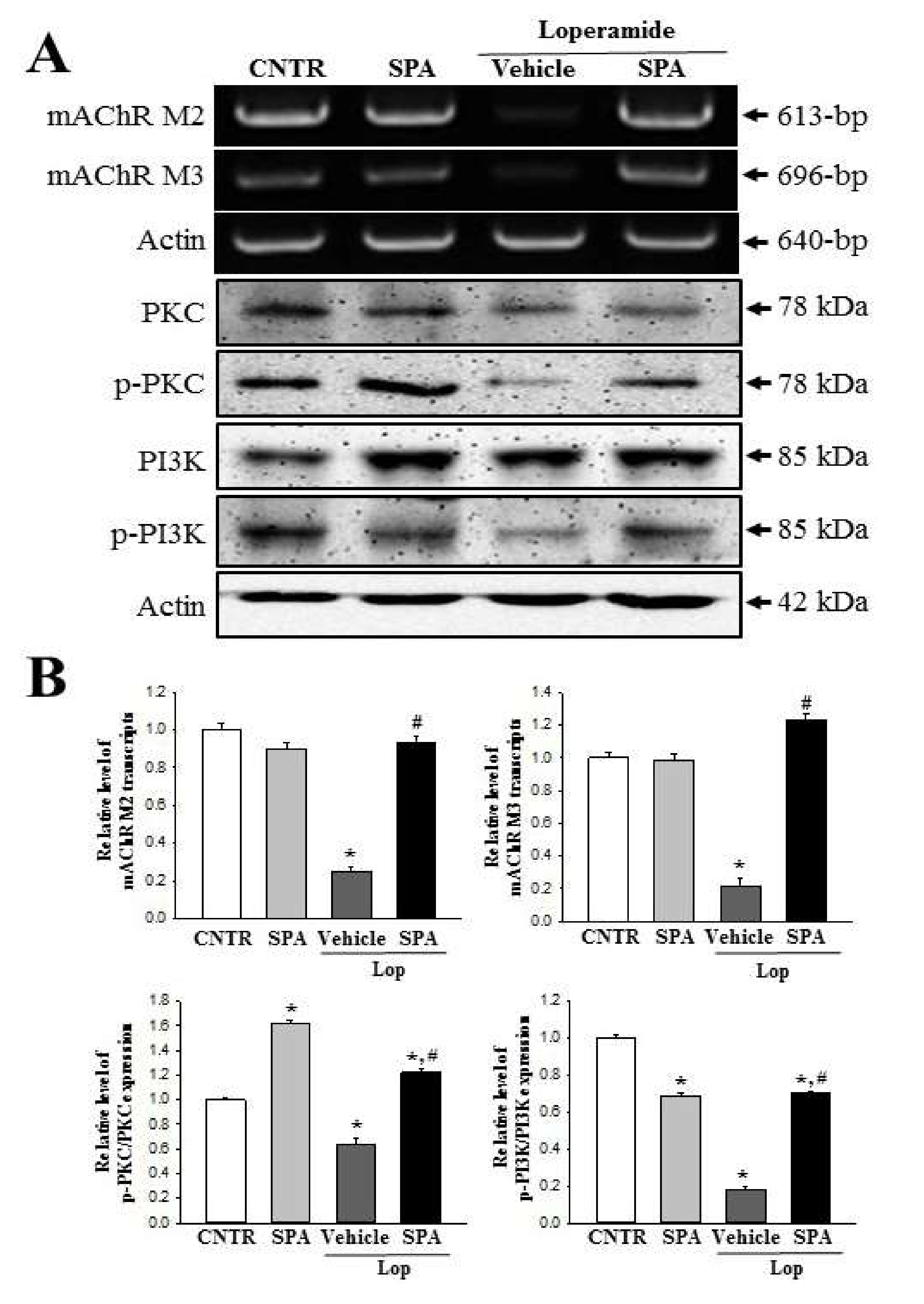
Finally, based on two previous studies, we selected mAChRs as the target protein for the investigation of the molecular mechanism responsible for the laxative effects of SPA [
5,
7]. The results of the previous studies report significant alterations in the levels of the mAChR M2 transcripts in the colon after Lop administration. The levels of mAChR M2 and M3 transcripts dramatically increased in the colon of SD rats during Lop-induced constipation; however, these increases decreased after AEtLP and GEGR treatment. Furthermore, the expression patterns of PI-3K and PKC in the downstream signaling pathway were reported to be very similar to that of mAChR M2 and M3. Also, the increase in PI-3K and PKC expression induced by Lop administration was inhibited by AEtLP and GEGR treatment [
5,
7]. In the current study, we observed similar alterations in the mAChR M2 and M3 signaling pathway in the Lop + SPA-treated group, although there were some variations in their range of alteration. We postulate that the alterations in the mAChR M2 and M3 signaling pathway observed in the present study can provide information for use in future investigations to assess the causes of constipation and to help in the selection of targets for treating constipation-related diseases.
