Inhibitory Effects of Menadione on Helicobacter pylori Growth and Helicobacter pylori-Induced Inflammation via NF-κB Inhibition

 ,
,
Abstract
:1. Introduction
2. Results
2.1. The Inhibitory Effect of Menadione on the Growth of H. pylori
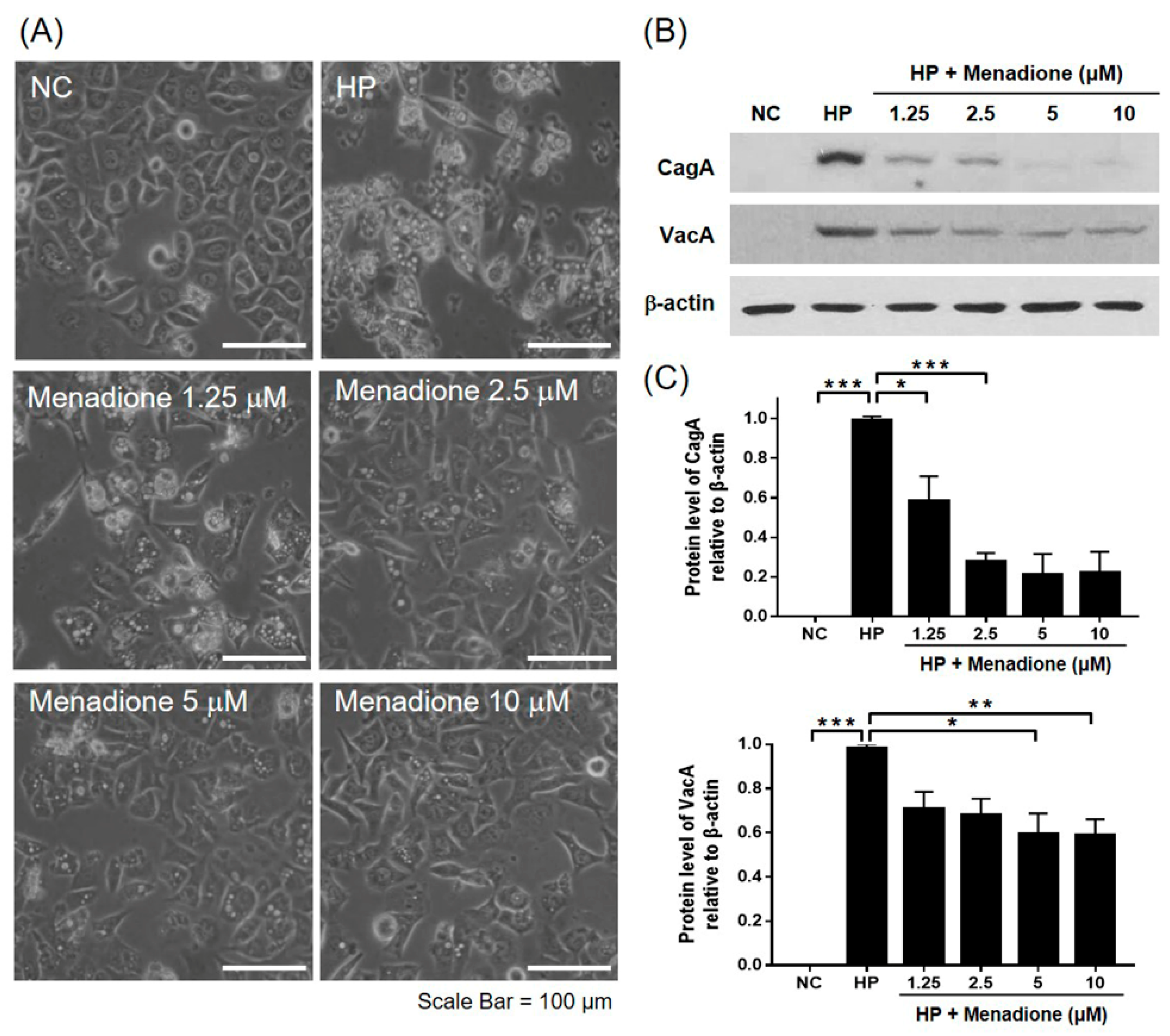
2.2. Menadione Reduced CagA and VacA Translocation to AGS Cells and Recovered Morphological Changes Caused by H. pylori Infection
2.3. Menadione Alleviated H. pylori-Induced Apoptosis and Death of AGS Cells
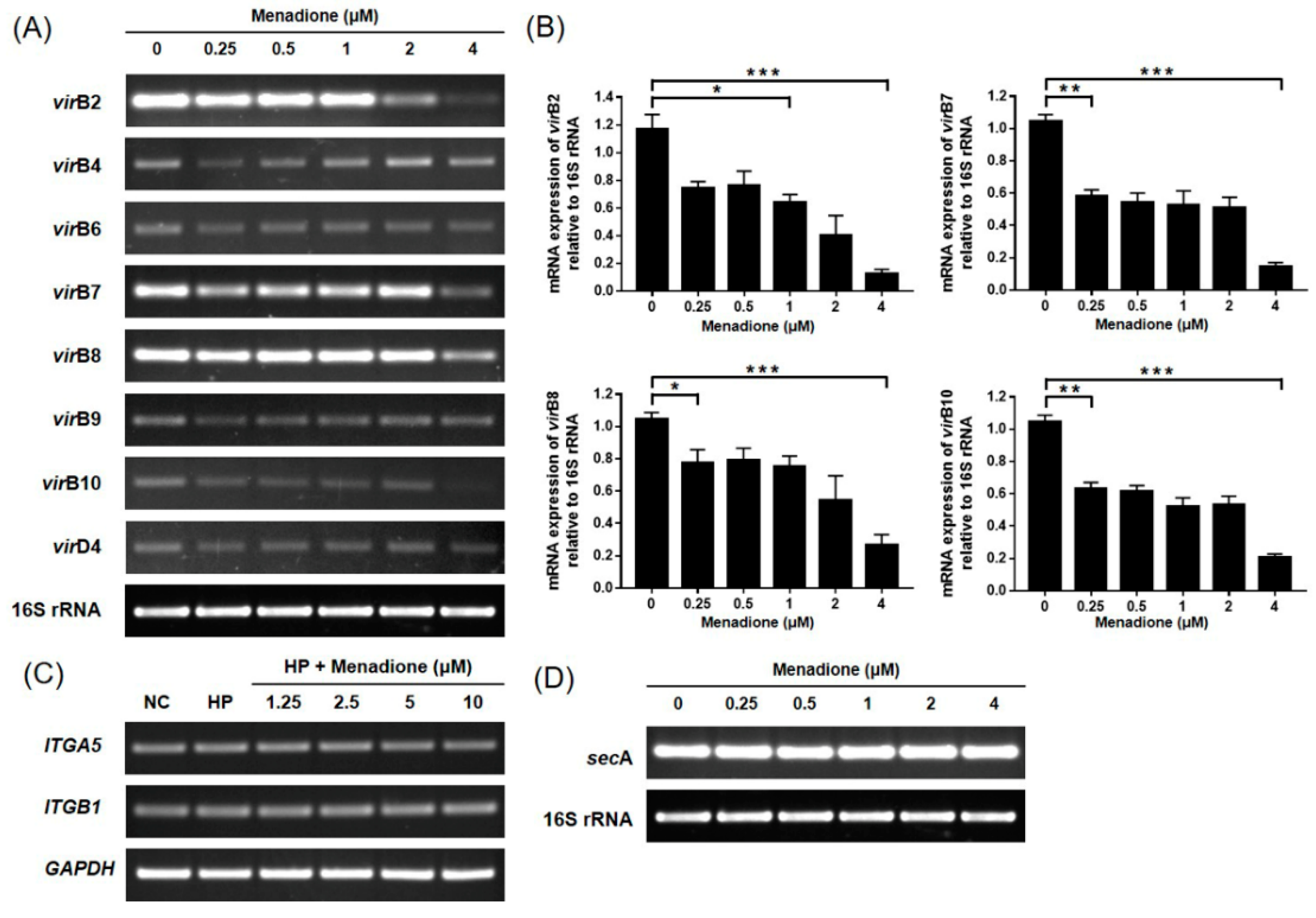
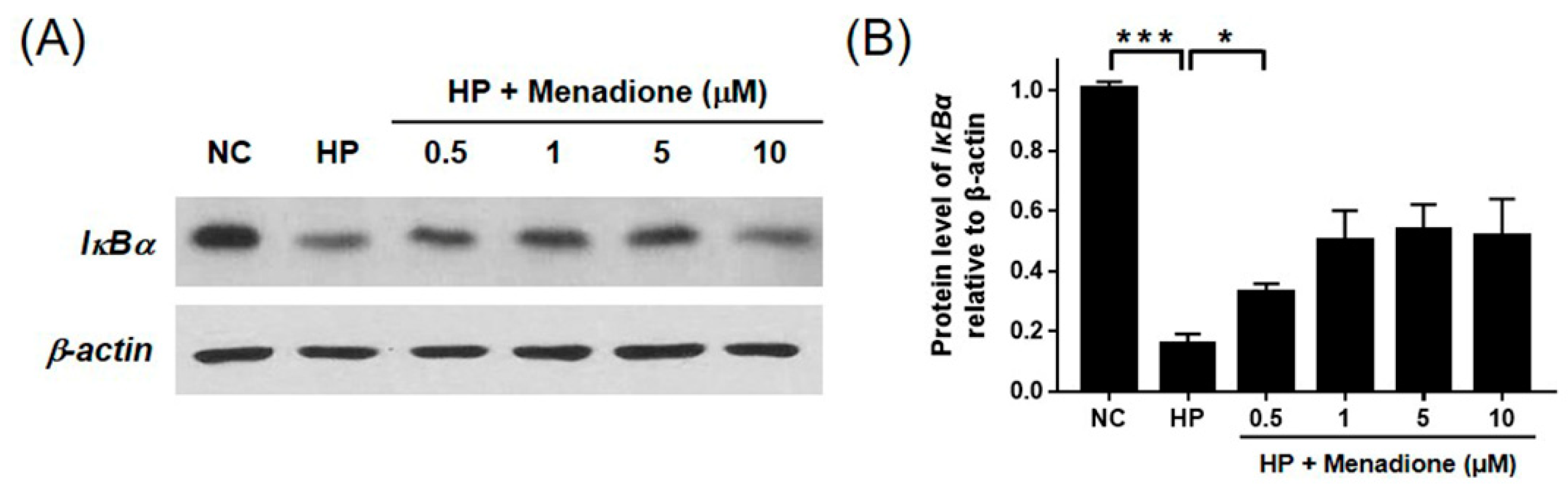
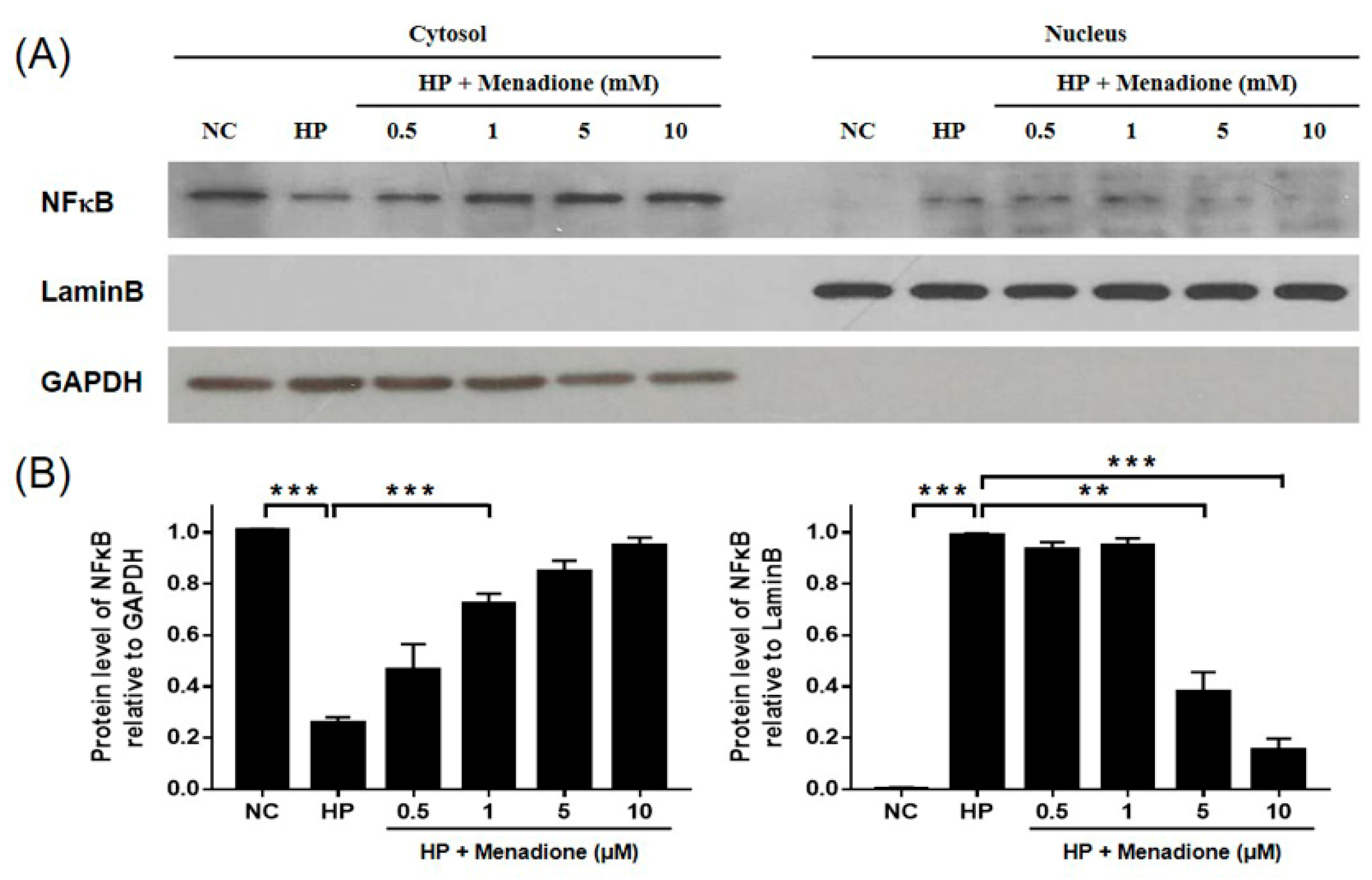
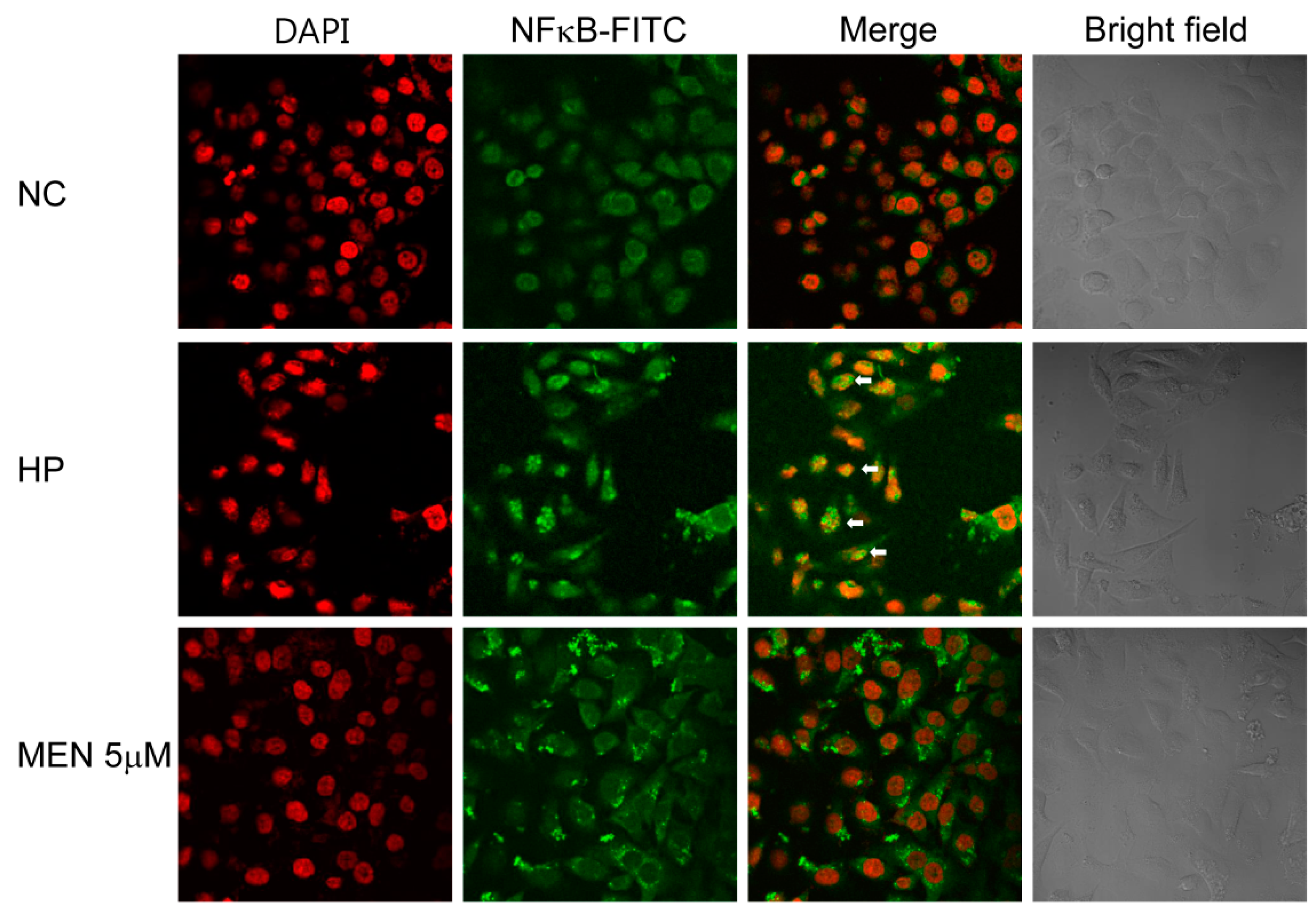
2.4. Menadione Reduces Expression of Inflammatory Cytokines Induced by H. pylori-Induced Infection via Inhibition of NF-κB Activation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Collection of H. pylori Clinical Isolates
4.3. Agar Dilution Test to Determine MIC
4.4. Mammalian Cell Culture
4.5. RT-PCR
4.6. Western Blotting
4.7. Subcellular Fractionation
4.8. Confocal Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wiedemann, T.; Loell, E.; Mueller, S.; Stoeckelhuber, M.; Stolte, M.; Haas, R.; Rieder, G. Helicobacter pylori cag-Pathogenicity island-dependent early immunological response triggers later precancerous gastric changes in Mongolian gerbils. PLoS ONE 2009, 4, e4754. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blaser, M.J. Helicobacter pylori in health and disease. Gastroenterology 2009, 136, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A. Cellular substrates of p34(cdc2) and its companion cyclin-dependent kinases. Trends Cell Biol. 1993, 3, 296–301. [Google Scholar] [CrossRef]
- De Martel, C.; Ferlay, J.; Franceschi, S.; Vignat, J.; Bray, F.; Forman, D.; Plummer, M. Global burden of cancers attributable to infections in 2008: A review and synthetic analysis. Lancet Oncol. 2012, 13, 607–615. [Google Scholar] [CrossRef]
- Odenbreit, S.; Puls, J.; Sedlmaier, B.; Gerland, E.; Fischer, W.; Haas, R. Translocation of Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science 2000, 287, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Kwok, T.; Zabler, D.; Urman, S.; Rohde, M.; Hartig, R.; Wessler, S.; Misselwitz, R.; Berger, J.; Sewald, N.; Konig, W.; et al. Helicobacter exploits integrin for type IV secretion and kinase activation. Nature 2007, 449, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Terradot, L.; Waksman, G. Architecture