Identification of Candidate Ergosterol-Responsive Proteins Associated with the Plasma Membrane of Arabidopsis thaliana


Abstract
:1. Introduction
2. Results
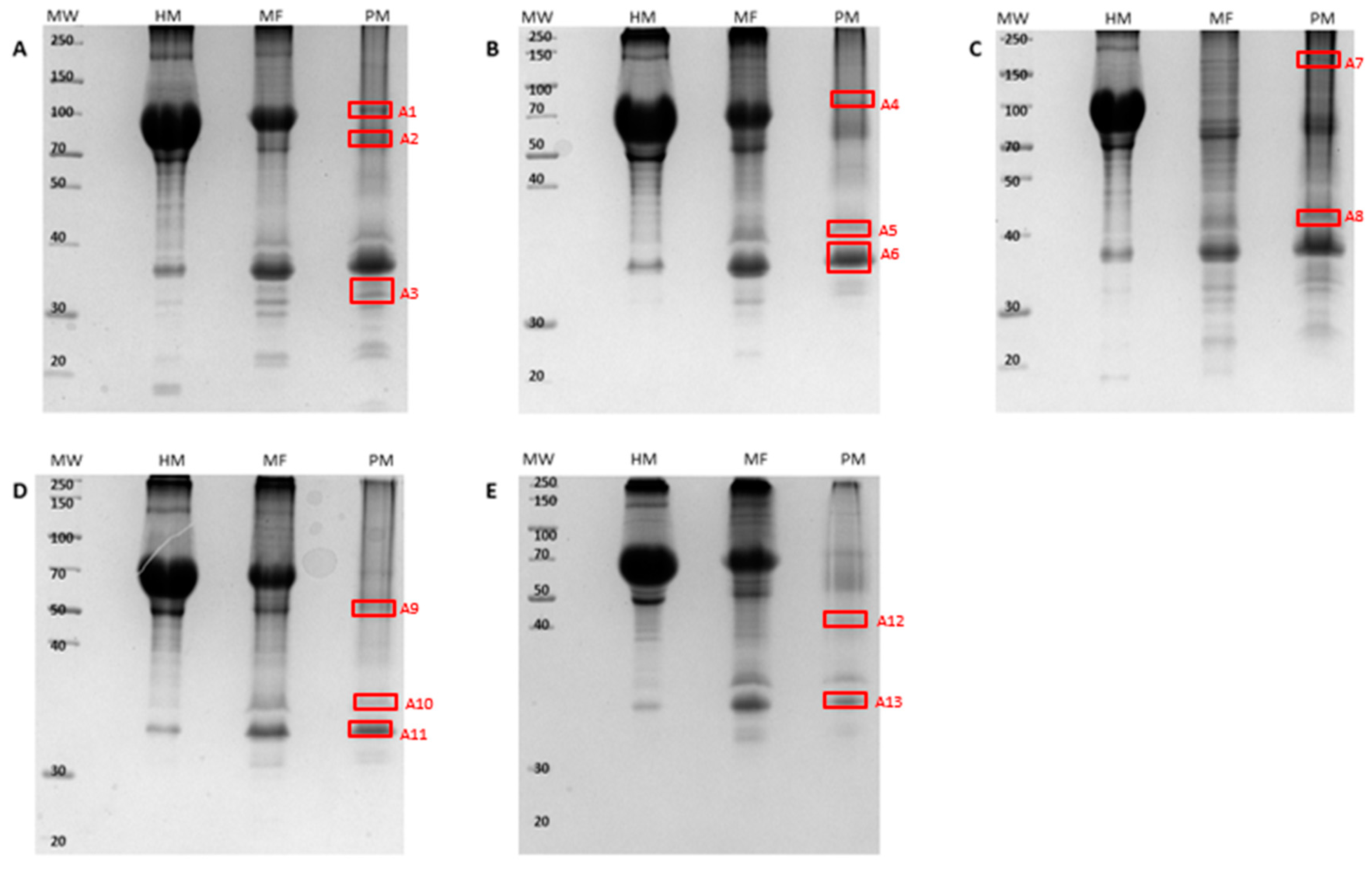
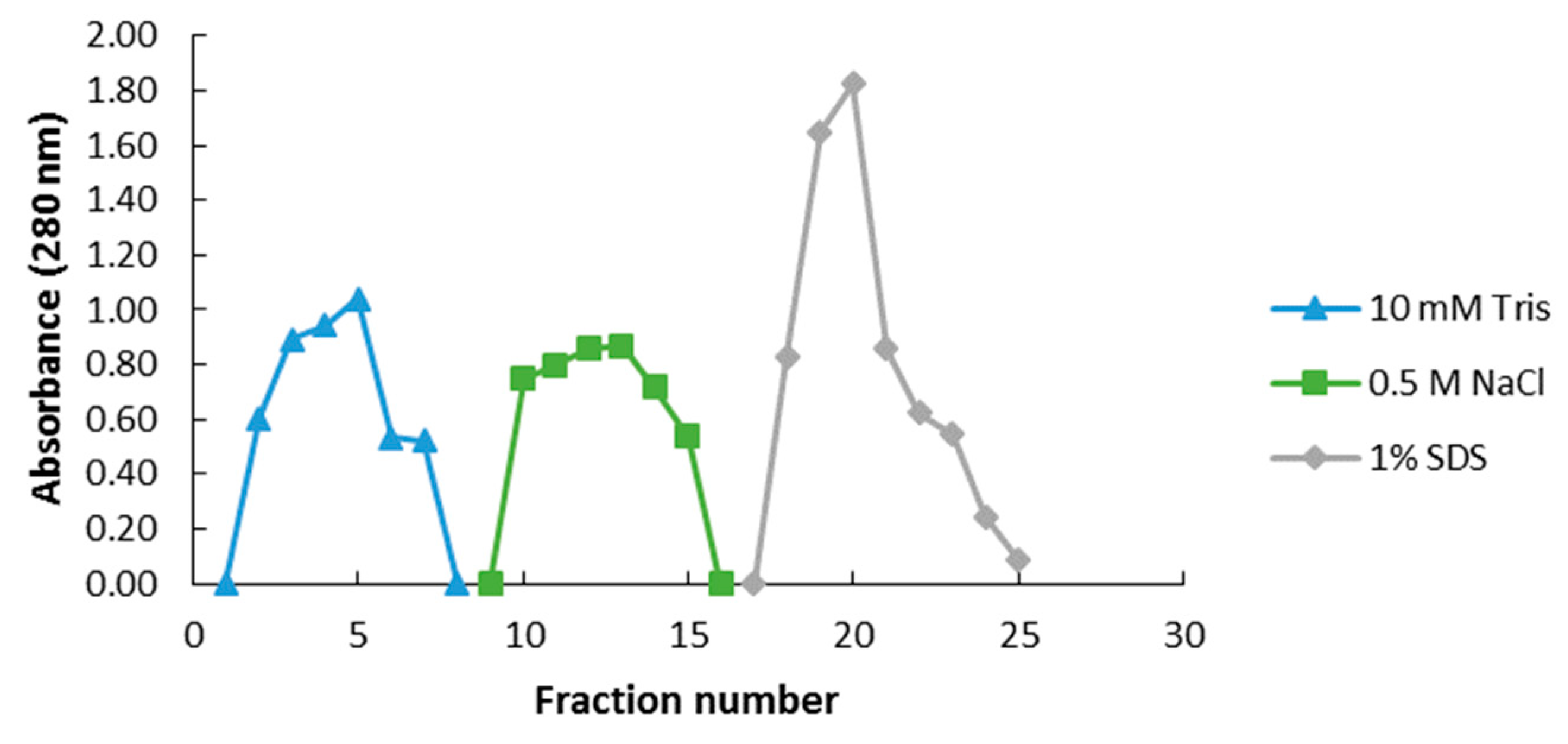
2.1. Plasma Membrane (PM)-Associated Fraction Isolation and Verification
2.2. PM-Associated Ergosterol-Responsive Candidate Protein Identification
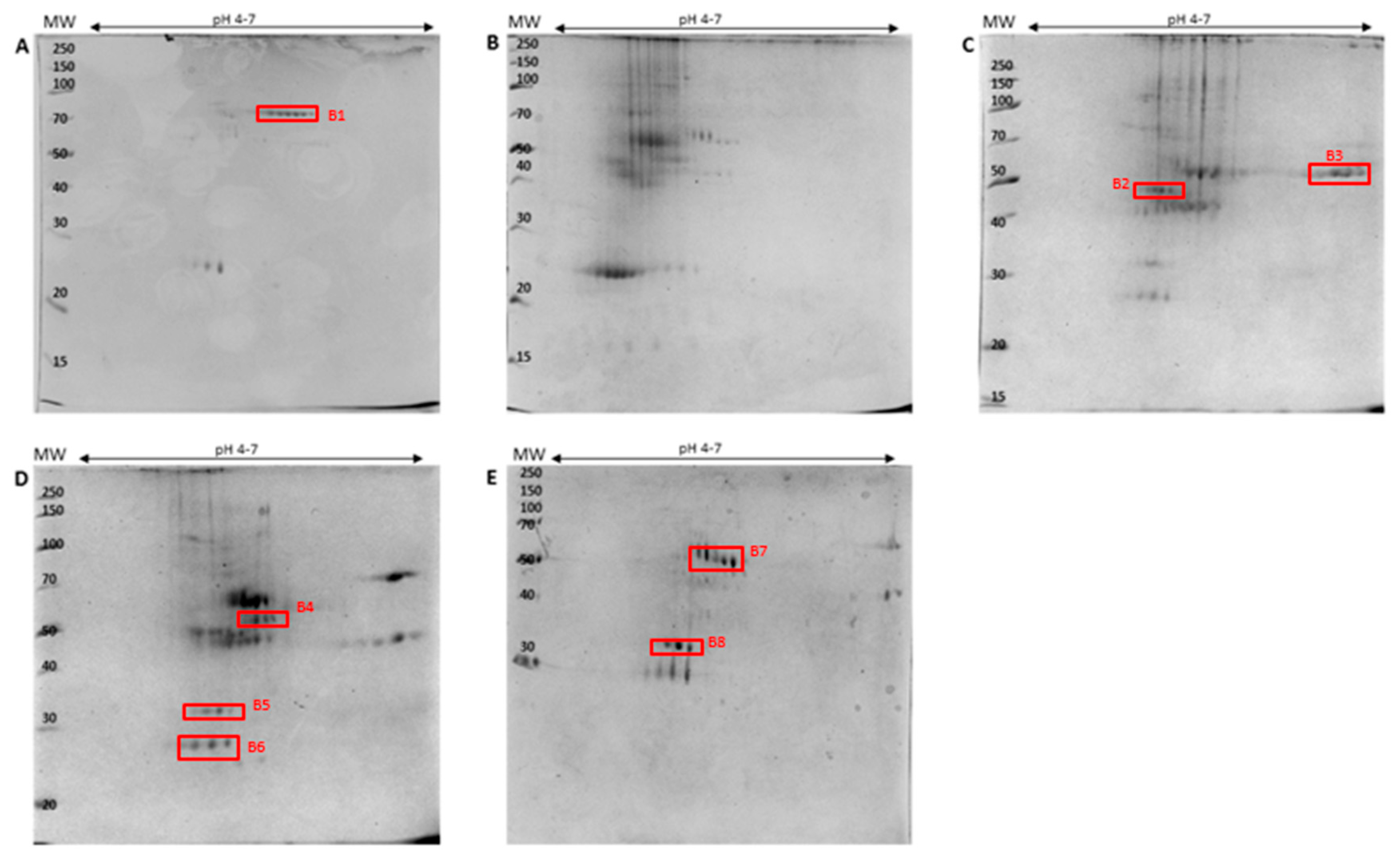
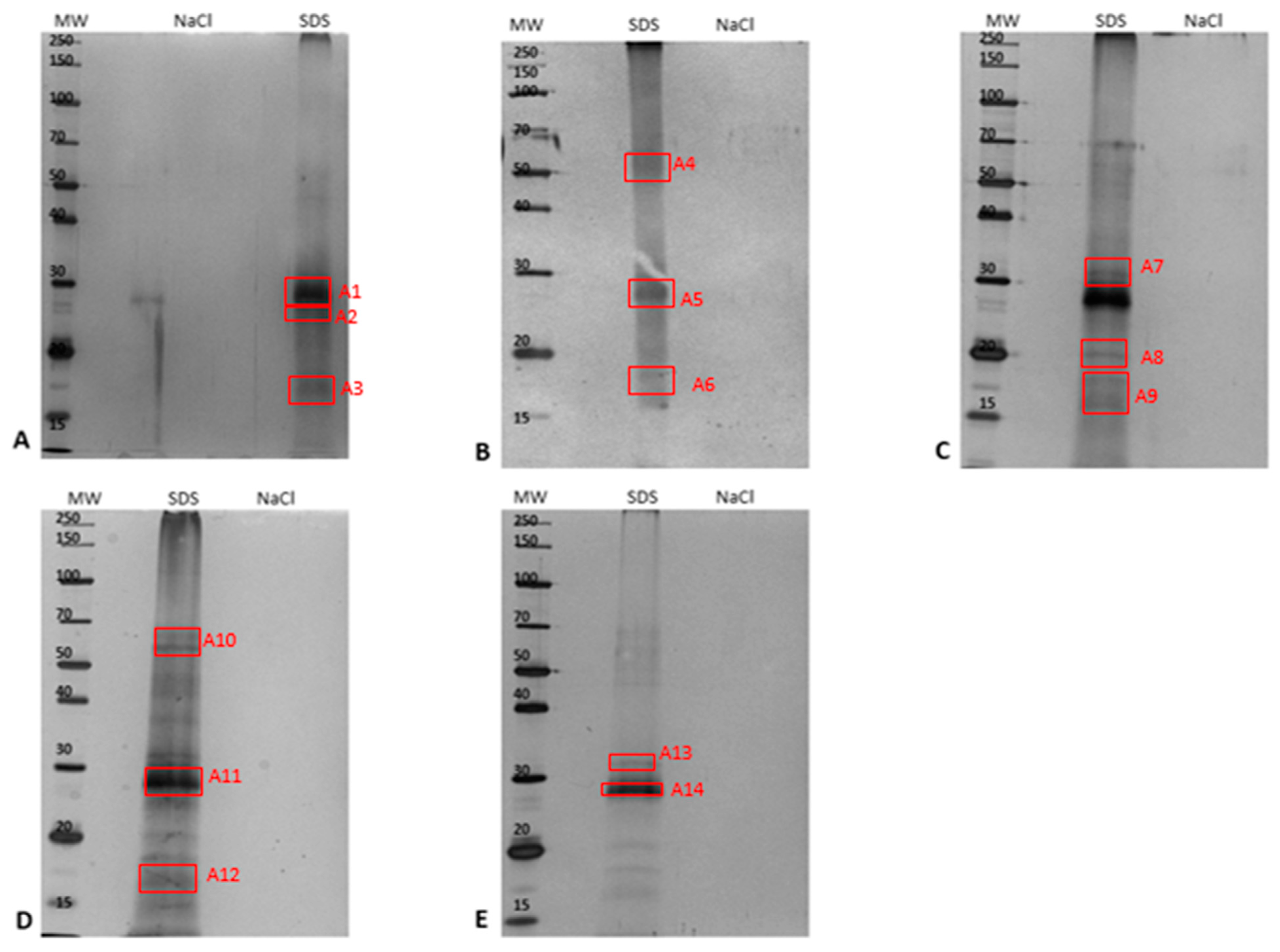
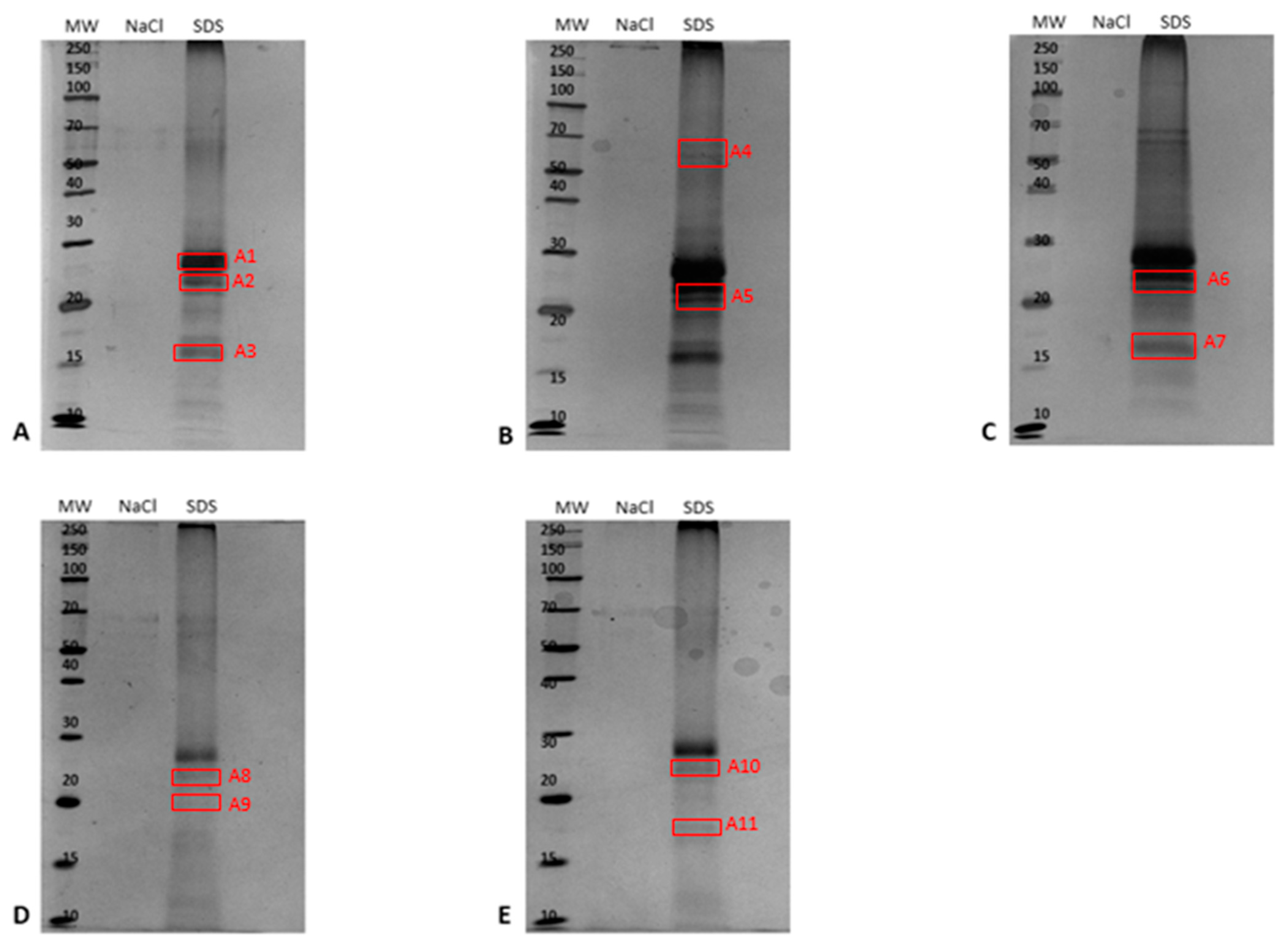
2.3. Identification of PM-Associated Ergosterol-Interacting Candidate Proteins
2.3.1. Epoxide Magnetic Microspheres-Based Ergosterol Immobilization
2.3.2. EAH Sepharose 4B Immobilized with Ergosterol-Hemisuccinate
3. Discussion
3.1. Functional Classification of Identified Ergosterol-Responsive – and Interacting Candidate PM-Associated Proteins from A. thaliana Leaf Tissue
3.1.1. Membrane Trafficking and Transporters
3.1.2. Signaling
3.1.3. Defense responses
4. Materials and Methods
4.1. Plant Growth and Elicitor Treatment
4.2. Small-Scale Isolation of the Plasma Membrane(PM)-Associated Fraction
4.3. Identification of PM-Associated Ergosterol-Responsive Candidate Proteins
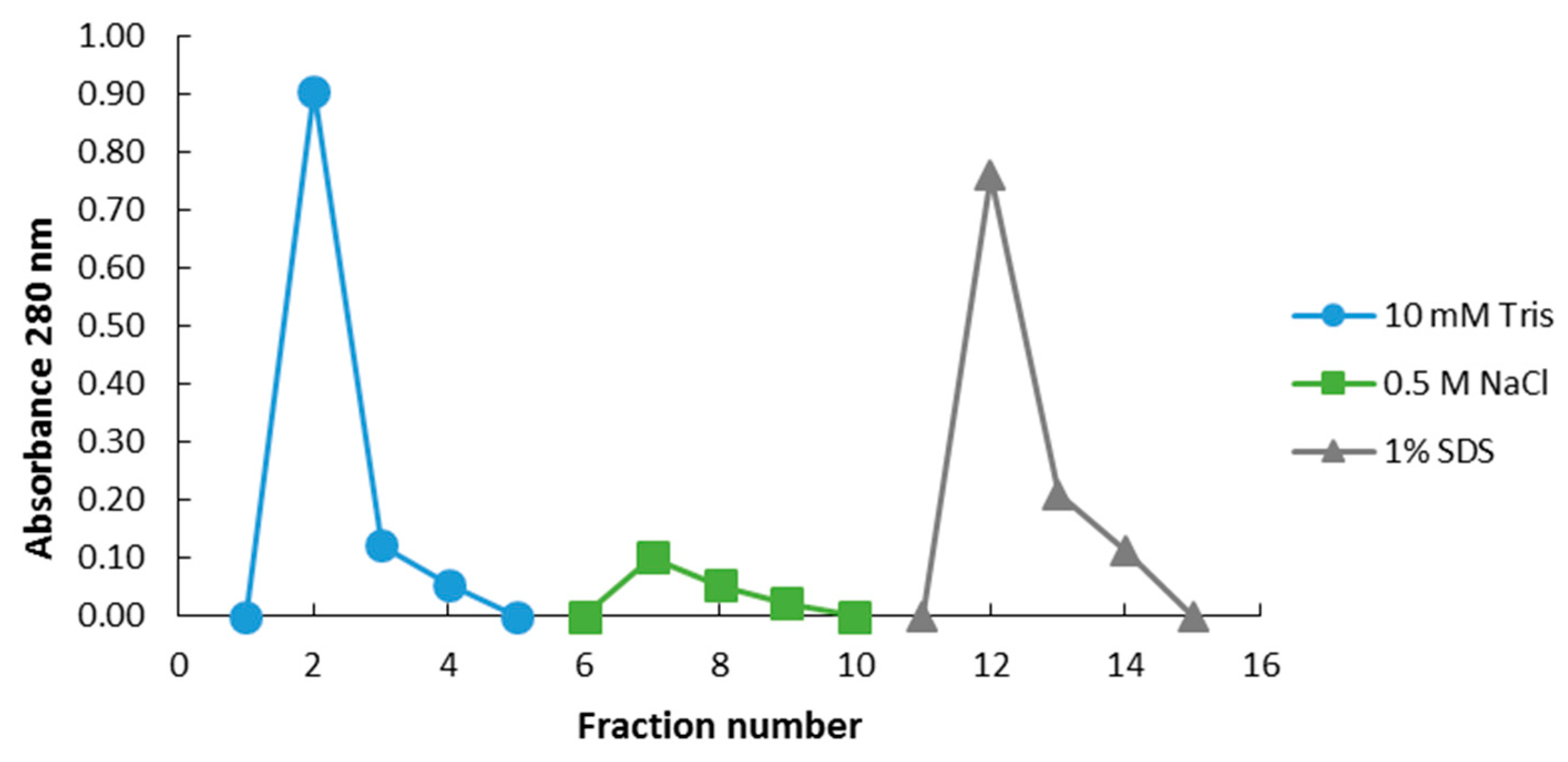
4.4. Affinity Chromatography
4.4.1. Magnetic Epoxide Microspheres
4.4.2. EAH Seharose 4B
4.5. Protein Precipitation and SDS-PAGE
4.6. In-Gel Trypsin Digestion
4.7. LC-MS/MS Analysis
4.8. Data analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gómez-Gómez, L.; Boller, T. Flagellin perception: A paradigm for innate immunity. Trends Plant Sci. 2002, 7, 251–256. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranf, S.; Gisch, N.; Schäffer, M.; Illig, T.; Westphal, L.; Knirel, Y.A.; Sánchez-Carballo, P.M.; Zähringer, U.; Hückelhoven, R.; Lee, J.; et al. A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat. Immunol. 2015, 16, 426–433. [Google Scholar] [CrossRef]