4. Materials and Methods
4.1. Preparation of SPA
We obtained fresh roots of A. cochinchinensis from the Gochang National Agricultural Cooperation Federation in Korea. After washing, the roots obtained were dried in a drying machine (FD5510S-FD5520S, Ilshinbiobase Co., Seoul, Korea) at 60 °C. Voucher specimens of A. cochinchinensis roots (WPC-14-003) were deposited in the functional materials bank of the Wellbeing RIS Center at Pusan National University. The samples were also identified by Shin Woo Cha at the Herbal Crop Research Division, National Institute of Horticultural & Herbal Science. The dried A. cochinchinensis roots were then reduced to a powder using a pulverizer (MF-3100S, Hanil Electric Co., Seoul, Korea), after which SPA was obtained via extraction for 24 h at 50 °C using a fixed liquor ratio (ratio of solid powdered A. cochinchinensis to ethyl acetate solvent, 1:10) in a circulating extraction equipment (SHWB-30/45; Woori Science Instrument Co., Pocheon, South Korea). The SPA was then concentrated and subsequently passed through a 0.4 µm filter, after which the pellets were dried in a rotary evaporator (EYELA, Tokyo, Japan) and stored at −80 °C until used. The collected SPA powder was finally dissolved in distilled water (in dH2O) to 600 mg/kg and diluted further to the required concentration before use.
Meanwhile, SPA contained polyphenols (102.1 μg/g), total flavonoid (88.5 μg/g), and crude saponins (55.7 mg/g). The IC
50 value of SPA on the inhibitory activity of the DPPH radical was determined at 587.6 μg/mL [
33].
4.2. Experimental Design for Animal Study
The Pusan National University-Institutional Animal Care and Use Committee (PNU-IACUC) reviewed and approved the animal protocol for the therapeutic effects of laxative drug based on the ethical procedures for scientific care (Approval Number PNU-2015-0952). The Pusan National University-Laboratory Animal Resources Center, accredited by the Korea Food and Drug Administration (KFDA)(Accredited Unit Number-000231) and The Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) International (Accredited Unit Number; 001525), handled the adult Sprague Dawley (SD) rats, which were purchased from Samtako BioKorea Inc. (Osan, Korea). The animals were provided with access to a standard irradiated chow diet (Samtako BioKorea Inc.) and water ad libitum. Additionally, rats were maintained in a specific pathogen-free (SPF) state under a strict light cycle (on at 08:00 h; off at 20:00 h) at 23 ± 2 °C and 50 ± 10% relative humidity throughout the experiment.