of the Helicobacter pylori Cag-type IV secretion system. FEBS J. 2011, 278, 1213–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, P.J. Type IV secretion: The Agrobacterium VirB/D4 and related conjugation systems. Biochim. Biophys. Acta 2004, 1694, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Christie, P.J.; Cascales, E. Structural and dynamic properties of bacterial type IV secretion systems (review). Mol. Membr. Biol. 2005, 22, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Bornschein, J.; Malfertheiner, P. Helicobacter pylori and gastric cancer. Dig. Dis. 2014, 32, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, M. Oncogenic mechanisms of the Helicobacter pylori CagA protein. Nat. Rev. Cancer 2004, 4, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Saadat, I.; Higashi, H.; Obuse, C.; Umeda, M.; Murata-Kamiya, N.; Saito, Y.; Lu, H.; Ohnishi, N.; Azuma, T.; Suzuki, A.; et al. Helicobacter pylori CagA targets PAR1/MARK kinase to disrupt epithelial cell polarity. Nature 2007, 447, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Bagnoli, F.; Halenbeck, R.; Rappuoli, R.; Fantl, W.J.; Covacci, A. C-Src/Lyn kinases activate Helicobacter pylori CagA through tyrosine phosphorylation of the EPIYA motifs. Mol. Microbiol. 2002, 43, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blanke, S.R. Helicobacter pylori VacA, a paradigm for toxin multifunctionality. Nat. Rev. Microbiol. 2005, 3, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Boquet, P.; Ricci, V. Intoxication strategy of Helicobacter pylori VacA toxin. Trends Microbiol. 2012, 20, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Leyton, D.L.; Rossiter, A.E.; Henderson, I.R. From self sufficiency to dependence: Mechanisms and factors important for autotransporter biogenesis. Nat. Rev. Microbiol. 2012, 10, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Woo, H.; Park, M.; Rhee, K.J.; Moon, C.; Lee, D.; Seo, W.D.; Kim, J.B. Cyanidin 3-O-glucoside reduces Helicobacter pylori VacA-induced cell death of gastric KATO III cells through inhibition of the SecA pathway. Int. J. Med Sci. 2014, 11, 742–747. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Schraw, W.; Ricci, V.; Boquet, P.; Cover, T.L. Acid activation of Helicobacter pylori vacuolating cytotoxin (VacA) results in toxin internalization by eukaryotic cells. Mol. Microbiol. 2000, 37, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boquet, P.; Ricci, V.; Galmiche, A.; Gauthier, N.C. Gastric cell apoptosis and H. pylori: Has the main function of VacA finally been identified? Trends Microbiol. 2003, 11, 410–413. [Google Scholar] [CrossRef]
