Treatment of Cells and Tissues with Chromate Maximizes Mitochondrial 2Fe2S EPR Signals


 ,
,
Abstract
:
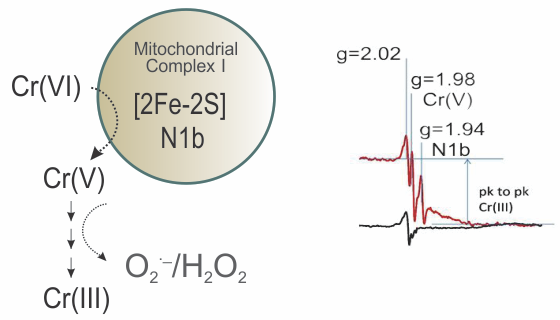{kind=link}
{kind=link}
{kind=link}
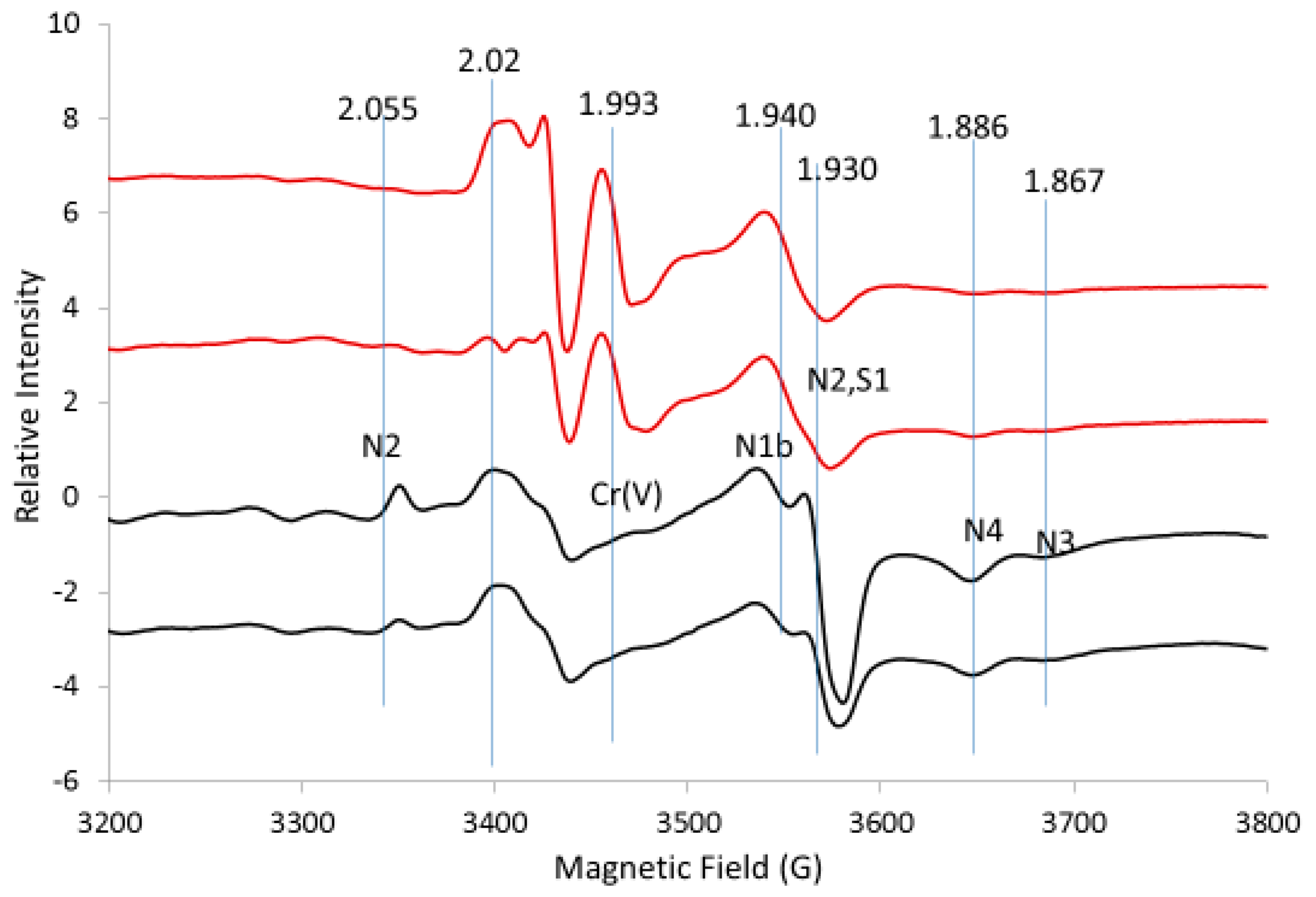{kind=link}
{kind=link}
{kind=link}
{kind=link}
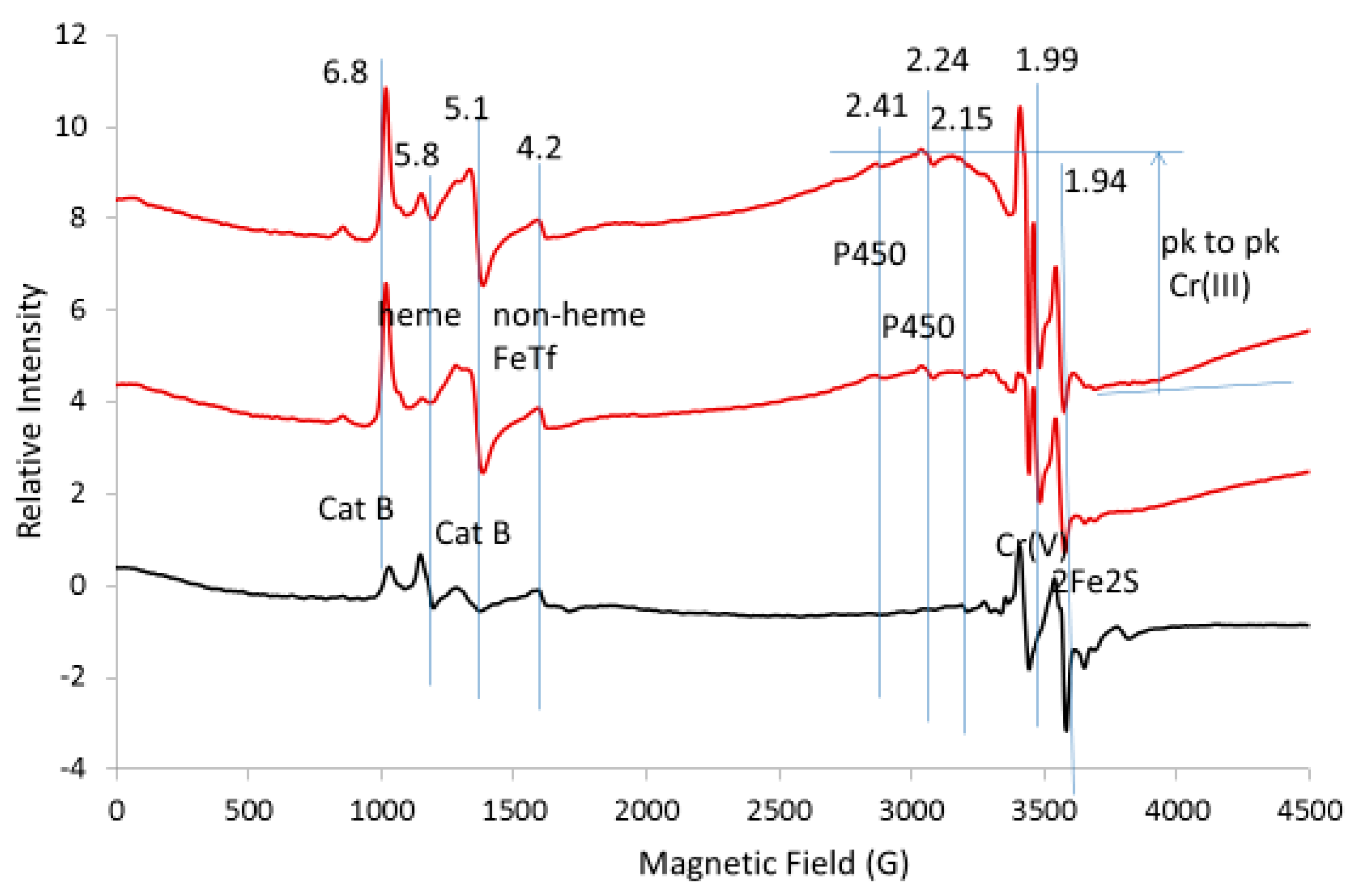{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Increase of the g = 1.94 2Fe2S Signal Upon Addition of Chromate to White Blood Cells
2.2. Chromate Added to Liver Cells
2.2.1. Chromate Does Not Oxidize the 2Fe2S Sites in Liver Tissue
2.2.2. Temperature Dependence for EPR Spectra from Liver Tissue Treated with Chromate
2.2.3. Increase of low-field EPR Lines Due to Oxidation by Chromate
2.2.4. Determining the Concentration of the Mitochondrial 2Fe2S Cluster in Liver Tissue
2.3. Use of the 2Fe2S Signal Obtained from Addition of Chromate to Melanoma Cells as An Internal Control
3. Discussion
4. Materials and Methods
4.1. Sodium Chromate
4.2. Liver Tissue
4.3. White Blood Cells
4.4. Melanoma Cells, Virus Infection, Western Blotting
4.5. EPR Equipment
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EPR | Electron paramagnetic resonance |
| Fe-S | Iron-sulfur cluster |
| PBS | Phosphate buffered saline |
| 2Fe2S | Two-iron two-sulfur cluster |
| 3Fe4S | Three-iron four-sulfur cluster |
| 4Fe4S | Four-iron four-sulfur cluster |
References
- Myers, C.R.; Antholine, W.E.; Myers, J.M. The pro-oxidant chromium(VI) inhibits mitochondrial complex I, complex II and aconitase in the bronchial epithelium: EPR markers for Fe-S proteins. Free Radic. Biol. Med. 2010, 49, 1903–1915. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T. Iron-sulfur clusters/semiquinones in complex I. Biochim. Biophys. Acta 1998, 1364, 186–206. [Google Scholar] [CrossRef]