- Erbs, G.; Molinaro, A.; Dow, J.M.; Newman, M.-A. Lipopolysaccharides and plant innate immunity. Subcell. Biochem. 2010, 53, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Henry, G.; Thonart, P.; Ongena, M. PAMPs, MAMPs, DAMPs and others: An update on the diversity of plant immunity elicitors. Biotechnol. Agron. Soc. Environ. 2012, 16, 257–268. [Google Scholar]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Boyd, L.A.; Ridout, C.; O’Sullivan, D.M.; Leach, J.E.; Leung, H. Plant-pathogen interactions: Disease resistance in modern agriculture. Trends Genet. 2013, 29, 233–240. [Google Scholar] [CrossRef]
- De Wit, P.J.G.M. How plants recognize pathogens and defend themselves. Cell. Mol. Life Sci. 2007, 64, 2726–2732. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Birch, P.R.J. The zig-zag model of plant-microbe interactions: Is it time to move on? Mol. Plant Pathol. 2014, 15, 865–870. [Google Scholar] [CrossRef]
- Klemptner, R.L.; Sherwood, J.S.; Tugizimana, F.; Dubery, I.A.; Piater, L.A. Ergosterol, an orphan fungal microbe-associated molecular pattern (MAMP). Mol. Plant Pathol. 2014, 15, 747–761. [Google Scholar] [CrossRef] [PubMed]
- Granado, J.; Felix, G.; Boller, T. Perception of fungal sterols in plants. Plant Physiol. 1995, 107, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Mannock, D.A.; Lewis, R.N.A.H.; McElhaney, R.N. A calorimetric and spectroscopic comparison of the effects of ergosterol and cholesterol on the thermotropic phase behavior and organization of dipalmitoylphosphatidylcholine bilayer membranes. Biochim. Biophys. Acta 2010, 1798, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanabria, N.M.; Huang, J.-C.; Dubery, I.A. Self/nonself perception in plants in innate immunity and defense. Self/Nonself 2010, 1, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Kasparovsky, T.; Milat, M.