Aggrecan Hypomorphism Compromises Articular Cartilage Biomechanical Properties and Is Associated with Increased Incidence of Spontaneous Osteoarthritis

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Agc1CreERT2/CreERT2 Dwarf Phenotype Persist through the Adulthood
2.2. Agc1CreERT2/CreERT2 Mice Are Characterized by Reduced Aggrecan Expression
2.3. Aggrecan Depletion Results in Stiffening of the Articular Cartilage
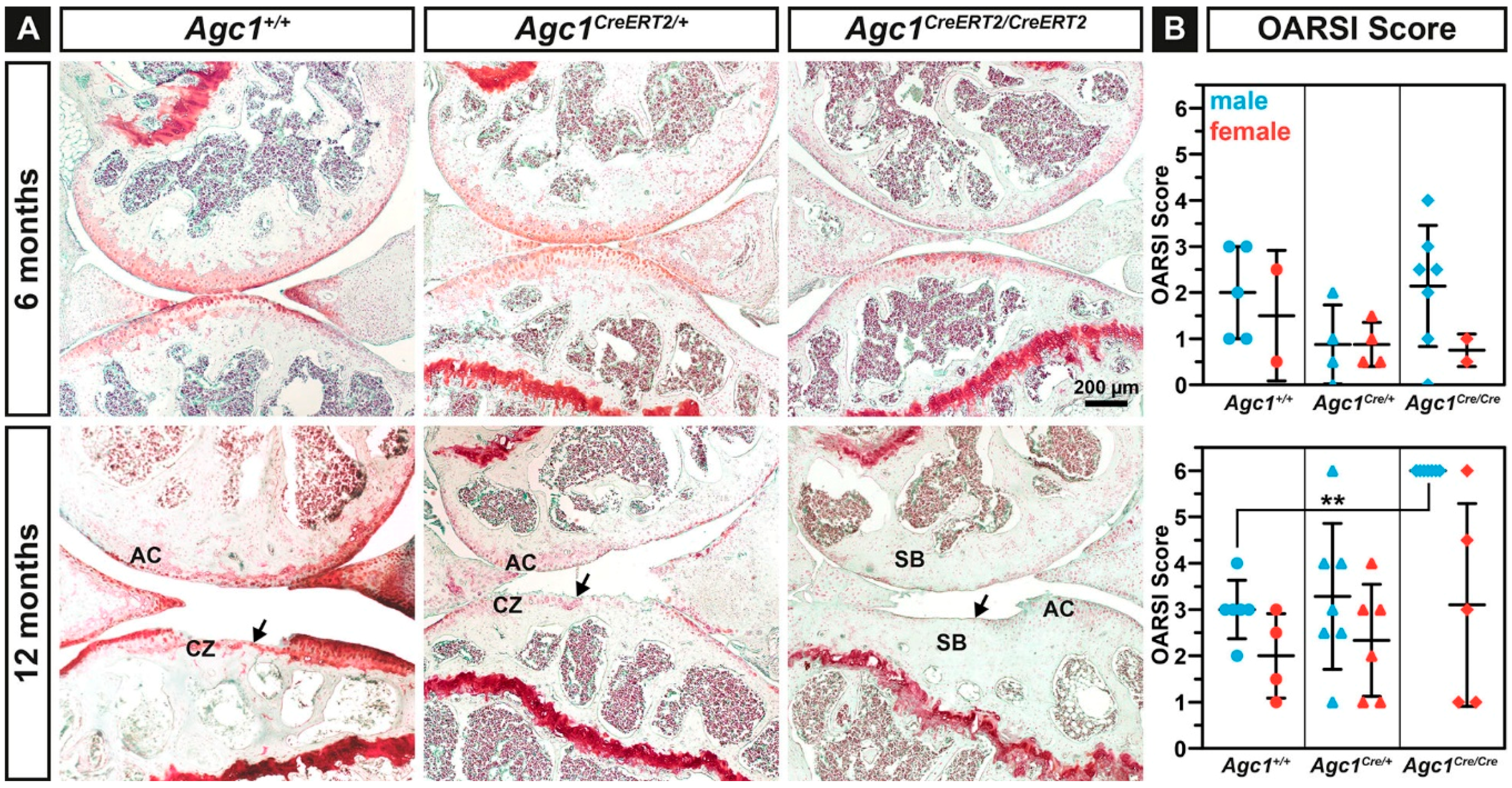
2.4. Agc1CreERT2/CreERT2 Mice Develop Spontaneous Osteoarthritis during Aging
2.5. Higher Incidence of OA in Agc1CreERT2/CreERT2 Mice Is Not Correlated with Increased AC Catabolism
3. Discussion
4. Materials and Methods
4.1. Mouse Model and Husbandry
4.2. Radiography and Morphometry
4.3. Sample Preparation
4.4. Histology
4.5. Scoring of Articular Cartilage Erosion
4.6. Immunohistochemistry
4.7. RNA Isolation and Quantitative RT-PCR
4.8. Atomic Force Microscopy
4.9. Enzyme-Linked Immunosorbent Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aspberg, A. The Different Roles of Aggrecan Interaction Domains. J. Histochem. Cytochem. 2012, 60, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Aspberg, A. Cartilage Proteoglycans. In Cartilage; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–22. [Google Scholar]
- Kiani, C.; Chen, L.; Wu, Y.J.; Yee, A.J.; Yang, B.B. Structure and function of aggrecan. Cell Res. 2002, 12, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedlund, H.; Hedbom, E.; Heinegrd, D.; Mengarelli-Widholm, S.; Reinholt, F.P.; Svensson, O. Association of the aggrecan keratan sulfate-rich region with collagen in bovine articular cartilage. J. Biol. Chem. 1999, 274, 5777–5781. [Google Scholar] [CrossRef] [PubMed]
- Cortes, M.; Baria, A.T.; Schwartz, N.B. Sulfation of chondroitin sulfate proteoglycans is necessary for proper Indian hedgehog signaling in the developing growth plate. Development 2009, 136, 1697–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.G.; Kuffner, T.; Cowan, S.K.; Cheah, K.S.E.; Mason, R.M. Expression of collagen and aggrecan genes in normal and osteoarthritic murine knee joints. Osteoarthr. Cartil. 2002, 10, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroudas, A.; Bayliss, M.T.; Uchitel-Kaushansky, N.; Schneiderman, R.; Gilav, E. Aggrecan turnover in human articular cartilage: Use of aspartic acid racemization as a marker of molecular age. Arch. Biochem. Biophys. 1998, 350, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.G.; Briggs, M.D. The aggrecanopathies; an evolving phenotypic spectrum of human genetic skeletal diseases. Orphanet J. Rare Dis. 2016, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Tompson, S.W.; Merriman, B.; Funari, V.A.; Fresquet, M.; Lachman, R.S.; Rimoin, D.L.; Nelson, S.F.; Briggs, M.D.; Cohn, D.H.; Krakow, D. A Recessive Skeletal Dysplasia, SEMD Aggrecan Type, Results from a Missense Mutation Affecting the C-Type Lectin Domain of Aggrecan. Am. J. Hum. Genet. 2009, 84, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stattin, E.-L.; Tegner, Y.; Domellöf, M.; Dahl, N. Familial osteochondritis dissecans associated with early osteoarthritis and disproportionate short stature. Osteoarthr. Cartil. 