- Myers, J.M.; Antholine, W.E.; Myers, C.R. Hexavalent chromium causes the oxidation of thioredoxin in human bronchial epithelial cells. Toxicology 2008, 246, 222–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.M.; Myers, C.R. The effects of hexavalent chromium on thioredoxin reductase and peroxiredoxins in human bronchial epithelial cells. Free Radic. Biol. Med. 2009, 47, 1477–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.M.; Antholine, W.E.; Myers, C.R. The intracellular redox stress caused by hexavalent chromium is selective for proteins that have key roles in cell survival and thiol redox control. Toxicology 2011, 281, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulteau, A.-L.; Lundberg, K.C.; Ikeda-Saito, M.; Isaye, G.; Szweda, L.I. Reversible redox-dependent modulation of mitochondrial aconitase and proteolytic activity during in vivo cardiac ischemia/reperfusion. Proc. Natl. Acad. Sci. USA 2005, 102, 5987–5991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudder, B.N.; Morales, J.G.; Stubna, A.; Munck, E.; Hendrich, M.P.; Lindahl, P.A. Electron paramagnetic resonance and Mössbauer spectroscopy of intact mitochondria from respiring Saccharomyces cerevisiae. J. Biol. Inorg. Chem. 2007, 12, 1029–1053. [Google Scholar] [CrossRef] [PubMed]
- Svistunenko, D.A.; Davies, N.; Brealy, D.; Singer, M.; Cooper, C.E. Mitochondrial dysfunction in patients with severe sepsis: an EPR interrogation of individual respiratory chain components. Biochim. Biophys. 