-L.; Humbert, C.; Blein, J.-P.; Havel, L.; Mikes, V. Elicitation of tobacco cells with ergosterol activates a signal pathway including mobilization of internal calcium. Plant Physiol. Biochem. 2003, 41, 495–501. [Google Scholar] [CrossRef]
- Lochman, J.; Mikes, V. Ergosterol treatment leads to the expression of a specific set of defence-related genes in tobacco. Plant Mol. Biol. 2006, 62, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Rossard, S.; Luini, E.; Pérault, J.-M.; Bonmort, J.; Roblin, G. Early changes in membrane permeability, production of oxidative burst and modification of PAL activity induced by ergosterol in cotyledons of Mimosa pudica. J. Exp. Bot. 2006, 57, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Rossard, S.; Roblin, G.; Atanassova, R. Ergosterol triggers characteristic elicitation steps in Beta vulgaris leaf tissues. J. Exp. Bot. 2010, 61, 1807–1816. [Google Scholar] [CrossRef]
- Gaulin, E.; Bottin, A.; Dumas, B. Sterol biosynthesis in oomycete pathogens. Plant Signal. Behav. 2010, 5, 258–260. [Google Scholar] [CrossRef] [Green Version]
- Keinath, N.F.; Kierszniowska, S.; Lorek, J.; Bourdais, G.; Kessler, S.A.; Shimosato-Asano, H.; Grossniklaus, U.; Schulze, W.X.; Robatzek, S.; Panstruga, R. PAMP (pathogen-associated molecular pattern)-induced changes in plasma membrane compartmentalization reveal novel components of plant immunity. J. Biol. Chem. 2010, 285, 39140–39149. [Google Scholar] [CrossRef]
- Vatsa, P.; Chiltz, A.; Luini, E.; Vandelle, E.; Pugin, A.; Roblin, G. Cytosolic calcium rises and related events in ergosterol-treated Nicotiana cells. Plant Physiol. Biochem. 2011, 49, 764–773. [Google Scholar] [CrossRef]
- Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Ergosterol-induced sesquiterpenoid synthesis in tobacco cells. Molecules 2012, 17, 1698–1715. [Google Scholar] [CrossRef]