The induction of constipation, the administration schedule of the laxative drug, and the measurement method of several related parameters were performed as per protocols described previously [
7,
14]. The optimal dose of SPA was determined based on our preliminary data which revealed a dose-dependent increase in stool excretion after exposures to 250, 500, and 1000 mg/kg of SPA in SD rats. We selected 1000 mg/kg of SPA as the optimal dosage for the laxative effect because it showed maximal effects without any specific toxicity. In the current study, 8-week-old SD rats (
n = 28) were assigned to either a non-constipation group (CNTR group,
n = 14) or a constipation group (
n = 14). A subcutaneous injection of Lop (4 mg/kg weight) in 0.5% Tween 20 in saline was administered twice daily to induce constipation (9 a.m. and 6 p.m.) for 3 days, whereas the non-constipation group received 0.5% Tween 20 in saline alone under the same pattern. The non-constipation group was further divided into a CNTR group (
n = 7) and a SPA-treated group (
n = 7). At 9 a.m. on the 4th day, the CNTR group did not receive any solution, whereas the SPA-treated group was orally administered a single dose of 1000 mg/kg of SPA. Additionally, the constipation group was divided into a Lop + Vehicle-treated group (
n = 7) and a Lop + SPA-treated group (
n = 7). The Lop + SPA-treated groups were orally administered a single dose of 1000 mg/kg body weight SPA, while the Lop + Vehicle-treated group received the same volume of 1× phosphate buffer saline (PBS) under the same pattern. At 9 a.m. on the 5th day, the total stools, urine, water, and food were collected from the metabolic cage of each group, and the levels were measured using appropriate methods. Moreover, all SD rats were euthanized using CO
2 gas, after which the tissue samples were acquired and stored at −70 °C in Eppendorf tubes until assay.
4.3. Gastrointestinal Transit Ratio Analysis
The gastrointestinal motility was measured by the method described by Choi et al. [
51] and Kim et al. [
52] with some modifications. Briefly, SD rats were fasted for 18 h prior to the experiment but were allowed to consume water
ad libitum. These rats were fed with 1 mL of charcoal meal (3% suspension of activated charcoal in 0.5% aqueous methylcellulose) (Sigma-Aldrich Co., St. Louis, MO, USA). After 30 min of treatment, these rats were euthanized using CO
2 and the intestinal tract was collected from the abdominal cavity. The intestinal charcoal transit ratio was calculated as follows:
4.4. Analysis of Food Intake, Water Intake, and Body Weight
The food weight, water volume, and body weight of SD rats treated with Vehicle or SPA were measured at 9 a.m. on the 5th day using an electrical balance (for food and body weight) and a measuring cylinder (for water volume). The average food intake and water consumption were then calculated using the above data. All measurements were performed three times to ensure the accuracy.
4.5. Measurement of Stool Parameters and Urine Volume
SD rats were individually bred in metabolic cages throughout the experiment to provide uncontaminated stool and urine samples (Daejong Instrument Industry Co., LTD, Seoul, Korea). The stool number and weight were measured as previously described [
5,
12]. Briefly, stools excreted from each SD rat were collected at 9 a.m. from the 1st day to the 5th day, weighed three times each using an electric balance, and counted three times. The stool moisture content was also analyzed as follows:
where A is the weight of fresh stools collected after Lop administration, and B is the weight of stools after drying at 60 °C for 12 h. Furthermore, the urine volume collected at 9 a.m. on the 5th day was measured three times per sample using a cylinder.