2006, 1757, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Bennett, B.; Helbling, D.; Meng, H.; Jarzembowski, J.; Geurts, A.M.; Friederich, M.W.; Van Hove, J.L.K.; Lawlor, M.W.; Dimmock, D.P. Potentially diagnostic electron paramagnetic spectra elucidate the underlying mechanism of mitochondrial dysfunction in deoxyguanosine kinase deficient rat model of a genetic mitochondrial DNA depletion syndrome. Free Radic. Biol. Med. 2016, 93, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Reda, T.; Barker, C.D.; Hirst, J. Reduction of the iron-sulfur clusters in mitochondrial NADH:ubiquinone oxidoreductase (complex I) by EuII-DTPA, a very low potential reductant. Biochemistry 2008, 47, 8885–8893. [Google Scholar] [CrossRef] [PubMed]
- Shergill, J.K.; Cammack, R.; Chen, J.-H.; Fisher, M.J.; Madden, S.; Rees, H.H. EPR spectroscopic characterization of the iron-sulphur proteins and cytochrome P-450 in mitochondria from the insect Spodoptera littoralis (cotton leafworm). Biochem. J. 1995, 307, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Borthiry, G.R.; Antholine, W.E.; Myers, J.M.; Myers, C.R. Reductive activation of hexavalent chromium by human lung epithelial cells: generation of Cr(V) and Cr(V)-thiol species. J. Inorg. Biochem. 2008, 102, 1449–1462. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Ohnishi, S.T.; Shinzawa-Itoh, K.; Yoshikawa, S.; Weber, R.T. EPR detection of two protein-associated ubiquinone components (SQ(Nf) and SQ(Ns)) in the membrane in situ and in proteoliposomes of isolated bovine heart complex I. Biochim. Biophys. Acta 2012, 1817, 1803–1809. [Google Scholar] [CrossRef] [PubMed]