- Dohnal, V.; Jezkova, A.; Pavlikova, L.; Musilek, K.; Jun, D.; Kuca, K. Fluctuation in the ergosterol and deoxynivalenol content in barley and malt during malting process. Anal. Bioanal. Chem. 2010, 397, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Amborabé, B.-E.; Rossard, S.; Pérault, J.-M.; Roblin, G. Specific perception of ergosterol by plant cells. C. R. Biol. 2003, 326, 363–370. [Google Scholar] [CrossRef]
- Xu, X.; Bittman, R.; Duportail, G.; Heissler, D.; Vilcheze, C.; London, E. Effect of the structure of natural sterols and sphingolipids on the formation of ordered sphingolipid/sterol domains (rafts): Comparison of cholesterol to plant, fungal, and disease-associated sterols and comparison of sphingomyelin, cerebrosides and ceramide. J. Biol. Chem. 2001, 276, 33540–33546. [Google Scholar] [CrossRef] [PubMed]
- Giannini, J.L.; Ruiz-Cristin, J.; Briskin, D.P. A small scale procedure for the isolation of transport competent vesicles from plant tissues. Anal. Biochem. 1988, 174, 561–567. [Google Scholar] [CrossRef]
- Tsolis, K.C.; Economou, A. Quantitative proteomics of the E. coli membranome. Methods Enzymol. 2017, 586, 15–36. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Polikarpov, I.; Craievich, A.F. Average protein density is a molecular-weight-dependent function. Protein Sci. 2004, 13, 2825–2828. [Google Scholar] [CrossRef]
- Bern, M.; Kil, Y.J. Comments on “unbiased statistical analysis for multi-stage proteomic search strategies. J. Proteome Res. 2011, 10, 2123–2127. [Google Scholar] [CrossRef]
- Santoni, V.; Doumas, P.; Rouquié, D.; Mansion, M.; Rabilloud, T.; Rossignol, M. Large scale characterization of plant plasma membrane proteins. Biochimie 1999, 81, 655–661. [Google Scholar] [CrossRef]