2008, 16, 890–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, O.; Guo, M.H.; Dunbar, N.; Popovic, J.; Flynn, D.; Jacobsen, C.; Lui, J.C.; Hirschhorn, J.N.; Baron, J.; Dauber, A. Short stature, accelerated bone maturation, and early growth cessation due to heterozygous aggrecan mutations. J. Clin. Endocrinol. Metab. 2014, 99, E1510–E1518. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Kimata, K.; Line, S.; Strong, D.; Gao, L.Y.; Kozak, C.A.; Yamada, Y. Mouse cartilage matrix deficiency (cmd) caused by a 7 bp deletion in the aggrecan gene. Nat. Genet. 1994, 7, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Juriloff, M.; Harris, M.J. A new mutation at the cmd locus in the mouse. J. Hered. 1986, 77, 205–206. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Nakata, K.; Kimata, K.; Nakanishi, I.; Yamada, Y. Dwarfism and age-associated spinal degeneration of heterozygote cmd mice defective in aggrecan. Proc. Natl. Acad. Sci. USA 1997, 94, 6943–6947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, S.P.; Jang, C.-W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Generation of aggrecan-CreERT2 knockin mice for inducible Cre activity in adult cartilage. Genesis 2009, 47, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Garcia, O.; Matsuzaki, T.; Olmer, M.; Miyata, K.; Mokuda, S.; Sakai, D.; Masuda, K.; Asahara, H.; Lotz, M.K. FOXO are required for intervertebral disk homeostasis during aging and their deficiency promotes disk degeneration. Aging Cell 2018, 17, e12800. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Abu-Amer, Y.; O’Keefe, R.J.; Shen, J. Loss of Dnmt3b in Chondrocytes Leads to Delayed Endochondral Ossification and Fracture Repair. J. Bone Miner. Res. 2018, 33, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Diekman, B.O.; Sessions, G.A.; Collins, J.A.; Knecht, A.K.; Strum, S.L.; Mitin, N.K.; Carlson, C.S.; Loeser, R.F.; Sharpless, N.E. Expression of p16INK4ais a biomarker of chondrocyte aging but does not cause osteoarthritis. Aging Cell 2018, 17, e12771. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, B.; Liu, C.; Serra, R. Tgfbr2 is required in Acan-expressing cells for maintenance of the intervertebral and sternocostal joints. JOR Spine 2018, 1, e1025. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Zhang, S.; Gu, J.; Takarada, T.; Yoneda, Y.; Huang, J.; Zhao, L.; Oh, C.D.; Li, J.; Wang, B.; et al. Deletion of Runx2 in Articular Chondrocytes Decelerates the Progression of DMM-Induced Osteoarthritis in Adult Mice. Sci. Rep. 2017, 7, 2371. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.P.; Liang, S.; Akdemir, K.C.; De Crombrugghe, B. The postnatal role of Sox9 in cartilage. J. Bone Miner. Res. 2012, 27, 2511–2525. [Google Scholar] [CrossRef] [PubMed]
- Couasnay, G.; Bon, N.; Devignes, C.S.; Sourice, S.; Bianchi, A.; Véziers, J.; Weiss, P.; Elefteriou, F.; Provot, S.; Guicheux, J.; et al. PiT1/Slc20a1 Is Required for Endoplasmic Reticulum Homeostasis, Chondrocyte Survival, and Skeletal Development. J. Bone Miner. Res. 2018, 34, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Jiang, H.; Fan, Y.; Lu, R.S.; Wei, C.; Takarada, T.; He, S.; Chen, D. Runx2 is required for postnatal intervertebral disc tissue growth and development. J. Cell. Physiol. 2019, 234, 6679–6687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sheu, T.; Hoak, D.; Shen, J.; Hilton, M.J.; Zuscik, M.J.; Jonason, J.H.; O’Keefe, R.J. CCN1 Regulates Chondrocyte Maturation and Cartilage Development. J. Bone Miner. Res. 2016, 31, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; von der Mark, K.; Henry, S.; Norton, W.; Adams, H.; de Crombrugghe, B. Chondrocytes Transdifferentiate into Osteoblasts in Endochondral Bone during Development, Postnatal Growth and Fracture Healing in Mice. PLoS Genet. 2014, 10, e1004820. [Google Scholar] [CrossRef] [PubMed]