- Cover, T.L.; Krishna, U.S.; Israel, D.A.; Peek, R.M., Jr. Induction of gastric epithelial cell apoptosis by Helicobacter pylori vacuolating cytotoxin. Cancer Res. 2003, 63, 951–957. [Google Scholar] [PubMed]
- Galmiche, A.; Rassow, J.; Doye, A.; Cagnol, S.; Chambard, J.C.; Contamin, S.; de Thillot, V.; Just, I.; Ricci, V.; Solcia, E.; et al. The N-terminal 34 kDa fragment of Helicobacter pylori vacuolating cytotoxin targets mitochondria and induces cytochrome c release. EMBO J. 2000, 19, 6361–6370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuck, D.; Kolmerer, B.; Iking-Konert, C.; Krammer, P.H.; Stremmel, W.; Rudi, J. Vacuolating cytotoxin of Helicobacter pylori induces apoptosis in the human gastric epithelial cell line AGS. Infect. Immun. 2001, 69, 5080–5087. [Google Scholar] [CrossRef] [PubMed]
- Qadri, Q.; Rasool, R.; Gulzar, G.M.; Naqash, S.; Shah, Z.A.H. pylori infection, inflammation and gastric cancer. J. Gastrointest. Cancer 2014, 45, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.; Seymour, R.; Henderson, B. Bacterial perturbation of cytokine networks. Infect. Immun. 1998, 66, 2401–2409. [Google Scholar] [PubMed]
- Zarrilli, R.; Ricci, V.; Romano, M. Molecular response of gastric epithelial cells to Helicobacter pylori-induced cell damage. Cell. Microbiol. 1999, 1, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munzenmaier, A.; Lange, C.; Glocker, E.; Covacci, A.; Moran, A.; Bereswill, S.; Baeuerle, P.A.; Kist, M.; Pahl, H.L. A secreted/shed product of Helicobacter pylori activates transcription factor nuclear factor-kappa B. J. Immunol. 1997, 159, 6140–6147. [Google Scholar] [PubMed]
- Sharma, S.A.; Tummuru, M.K.; Blaser, M.J.; Kerr, L.D. Activation of IL-8 gene expression by Helicobacter pylori is regulated by transcription factor nuclear factor-kappa B in gastric epithelial cells. J. Immunol. 1998, 160, 2401–2407. [Google Scholar] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Hatz, R.A.; Rieder, G.; Stolte, M.; Bayerdorffer, E.; Meimarakis, G.; Schildberg, F.W.; Enders, G. Pattern of adhesion molecule expression on vascular endothelium in Helicobacter pylori-associated antral gastritis. Gastroenterology 1997, 112, 1908–1919. [Google Scholar] [CrossRef] [PubMed]