- Alexandersson, E.; Saalbach, G.; Christer Larsson, C.; Kjellbom, P. Arabidopsis plasma membrane proteomics identifies components of transport, signal transduction and membrane trafficking. Plant Cell Physiol. 2004, 45, 1543–1556. [Google Scholar] [CrossRef] [PubMed]
- Ephritikhine, G.; Ferro, M.; Rolland, N. Plant membrane proteomics. Plant Physiol. Biochem. 2004, 42, 943–962. [Google Scholar] [CrossRef]
- He, P.; Shan, L.; Lin, N.-C.; Martin, G.B.; Kemmerling, B.; Nürnberger, T.; Sheen, J. Specific bacterial suppressors of MAMP signaling upstream of MAPKKK in Arabidopsis innate immunity. Cell 2006, 125, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fukumoto, T.; Matsumoto, T.; Gena, P.; Frascaria, D.; Kaneko, T.; Katsuhara, M.; Zhong, S.; Sun, X.; Zhu, Y.; et al. Aquaporin OsPIP1; promotes rice salt resistance and seed germination. Plant Physiol. Biochem. 2013, 63, 151–158. [Google Scholar] [CrossRef]
- Kang, J.; Park, J.; Choi, H.; Burla, B.; Kretzschmar, T.; Lee, Y.; Martinoia, E. Plant ABC transporters. Arabidopsis Book 2011, 9, e0153. [Google Scholar] [CrossRef]
- Ji, H.; Peng, Y.; Meckes, N.; Allen, S.; Stewart, C.N., Jr.; Traw, M.B. ATP-dependent binding cassette transporter G family member 16 increases plant tolerance to abscisic acid and assists in basal resistance against Pseudomonas syringae DC3000. Plant Physiol. 2014, 166, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, H.; Feng, J.; Zeng, L.; Wang, Z.; Chen, S. Grape berry plasma membrane proteome analysis and differential expression during ripening. J. Exp. Bot. 2008, 59, 2979–2990. [Google Scholar] [CrossRef] [PubMed]
- Nouri, M.-Z.; Komatsu, S. Comparative analysis of soybean plasma membrane proteins under osmotic stress using gel-based and LC MS/MS-based proteomics approaches. Proteomics 2010, 10, 1930–1945. [Google Scholar] [CrossRef]