- Rashid, H.; Chen, H.; Hassan, Q.; Javed, A. Dwarfism in homozygous Agc1CreERT mice is associated with decreased expression of aggrecan. Genesis 2017, 55, e23070. [Google Scholar] [CrossRef] [PubMed]
- Loparic, M.; Wirz, D.; Daniels, A.U.; Raiteri, R.; Vanlandingham, M.R.; Guex, G.; Martin, I.; Aebi, U.; Stolz, M. Micro- and nanomechanical analysis of articular cartilage by indentation-type atomic force microscopy: Validation with a gel-microfiber composite. Biophys. J. 2010, 98, 2731–2740. [Google Scholar] [CrossRef] [PubMed]
- Prein, C.; Warmbold, N.; Farkas, Z.; Schieker, M.; Aszodi, A.; Clausen-Schaumann, H. Structural and mechanical properties of the proliferative zone of the developing murine growth plate cartilage assessed by atomic force microscopy. Matrix Biol. 2016, 50, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gronau, T.; Krüger, K.; Prein, C.; Aszodi, A.; Gronau, I.; Iozzo, R.V.; Mooren, F.C.; Clausen-Schaumann, H.; Bertrand, J.; Pap, T.; et al. Forced exercise-induced osteoarthritis is attenuated in mice lacking the small leucine-rich proteoglycan decorin. Ann. Rheum. Dis. 2017, 76, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Hauer, N.N.; Sticht, H.; Boppudi, S.; Büttner, C.; Kraus, C.; Trautmann, U.; Zenker, M.; Zweier, C.; Wiesener, A.; Jamra, R.A.; et al. Genetic screening confirms heterozygous mutations in ACAN as a major cause of idiopathic short stature. Sci. Rep. 2017, 7, 12225. [Google Scholar] [CrossRef] [PubMed]
- Gleghorn, L.; Ramesar, R.; Beighton, P.; Wallis, G. A mutation in the variable repeat region of the aggrecan gene (AGC1) causes a form of spondyloepiphyseal dysplasia associated with severe, premature osteoarthritis. Am. J. Hum. Genet. 2005, 77, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Gkourogianni, A.; Andrew, M.; Tyzinski, L.; Crocker, M.; Douglas, J.; Dunbar, N.; Fairchild, J.; Funari, M.F.A.; Heath, K.E.; Jorge, A.A.L.; et al. Clinical Characterization of Patients With Autosomal Dominant Short Stature due to Aggrecan Mutations. J. Clin. Endocrinol. Metab. 2017, 102, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Primorac, D.; Stover, M.L.; Clark, S.H.; Rowe, D.W. Molecular basis of nanomelia, a heritable chondrodystrophy of chicken. Matrix Biol. 1994, 14, 297–305. [Google Scholar] [CrossRef]
- Vertel, B.M.; Grier, B.L.; Li, H.; Schwartz, N.B. The chondrodystrophy, nanomelia: Biosynthesis and processing of the defective aggrecan precursor. Biochem. J. 1994, 301, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, J.A.L.; Tammen, I.; Windsor, P.A.; Bateman, J.F.; Savarirayan, R.; Nicholas, F.W.; Raadsma, H.W. Bulldog dwarfism in Dexter cattle is caused by mutations in ACAN. Mamm. Genome 2007, 18, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Lauing, K.L.; Cortes, M.; Domowicz, M.S.; Henry, J.G.; Baria, A.T.; Schwartz, N.B. Aggrecan is required for growth plate cytoarchitecture and differentiation. Dev. Biol. 2014, 396, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritsch, A.; Loeckermann, S.; Kern, J.S.; Braun, A.; Bösl, M.R.; Bley, T.A.; Schumann, H.; von Elverfeldt, D.; Paul, D.; Erlacher, M.; et al. A hypomorphic mouse model of dystrophic epidermolysis bullosa reveals mechanisms of disease and response to fibroblast therapy. J. Clin. Investig. 