- Hassan, G.S. Menadione. In Profiles of Drug Substances, Excipients, and Related Methodology; Academic Press: Cambridge, MA, USA, 2013; Volume 38, pp. 227–313. [Google Scholar]
- Al-Suhaimi, E. Molecular mechanisms of leptin and pro-apoptotic signals induced by menadione in HepG2 cells. Saudi J. Biol. Sci. 2014, 21, 582–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, D.; Ray, R.; Hazra, B. Antitubercular and antibacterial activity of quinonoid natural products against multi-drug resistant clinical isolates. Phytother. Res. PTR 2014, 28, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Park, B.S.; Lee, H.K.; Lee, S.E.; Piao, X.L.; Takeoka, G.R.; Wong, R.Y.; Ahn, Y.J.; Kim, J.H. Antibacterial activity of Tabebuia impetiginosa Martius ex DC (Taheebo) against Helicobacter pylori. J. Ethnopharmacol. 2006, 105, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Merriman, J.A.; Salgado-Pabon, W.; Mueller, E.A.; Spaulding, A.R.; Vu, B.G.; Chuang-Smith, O.N.; Kohler, P.L.; Kirby, J.R. Menaquinone analogs inhibit growth of bacterial pathogens. Antimicrob. Agents Chemother. 2013, 57, 5432–5437. [Google Scholar] [CrossRef] [PubMed]
- Sreelatha, T.; Kandhasamy, S.; Dinesh, R.; Shruthy, S.; Shweta, S.; Mukesh, D.; Karunagaran, D.; Balaji, R.; Mathivanan, N.; Perumal, P.T. Synthesis and SAR study of novel anticancer and antimicrobial naphthoquinone amide derivatives. Bioorg. Med. Chem. Lett. 2014, 24, 3647–3651. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.C.; Morais Braga, M.F.; Guedes, G.M.; Tintino, S.R.; Freitas, M.A.; Quintans, L.J., Jr.; Menezes, I.R.; Coutinho, H.D. Menadione (vitamin K) enhances the antibiotic activity of drugs by cell membrane permeabilization mechanism. Saudi J. Biol. Sci. 2017, 24, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, S.; Nishiumi, S.; Nishida, M.; Mizushina, Y.; Kobayashi, K.; Masuda, A.; Fujita, T.; Morita, Y.; Mizuno, S.; Kutsumi, H.; et al. Vitamin K3 attenuates lipopolysaccharide-induced acute lung injury through inhibition of nuclear factor-kappaB activation. Clin. Exp. Immunol. 2010, 160, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Segal, E.D.; Cha, J.; Lo, J.; Falkow, S.; Tompkins, L.S. Altered states: Involvement of phosphorylated CagA in the induction of host cellular growth changes by Helicobacter pylori. Proc. Natl. Acad. Sci. USA 1999, 96, 14559–14564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.X.; Zhou, L.Y.; Song, Z.Q.; Zhang, J.Z.; He, L.H.; Ding, Y. Primary antibiotic resistance of Helicobacter pylori strains isolated from patients with dyspeptic symptoms in Beijing: A prospective serial study. World J. Gastroenterol. 2015, 21, 2786–2792. [Google Scholar] [CrossRef] [PubMed]