- Vilakazi, C.S.; Dubery, I.A.; Piater, L.A. Identification of lipopolysaccharide-interacting plasma membrane-type proteins in Arabidopsis thaliana. Plant Physiol. Biochem. 2017, 111, 155–165. [Google Scholar] [CrossRef]
- Wang, C.; Yan, X.; Chen, Q.; Jiang, N.; Fu, W.; Ma, B.; Liu, J.; Li, C.; Bednarek, S.Y.; Pan, J. Clathrin light chains regulate clathrin-mediated trafficking, auxin signaling, and development in Arabidopsis. Plant Cell 2013, 25, 499–516. [Google Scholar] [CrossRef]
- Mgcina, L.S.; Dubery, I.A.; Piater, L.A. Comparative conventional- and quantum dot-labeling strategies for LPS binding sites detection in Arabidopsis thaliana mesophyll protoplasts. Front. Plant Sci. 2015, 6, 335. [Google Scholar] [CrossRef] [PubMed]
- Pandey, D.; Rajendran, S.R.C.K.; Gaur, M.; Sajeesh, P.K.; Kumar, A. Plant defense signaling and response against necrotrophic fungal pathogens. J. Plant Growth Regul. 2016, 35, 1159–1174. [Google Scholar] [CrossRef]
- Borner, G.H.H.; Liley, K.S.; Stevens, T.J.; Dupree, P. Identification of glycosylphosphatidylinositol-anchored proteins in Arabidopsis. A proteomic and genomic analysis. Plant Physiol. 2003, 132, 568–577. [Google Scholar] [CrossRef]
- Peskan, T.; Westermann, M.; Oelmüller, R. Identification of low-density Triton X-100-insoluble plasma membrane microdomains in higher plants. Eur. J. Biochem. 2000, 267, 6989–6995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borner, G.H.H.; Sherrier, D.J.; Stevens, T.J.; Arkin, I.T.; Dupree, P. Prediction of glycosylphosphatidylinositol-anchored proteins in Arabidopsis. A genomic analysis. Plant Physiol. 2002, 129, 489–499. [Google Scholar] [CrossRef]
- Jarsch, I.K.; Ott, T. Perspectives on remorin proteins, membrane rafts, and their role during plant-microbe interactions. Mol. Plant Microbe Interact. 2011, 24, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, E. Roles of lipid rafts in membrane transport. Curr. Opin. Cell Biol. 2001, 13, 470–477. [Google Scholar] [CrossRef]