- Roessler, M.M.; King, M.S.; Robinson, A.J.; Harmer, J.; Hirst, J. Direct assignment of EPR spectra to structurally defined iron-sulfur clusters in complex I by double electron-electron resonance. Proc. Natl. Acad. Sci. USA 2010, 107, 1930–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krepkiy, D.; Antholine, W.E.; Myers, C.; Petering, D.H. Model reactions of Cr(VI) with DNA mediated by thiol species. Mol. Cell. Biochem. 2001, 222, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Borthiry, G.R.; Antholine, W.E.; Kalyanaraman, B.; Myers, J.M.; Myers, C.R. Reduction of hexavalent chromium by human cytochrome b5: Generation of hydroxyl radical and superoxide. Free Radic. Biol. Med. 2007, 42, 735–737. [Google Scholar] [CrossRef] [PubMed]
- Abdrakhmanova, A.; Dobrynin, K.; Zwicker, K.; Kerscher, S.; Brandt, U. Functional sulfurtransferase is associated with mitochondrial complex I from Yarrowia lipolytica but is not required for assembly of its iron-sulfur clusters. FEBS Lett. 2005, 579, 6781–6785. [Google Scholar] [CrossRef] [PubMed]
- Beinert, H.; Sands, R.H. Studies on succinic and DPNH dehydrogenase preparations by paramagnetic resonance (EPR) spectroscopy. Biochem. Biophys. Res. Commun. 1960, 3, 41–46. [Google Scholar] [CrossRef]
- Salerno, J.C.; Lim, J.; King, T.E.; Blum, H.; Ohnishi, T.J. The spatial relationship and structure of the binuclear iron-sulfur clusters in succinate dehydrogenase. Biol. Chem. 1979, 254, 4828–4835. [Google Scholar]
- Edmondson, D.; Ballou, D.; Van Heuvelen, A.; Palmer, G.; Massey, V. Kinetic studies on the substrate reduction of xanthine oxidase. J. Biol. Chem. 1973, 248, 6135–6144. [Google Scholar] [PubMed]
- Toshio Iwasaki Group Homepage. Available online: www2.nms.ac.jp/fesworld/LigandMutagenesis.html (accessed on 10 June 2016).