4.6. Western Blotting
Pro-Prep Protein Extraction Solution (Intron Biotechnology Inc., Seongnam, Korea) was used to obtain the total proteins from the colons of subset groups (CNTR, SPA-, Lop + Vehicle-, and Lop + SPA-treated SD rats). The acquired proteins were subsequently centrifuged at 13,000 rpm and 4 °C for 5 min, after which the protein concentrations were determined using a SMARTTM Bicinchoninic Acid Protein assay kit (Thermo Fisher Scientific Inc.). The proteins (30 μg) were subjected to 4%–20% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) for 3 h, and the resolved proteins were transferred to nitrocellulose membranes for 2 h at 40 V. The membranes were then probed with the following primary antibodies, overnight at 4 °C: anti-PI-3K (Cell Signaling Technology Inc., Cambridge, MA, USA), anti-p-PI3K (Cell Signaling Technology Inc.), anti-PKC (Cell Signaling Technology Inc.), anti-p-PKC (Cell Signaling Technology Inc.), anti-MLC (1:1000, Abcam, Cambridge, UK), anti-p-MLC (1:1000, Abcam), or anti-actin (Sigma-Aldrich Co.). After washing with a washing buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, and 0.05% Tween 20), the membranes were incubated with 1:1000 diluted horseradish peroxidase-conjugated goat anti-rabbit IgG (Zymed Laboratories, South San Francisco, CA, USA) for 2 h at room temperature, after which the blots were developed using a Chemiluminescence Reagent Plus kit (Pfizer Inc., Gladstone, NJ, USA). Signal images of each protein were subsequently acquired using a digital camera (1.92 MP resolution) of the FluorChem® FC2 Imaging system (Alpha Innotech Corporation, San Leandro, CA, USA). The protein densities were semi-quantified using the AlphaView Program version 3.2.2 (Cell Biosciences Inc., Santa Clara, CA, USA).
4.7. Semiquantitative PCR Anlysis
The total RNA was isolated from the frozen tissue of the colons using RNAzol B solution (Tet-Test Inc.) according to the manufacturer’s protocols. Following the synthesis of cDNA, the genes were amplified by subjecting the samples to 28 cycles of 30 s at 94 °C, 30 s at 62 °C, and 45 s at 72 °C in a Perkin-Elmer Thermal Cycler. The primer sequences used to evaluate various gene expressions were as follows: mAChR M2 sense primer 5′-CCAGT ATCTC CAAGT CTGGT GCAAG G-3′ and antisense primer 5′-GTTCT TGTAA CACAT GAGGA GGTGC-3′; mAChR M3 sense primer 5′-GTCAC TTCTG GTTCA CCACC AAGAG C-3′ and antisense primer 5′-GTGTT CACCA GGACC ATGAT GTTGT AGG-3′; AQP8 sense primer 5′-GTAGT ATGGA CCTAC GTGAG ATCAA GG-3′ and antisense primer 5′-AGAAC CTTTC CTCTG GACTC ACCAC C-3′; MUC2 sense primer 5′-GCTGC TCATT GAGAA GAACG ATGC-3′ and antisense primer 5′-CTCTC CAGGT ACACC ATGTT ACCAG G-3′; and β-actin sense and antisense primers 5′-TGGAA TCCTG TGGCA TCCAT GAAAC-3′ and 5′-TAAAA CGCAG CTCAG TAACA GTCCG-3′, respectively. All PCR products were quantified using 1% agarose gels and a Kodak Electrophoresis Documentation and Analysis System 120.
4.8. Quantitative Real-Time PCR Analysis
Quantitative real-time PCR assessed the relative quantities of mRNA for inflammatory cytokines. The total RNA molecules were isolated from frozen colon tissues using RNA Bee solution (Tet-Test Inc., Friendswood, TX, USA). After the quantification of the RNA concentration, the complement DNA (cDNA) was synthesized using a mixture of oligo-dT primer (Invitrogen, Carlsbad, CA, USA), dNTP, and reverse transcriptase (Superscript II, 18064-014, Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA). Q-PCR was then conducted using a cDNA template and 2×Power SYBR Green (TOYOBO Co., Osaka, Japan) as described in previous studies [
34]. The primer sequences used to evaluate the various gene expressions were as follows: TNF-α sense primer 5′-CCTGT AGCCC ACGTC GTAGC-3′ and antisense primer 5‘-TTGAC CTCAG CGCTG ACTTG-3′; IL-1β sense primer 5′-GCACA TCAAC AAGAG CTTCA GGCAG-3′ and antisense primer 5′-GCTGC TTGTG AGGTG CTGAT GTAC-3′; IL-6 sense primer 5′-TTGGG ACTGA TGTTG TTGACA-3′ and antisense primer 5′-TCATC GCTGT TGATA CAATC AGA-3′; and β-actin sense and antisense primers 5′-TGGAA TCCTG TGGCA TCCAT GAAAC-3′ and 5′-TAAAA CGCAG CTCAG TAACA GTCCG-3′, respectively. The reaction cycle at which PCR products exceeded this fluorescence intensity threshold during the exponential phase of PCR amplification was considered as the threshold cycle (CT).