- Sharma, M.; Pandey, G.K. Expansion and function of repeat domain proteins during stress and development in plants. Front. Plant Sci. 2015, 6, 1218. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, J.; Zipfel, C. Plant pattern recognition receptor complexes at the plasma membrane. Curr. Opin. Cell Biol. 2012, 15, 349–357. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.-M. Receptor kinases in plant-pathogen interactions: More than pattern recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [PubMed]
- Sanabria, N.M.; van Heerden, H.; Dubery, I.A. Molecular characterization and regulation of a Nicotiana tabacum S-domain receptor-like kinase gene induced during an early rapid response to lipopolysacharrides. Gene 2012, 501, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Baloyi, N.M.; Dubery, I.A.; Piater, L.A. Proteomic analysis of Arabidopsis plasma membranes reveals lipopolysaccharide-responsive changes. Biochem. Biophys. Res. Commun. 2017, 486, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Esch, L.; Schaffrath, U. An update on jacalin-related lectins and their role in plant defense. Int. J. Mol. Sci. 2017, 18, 1592. [Google Scholar] [CrossRef]
- Keller, C.K.; Radwan, O. The functional role of 14-3-3 proteins in plant stress interactions. i-ACES 2015, 1, 100–110. [Google Scholar]
- Hayafune, M.; Berisio, R.; Marchetti, R.; Silipo, A.; Kayama, M.; Desaki, Y.; Arima, S.; Squeglia, F.; Ruggiero, A.; Tokuyasu, K.; et al. Chitin-induced activation of immune signaling by the rice receptor CEBiP relies on a unique sandwich-type dimerization. Proc. Natl. Acad. Sci. USA 2014, 111, E404–E413. [Google Scholar] [CrossRef] [Green Version]