- Wu, I.T.; Chuah, S.K.; Lee, C.H.; Liang, C.M.; Lu, L.S.; Kuo, Y.H.; Yen, Y.H.; Hu, M.L.; Chou, Y.P.; Yang, S.C.; et al. Five-year sequential changes in secondary antibiotic resistance of Helicobacter pylori in Taiwan. World J. Gastroenterol. 2015, 21, 10669–10674. [Google Scholar] [CrossRef] [PubMed]
- Ghotaslou, R.; Leylabadlo, H.E.; Asl, Y.M. Prevalence of antibiotic resistance in Helicobacter pylori: A recent literature review. World J. Methodol. 2015, 5, 164–174. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Moon, B.S.; Kim, H.; Lim, H.C.; Lee, Y.C.; Lee, G.; Kim, S.H.; Park, M.; Kim, J.B. Antibiotic resistance in Helicobacter pylori strains and its effect on H. pylori eradication rates in a single center in Korea. Ann. Lab. Med. 2013, 33, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.A.; Han, M.J.; Baek, N.I.; Kim, D.H. In vitro anti-Helicobacter pylori activity of panaxytriol isolated from ginseng. Arch. Pharmacal Res. 2001, 24, 297–299. [Google Scholar] [CrossRef]
- Kubo, J.; Lee, J.R.; Kubo, I. Anti-Helicobacter pylori agents from the cashew apple. J. Agric. Food Chem. 1999, 47, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Rho, T.C.; Bae, E.A.; Kim, D.H.; Oh, W.K.; Kim, B.Y.; Ahn, J.S.; Lee, H.S. Anti-Helicobacter pylori activity of quinolone alkaloids from Evodiae fructus. Biol. Pharm. Bull. 1999, 22, 1141–1143. [Google Scholar] [CrossRef] [PubMed]
- Ohsaki, A.; Takashima, J.; Chiba, N.; Kawamura, M. Microanalysis of a selective potent anti-Helicobacter pylori compound in a Brazilian medicinal plant, Myroxylon peruiferum and the activity of analogues. Bioorg. Med. Chem. Lett. 1999, 9, 1109–1112. [Google Scholar] [CrossRef]
- Matsubara, S.; Shibata, H.; Ishikawa, F.; Yokokura, T.; Takahashi, M.; Sugimura, T.; Wakabayashi, K. Suppression of Helicobacter pylori-induced gastritis by green tea extract in Mongolian gerbils. Biochem. Biophys. Res. Commun. 2003, 310, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Lamb, A.; Yang, X.D.; Tsang, Y.H.; Li, J.D.; Higashi, H.; Hatakeyama, M.; Peek, R.M.; Blanke, S.R.; Chen, L.F. Helicobacter pylori CagA activates NF-kappaB by targeting TAK1 for TRAF6-mediated Lys 63 ubiquitination. EMBO Rep. 2009, 10, 1242–1249. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Lee, S.J.; Kagnoff, M.F. Nod1 is an essential signal transducer in intestinal epithelial cells infected with bacteria that avoid recognition by toll-like receptors. Infect. Immun. 2004, 72, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Brandt, S.; Kwok, T.; Hartig, R.; Konig, W.; Backert, S. NF-kappaB activation and potentiation of proinflammatory responses by the Helicobacter pylori CagA protein. Proc. Natl. Acad. Sci. USA 2005, 102, 9300–9305. [Google Scholar] [CrossRef] [PubMed]
- Papadakos, K.S.; Sougleri, I.S.; Mentis, A.F.; Hatziloukas, E.; Sgouras, D.N. Presence of terminal EPIYA phosphorylation motifs in Helicobacter pylori CagA contributes to IL-8 secretion, irrespective of the number of repeats. PLoS ONE 2013, 8, e56291. [Google Scholar] [CrossRef] [PubMed]
- Viala, J.; Chaput, C.; Boneca, I.G.; Cardona, A.; Girardin, S.E.; Moran, A.P.; Athman, R.; Memet, S.; Huerre, M.R.; Coyle, A.J.; et al. Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat. Immunol. 2004, 5, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Aoganghua, A.; Nishiumi, S.; Kobayashi, K.; Nishida, M.; Kuramochi, K.; Tsubaki, K.; Hirai, M.; Tanaka, S.; Azuma, T.; Yoshida, H.; et al. Inhibitory effects of vitamin K(3) derivatives on DNA polymerase and inflammatory activity. Int. J. Mol. Med. 2011, 28, 937–945. [Google Scholar] [PubMed]
- Checker, R.; Sharma, D.; Sandur, S.K.; Khan, N.M.; Patwardhan, R.S.; Kohli, V.; Sainis, K.B. Vitamin K3 suppressed inflammatory and immune responses in a redox-dependent manner. Free Radic. Res. 2011, 45, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Pinho, B.R.; Sousa, C.; Valentao, P.; Andrade, P.B. Is nitric oxide decrease observed with naphthoquinones in LPS stimulated RAW 264.7 macrophages a beneficial property? PLoS ONE 2011, 6, e24098. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Goto, S.; Wada, A.; Yahiro, K.; Niidome, T.; Hatakeyama, T.; Aoyagi, H.; Hirayama, T.; Kondo, T. Vacuolating cytotoxin purified from Helicobacter pylori causes mitochondrial damage in human gastric cells. Microb. Pathog. 1999, 26, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Willhite, D.C.; Blanke, S.R. Helicobacter pylori vacuolating cytotoxin enters cells, localizes to the mitochondria, and induces mitochondrial membrane permeability changes correlated to toxin channel activity. Cell. Microbiol. 2004, 6, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Supajatura, V.; Ushio, H.; Wada, A.; Yahiro, K.; Okumura, K.; Ogawa, H.; Hirayama, T.; Ra, C. Cutting edge: VacA, a vacuolating cytotoxin of Helicobacter pylori, directly activates mast cells for migration and production of proinflammatory cytokines. J. Immunol. 2002, 168, 2603–2607. [Google Scholar] [CrossRef] [PubMed]