2008, 118, 1669–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohn, A.R.; Gainetdinov, R.R.; Caron, M.G.; Koller, B.H. Mice with reduced NMDA receptor expression display behaviors related to schizophrenia. Cell 1999, 98, 427–436. [Google Scholar] [CrossRef]
- Nagy, A.; Moens, C.; Ivanyi, E.; Pawling, J.; Gertsenstein, M.; Hadjantonakis, A.K.; Pirity, M.; Rossant, J. Dissecting the role of N-myc in development using a single targeting vector to generate a series of alleles. Curr. Biol. 1998, 8, 661–664. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, L.; Liu, Q.; Li, Y.; Guo, H.; Peng, Y.; Peng, H.; Tang, B.; Hu, Z.; Zhao, J.; et al. Insertion of a knockout-first cassette in Ampd1 gene leads to neonatal death by disruption of neighboring genes expression. Sci. Rep. 2016, 6, 35970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, C.T.; MacIvor, D.M.; Hug, B.A.; Heusel, J.W.; Ley, T.J. Long-range disruption of gene expression by a selectable marker cassette. Proc. Natl. Acad. Sci. USA 1996, 93, 13090–13095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffai, R.L.; Weisgraber, K.H. Hypomorphic apolipoprotein E mice: A new model of conditional gene repair to examine apolipoprotein E-mediated metabolism. J. Biol. Chem. 2002, 277, 11064–11068. [Google Scholar] [CrossRef] [PubMed]
- Stolz, M.; Raiteri, R.; Daniels, A.U.; VanLandingham, M.R.; Baschong, W.; Aebi, U. Dynamic elastic modulus of porcine articular cartilage determined at two different levels of tissue organization by indentation-type atomic force microscopy. Biophys. J. 2004, 86, 3269–3283. [Google Scholar] [CrossRef]
- Stolz, M.; Gottardi, R.; Raiteri, R.; Miot, S.; Martin, I.; Imer, R.; Staufer, U.; Raducanu, A.; Düggelin, M.; Baschong, W.; et al. Early detection of aging cartilage and osteoarthritis in mice and patient samples using atomic force microscopy. Nat. Nanotechnol. 2009, 4, 186–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aro, E.; Salo, A.M.; Khatri, R.; Finnilä, M.; Miinalainen, I.; Sormunen, R.; Pakkanen, O.; Holster, T.; Soininen, R.; Prein, C.; et al. Severe Extracellular Matrix Abnormalities and Chondrodysplasia in Mice Lacking Collagen Prolyl 4-Hydroxylase Isoenzyme II in Combination with a Reduced Amount of Isoenzyme I. J. Biol. Chem. 2015, 290, 16964–16978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamper, M.; Hamann, N.; Prein, C.; Clausen-Schaumann, H.; Farkas, Z.; Aszodi, A.; Niehoff, A.; Paulsson, M.; Zaucke, F. Early changes in morphology, bone mineral density and matrix composition of vertebrae lead to disc degeneration in aged collagen IX -/- mice. Matrix Biol. 2016, 49, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.-Y.; Wu, C.-B.; Tang, B.; Wang, T.; Yan, C.-H.; Lu, W.W.; Pan, H.; Hu, Y.; Chiu, K.-Y. Collagen fibril stiffening in osteoarthritic cartilage of human beings revealed by atomic force microscopy. Osteoarthr. Cartil. 2012, 20, 916–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleemann, R.U.; Krocker, D.; Cedraro, A.; Tuischer, J.; Duda, G.N. Altered cartilage mechanics and histology in knee osteoarthritis: Relation to clinical assessment (ICRS Grade). Osteoarthr. Cartil. 