4.9. Histopathological Analysis
The colons collected from CNTR, SPA-, Lop + Vehicle-, and Lop + SPA-treated SD rats were fixed with 10% formalin for 48 h. The samples were subsequently embedded in paraffin wax, after which they were cut into 4 μm thick sections and stained with hematoxylin and eosin (H&E, Sigma-Aldrich Co.). The sections were then analyzed for mucosa thickness, flat luminal surface thickness, and number of goblet cells in the colons by light microscopy and the Leica Application Suite (Leica Microsystems, Switzerland).
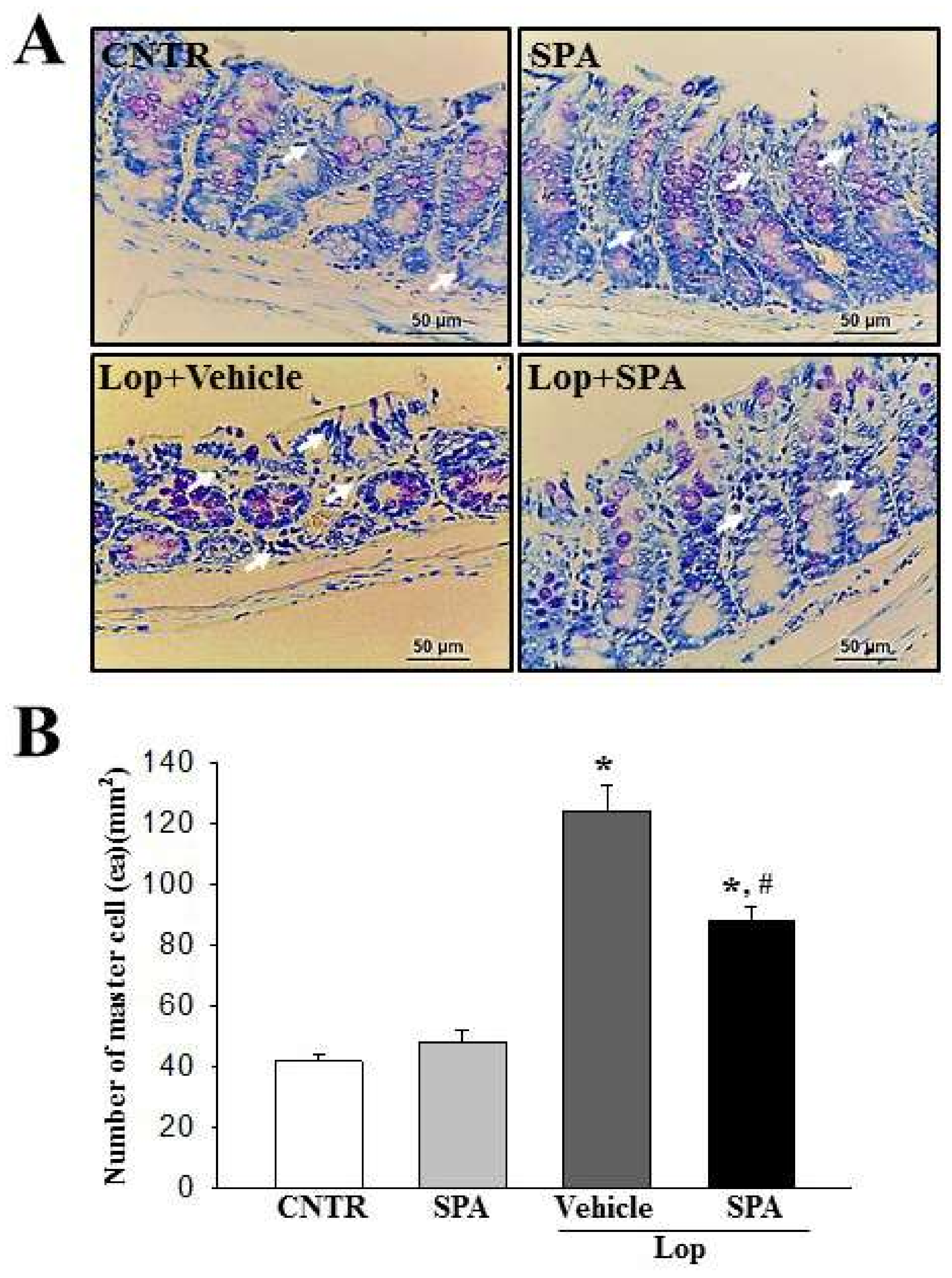
The infiltration of mast cells into the colon was detected by staining with toluidine blue as previously described. After deparaffinization and dehydration, these sections were stained with 0.25% Toluidine blue (Sigma-Aldrich, MO, USA) and measured by light microscopy for the total number of mast cells. The number of cells per specific area was measured using the Leica Application Suite (Leica Microsystems, Wetzlar, Germany).