- Boncristiano, M.; Paccani, S.R.; Barone, S.; Ulivieri, C.; Patrussi, L.; Ilver, D.; Amedei, A.; D’Elios, M.M.; Telford, J.L.; Baldari, C.T. The Helicobacter pylori vacuolating toxin inhibits T cell activation by two independent mechanisms. J. Exp. Med. 2003, 198, 1887–1897. [Google Scholar] [CrossRef] [PubMed]
- Tharmalingam, N.; Kim, S.H.; Park, M.; Woo, H.J.; Kim, H.W.; Yang, J.Y.; Rhee, K.J.; Kim, J.B. Inhibitory effect of piperine on Helicobacter pylori growth and adhesion to gastric adenocarcinoma cells. Infect. Agent Cancer 2014, 9, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, B.; Moon, B.S.; Lim, H.C.; Lee, Y.C.; Kim, H.; Lee, K.; Kim, S.H.; Park, M.; Kim, J.B. Analysis of Gene Mutations Associated with Antibiotic Resistance in Helicobacter pylori Strains Isolated from Korean Patients. Korean J. Helicobacter Upper Gastrointest. Res. 2014, 14, 95–102. [Google Scholar] [CrossRef]
- Boonjakuakul, J.K.; Canfield, D.R.; Solnick, J.V. Comparison of Helicobacter pylori virulence gene expression in vitro and in the Rhesus macaque. Infect. Immun. 2005, 73, 4895–4904. [Google Scholar] [CrossRef] [PubMed]
- Yeon, M.J.; Lee, M.H.; Kim, D.H.; Yang, J.Y.; Woo, H.J.; Kwon, H.J.; Moon, C.; Kim, S.H.; Kim, J.B. Anti-inflammatory effects of Kaempferol on Helicobacter pylori-induced inflammation. Biosci. Biotechnol. Biochem. 2019, 83, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, M.; Woo, H.; Tharmalingam, N.; Lee, G.; Rhee, K.J.; Eom, Y.B.; Han, S.I.; Seo, W.D.; Kim, J.B. Inhibitory effects of anthocyanins on secretion of Helicobacter pylori CagA and VacA toxins. Int. J. Med. Sci. 2012, 9, 838–842. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Cho, Y.; Jung, B.C.; Kim, S.H.; Kang, Y.W.; Pan, C.H.; Rhee, K.J.; Kim, Y.S. Parkin induces G2/M cell cycle arrest in TNF-alpha-treated HeLa cells. Biochem. Biophys. Res. Commun. 2015, 464, 63–69. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Menadione Concentration (M) | Number of Strains (n = 38) |
|---|---|
| 1 | 1 (2.6%) |
| 2 | 4 (10.5%) |
| 4 | 8 (21.1%) |
| 8 | 22 (57.9%) |
| 16 | 2 (5.3%) |
| 32 | 1 (2.6%) |