2005, 13, 958–963. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, R.E.; Zauscher, S.; Guilak, F. Micromechanical mapping of early osteoarthritic changes in the pericellular matrix of human articular cartilage. Osteoarthr. Cartil. 2013, 21, 1895–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, M.I. Sex differences in osteoarthritis of the hip and knee. J. Am. Acad. Orthop. Surg. 2007, 15, S22–S25. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, C.; Nguyen, C.; Lefevre-Colau, M.-M.; Rannou, F.; Poiraudeau, S. Risk factors and burden of osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Sokoloff, L. Natural history of degenerative joint disease in small laboratory animals. I. Pathological anatomy of degenerative joint disease in mice. AMA Arch. Pathol. 1956, 62, 118–128. [Google Scholar] [PubMed]
- van Osch, G.J.; van der Kraan, P.M.; Vitters, E.L.; Blankevoort, L.; van den Berg, W.B. Induction of osteoarthritis by intra-articular injection of collagenase in mice. Strain and sex related differences. Osteoarthr. Cartil. 1993, 1, 171–177. [Google Scholar] [CrossRef]
- Ma, H.-L.; Blanchet, T.J.; Peluso, D.; Hopkins, B.; Morris, E.A.; Glasson, S.S. Osteoarthritis severity is sex dependent in a surgical mouse model. Osteoarthr. Cartil. 2007, 15, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raducanu, A.; Hunziker, E.B.; Drosse, I.; Aszodi, A. Β1 Integrin Deficiency Results in Multiple Abnormalities of the Knee Joint. J. Biol. Chem. 2009, 284, 23780–23792. [Google Scholar] [CrossRef] [PubMed]
- Glasson, S.S.; Chambers, M.G.; Van Den Berg, W.B.; Little, C.B. The OARSI histopathology initiative—Recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18, S17–S23. [Google Scholar] [CrossRef] [PubMed]
- Butt, H.-J.; Jaschke, M. Calculation of thermal noise in atomic force microscopy. Nanotechnology 1995, 6. [Google Scholar] [CrossRef]
- Sneddon, I.N. The relation between load and penetration in the axisymmetric boussinesq problem for a punch of arbitrary profile. Int. J. Eng. Sci. 1965, 3, 47–57. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberton, P.; Dugonitsch, H.C.; Hartmann, B.; Li, P.; Farkas, Z.; Saller, M.M.; Clausen-Schaumann, H.; Aszodi, A. Aggrecan Hypomorphism Compromises Articular Cartilage Biomechanical Properties and Is Associated with Increased Incidence of Spontaneous Osteoarthritis. Int. J. Mol. Sci. 2019, 20, 1008. https://doi.org/10.3390/ijms20051008
Alberton P, Dugonitsch HC, Hartmann B, Li P, Farkas Z, Saller MM, Clausen-Schaumann H, Aszodi A. Aggrecan Hypomorphism Compromises Articular Cartilage Biomechanical Properties and Is Associated with Increased Incidence of Spontaneous Osteoarthritis. International Journal of Molecular Sciences. 2019; 20(5):1008. https://doi.org/10.3390/ijms20051008
Chicago/Turabian StyleAlberton, Paolo, Hans Christian Dugonitsch, Bastian Hartmann, Ping Li, Zsuzsanna Farkas, Maximilian Michael Saller, Hauke Clausen-Schaumann, and Attila Aszodi. 2019. "Aggrecan Hypomorphism Compromises Articular Cartilage Biomechanical Properties and Is Associated with Increased Incidence of Spontaneous Osteoarthritis" International Journal of Molecular Sciences 20, no. 5: 1008. https://doi.org/10.3390/ijms20051008