- Johnson, M.K.; Morningstar, J.E.; Bennett, D.E.; Ackrell, B.A.; Kearney, E.B. Magnetic circular dichroism studies of succinate dehydrogenase. J. Biol. Chem. 1985, 260, 7368–7378. [Google Scholar] [PubMed]
- Xing, G.; Diao, Y.; Hoffart, L.M.; Barr, E.W.; Prabhu, K.S.; Arner, R.J.; Reddy, C.C.; Kregs, C.; Bollinger, J.M. Evidence for C–H cleavage by an iron–superoxide complex in the glycol cleavage reaction catalyzed by myo-inositol oxygenase. Proc. Natl. Acad. Sci. USA 2006, 103, 6130–6135. [Google Scholar] [CrossRef] [PubMed]
- United Mitochondrial Disease Foundation. Available online: https://www.umdf.org/types/ (accessed on 12 December 2018).
- Stowe, D.F.; Camara, A.K. Mitochondrial reactive oxygen species production in excitable cells: modulators of mitochondrial and cell function. Antioxid. Redox Signal. 2009, 11, 1373–1414. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-15400-0. [Google Scholar]
- Hong, S.-K.; Yoon, S.; Moelling, C.; Arthan, D.; Park, J.-I. Noncatalytic function of ERK1/2 can promote Raf/MEK/ERK-mediated growth arrest signaling. J. Biol. Chem. 2009, 284, 33006–33018. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.-K.; Hong, S.-K.; Veeranki, S.; Karkhanis, M.; Starenki, D.; Plaza, J.A.; Park, J.-I. A mortalin/HSPA9-mediated switch in tumor-suppressive signaling of Raf/MEK/extracellular signal-regulated kinase. Mol. Cell. Biol. 2013, 33, 4051–4067. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.-K.; Hong, S.-K.; Park, J.-I. Steady-state levels of phosphorylated mitogen-activated protein kinase kinase 1/2 determined by mortalin/HSPA9 and protein phosphatase 1 alpha in KRAS and BRAF tumor cells. Mol. Cell. Biol. 2017, 37, e00061-17. [Google Scholar] [CrossRef] [PubMed]
- Stoll, S.; Schweiger, A.J. EasySpin, a comprehensive software package for spectral simulation and analysis in EPR. J. Magn. Reson. 2006, 178, 42. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antholine, W.E.; Vasquez-Vivar, J.; Quirk, B.J.; Whelan, H.T.; Wu, P.K.; Park, J.-I.; Myers, C.R. Treatment of Cells and Tissues with Chromate Maximizes Mitochondrial 2Fe2S EPR Signals. Int. J. Mol. Sci. 2019, 20, 1143. https://doi.org/10.3390/ijms20051143
Antholine WE, Vasquez-Vivar J, Quirk BJ, Whelan HT, Wu PK, Park J-I, Myers CR. Treatment of Cells and Tissues with Chromate Maximizes Mitochondrial 2Fe2S EPR Signals. International Journal of Molecular Sciences. 2019; 20(5):1143. https://doi.org/10.3390/ijms20051143
Chicago/Turabian StyleAntholine, William E., Jeannette Vasquez-Vivar, Brendan J. Quirk, Harry T. Whelan, Pui Kei Wu, Jong-In Park, and Charles R. Myers. 2019. "Treatment of Cells and Tissues with Chromate Maximizes Mitochondrial 2Fe2S EPR Signals" International Journal of Molecular Sciences 20, no. 5: 1143. https://doi.org/10.3390/ijms20051143