| Primers | Sequences (5′–3′) | Product Length (bp) | Annealing Temperature (°C) | Cycles | Reference * | |
|---|---|---|---|---|---|---|
| Forward | Reverse | |||||
| 16s rRNA | TGCAGCTAACGCATTAAGCATC | CATTCTGGCTTCAGTGTAACG | 642 | 52 | 16 | [63] |
| CagA | TGGCAGTGGGTTAGTCATA | CCTGTGAGTTGGTCTTCTTGT | 278 | 45 | 35 | [64] |
| VacA | AAACGACAAGAAAGAGATCAGT | CCAGCAAAAGGCCCATCAA | 291 | 57 | 22 | |
| VirB2 | CAGTCGCCTGACCTCTTTTGA | CGGTCACCAGTCCTGCAAC | 156 | 62 | 25 | [65] |
| VirB4 | GTTATAGGGGCAACCGGAAG | TTGAACGCGTCATTCAAAGC | 449 | 62 | 37 | |
| VirB5 | TACAAGCGTCTGTGAAGCAG | GACCAACCAACAAGTGCTCA | 436 | 62 | 29 | |
| VirB6 | CCTCAACACCGCCTTTGGTA | TAGCCGCTAGCAATCTGGTG | 225 | 62 | 25 | |
| VirB7 | GATTACGCTCATAGGCGATGC | TGGCTGACTTCCTTGCAACA | 202 | 62 | 25 | |
| VirB8 | GTTGATCCTTGCGATCCCTCA | CGCCGCTGTAACGAGTATTG | 218 | 62 | 25 | |
| VirB9 | GCATGTCCTCTAGTCGTTCCA | TATCGTAGATGCGCCTGACC | 269 | 62 | 25 | |
| VirB10 | TCCACTTCATCAGCTTGTCG | CTAACGACAGAGCGGCTATC | 361 | 62 | 31 | |
| VirD4 | CCGCAAGTTTCCATAGTGTC | GCGAGTTGGGAAACTGAAGA | 263 | 62 | 25 | |
| SecA | AAAAATTTGACGCTGTGATCC | CCCCCAAGCTCCTTAATTTC | 274 | 47 | 27 | [66] |
| ITGA5 | GTGACTACTTTGCCGTGAAC | AGTCGCTTACTGGGAATAGC | 276 | 60 | 25 | |
| ITGB1 | GAGAATCCAGAGTGTCCCAC | ACAGTTGTTACGGCACTCTT | 215 | 60 | 21 | |
| GAPDH | CGGGAAGCTTGTCATCAATGG | GGCAGTGATGGCATGGACTG | 349 | 55 | 20 | [67] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.H.; Yang, J.Y.; Cho, Y.; Woo, H.J.; Kwon, H.J.; Kim, D.H.; Park, M.; Moon, C.; Yeon, M.J.; Kim, H.W.; et al. Inhibitory Effects of Menadione on Helicobacter pylori Growth and Helicobacter pylori-Induced Inflammation via NF-κB Inhibition. Int. J. Mol. Sci. 2019, 20, 1169. https://doi.org/10.3390/ijms20051169
Lee MH, Yang JY, Cho Y, Woo HJ, Kwon HJ, Kim DH, Park M, Moon C, Yeon MJ, Kim HW, et al. Inhibitory Effects of Menadione on Helicobacter pylori Growth and Helicobacter pylori-Induced Inflammation via NF-κB Inhibition. International Journal of Molecular Sciences. 2019; 20(5):1169. https://doi.org/10.3390/ijms20051169
Chicago/Turabian StyleLee, Min Ho, Ji Yeong Yang, Yoonjung Cho, Hyun Jun Woo, Hye Jin Kwon, Do Hyun Kim, Min Park, Cheol Moon, Min Ji Yeon, Hyun Woo Kim, and et al. 2019. "Inhibitory Effects of Menadione on Helicobacter pylori Growth and Helicobacter pylori-Induced Inflammation via NF-κB Inhibition" International Journal of Molecular Sciences 20, no. 5: 1169. https://doi.org/10.3390/ijms20051169