Mucin staining was conducted by fixing the colons collected from SD rats of all subset groups with 10% formalin for 48 h, then embedding the samples in paraffin wax, and sectioning them into 4 μm thick slices that were subsequently deparaffinized with xylene and rehydrated. The morphological features in the stained colon sections were then observed by light microscopy after the mounted tissue sections were rinsed with distilled water and stained using an Alcian Blue Stain kit (IHC WORLD, Woodstock, MD, USA).
4.10. Ultrastructure Analysis Using Transmission Electron Microscopy (TEM)
Colons collected from CNTR, SPA-, Lop + Vehicle-, and Lop + SPA-treated SD rats were fixed in 2.5% glutaraldehyde in a 1× PBS buffer. Next, the samples were washed, after which they were dehydrated with ascending concentrations of ethanol, incubated in 1% OsO4 for 1 h at room temperature, and finally embedded in Epon812 media (Polysciences Inc., Eppelheim, Germany). The embedded samples were then cut into ultrathin sections (70 nm) that were subsequently collected on holey formvar coated grids, contrasted by treatment with uranyl acetate and lead citrate, and examined by TEM (Hitachi, Tokyo, Japan) at 4000× magnification.
4.11. AChE Activity Analysis
The AChE activity was determined using an Acetylcholinesterase Assay Kit (Abcam, Cambridge, UK) according to the manufacturer’s protocols. Briefly, the colons of each rats was homogenized in a PRO-PREP protein extraction solution (1.0 mM PMSF, 1.0 mM EDTA, 1.0 μM pepstatin, 1.0 μM leupeptin, and 1.0 μM aprotinin) (iNtRON Biotechnology Inc., Seoul, Korea), after which the homogenates were stored at −70 °C until analysis. The sample or standards and the ACh reaction mixture were then incubated on a 96-well plate for 10 min at room temperature while protected from the light. Color alterations were read using a Vmax plate reader (Molecular Devices, Sunnyvale, CA, USA) at 410 nm.
4.12. Statistical Analysis
The statistical significance was evaluated using One-way Analysis of Variance (ANOVA) (SPSS for Windows, Release 10.10, Standard Version, Chicago, IL, USA) followed by Tukey post hoc t-test for multiple comparison. All the values were expressed as the means ± SD, and a p value < 0.05 was considered statistically significant.

 and
and 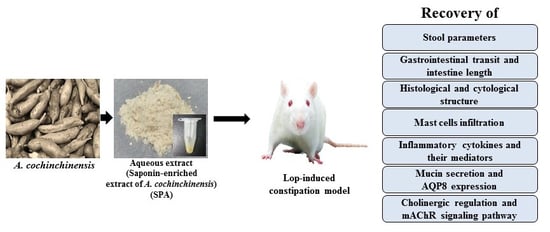











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}