- Simanshu, D.K.; Zhai, X.; Munch, D.; Hofius, D.; Markham, J.E.; Bielawski, J.; Bielawska, A.; Malinina, L.; Molotkovsky, J.G.; Mundy, J.W.; et al. Arabidopsis accelerated-cell-death11, ACD11, is a ceramide-1-phosphate transfer protein and intermediary regulator of phytoceramide levels. Cell Rep. 2014, 6, 388–399. [Google Scholar] [CrossRef]
- Liang, H.; Yao, N.; Song, J.T.; Luo, S.; Lu, H.; Greenberg, J.T. Ceramides modulate programmed cell death in plants. Genes Dev. 2003, 17, 2636–2641. [Google Scholar] [CrossRef] [Green Version]
- Varet, A.; Hause, B.; Hause, G.; Scheel, D.; Lee, J. The Arabidopsis NHL3 gene encodes a plasma membrane protein and its overexpression correlates with increased resistance to Pseudomonas syringae pv. tomato DC3000. Plant Physiol. 2003, 132, 2023–2033. [Google Scholar] [CrossRef]
- Elmore, J.M.; Liu, J.; Smith, B.; Phinney, B.; Coaker, G. Quantitative proteomics reveals dynamic changes in the plasma membrane during Arabidopsis immune signaling. Mol. Cell. Proteom. 2012, 11, M111–014555. [Google Scholar] [CrossRef]
- Raffaele, S.; Bayer, E.; Lafarge, D.; Cluzet, S.; Retana, S.G.; Boubekeur, T.; Leborgne-Castel, N.; Carde, J.-P.; Lherminier, J.; Noirot, E.; et al. Remorin, a Solanaceae protein in membrane rafts and plasmodesmata, impairs Potato virus X movement. Plant Cell 2009, 21, 1541–1555. [Google Scholar] [CrossRef]
- Marin, M.; Ott, T. Phosphorylation of intrinsically disordered regions in remorin proteins. Front. Plant Sci. 2012, 3, 86. [Google Scholar] [CrossRef]
- Zou, B.; Hong, X.; Ding, Y.; Wang, X.; Liu, H.; Hua, J. Identification and analysis of copine/BONZAI proteins among evolutionarily diverse plant series. Genome 2016, 59, 565–573. [Google Scholar] [CrossRef]
- Zhou, L.; Cheung, M.Y.; Li, M.-W.; Fu, Y.; Sun, Z.; Sun, S.-M.; Lam, H.-M. Rice hypersensitive induced reaction protein 1 (OsHIR1) associates with plasma membrane and triggers hypersensitive cell death. BMC Plant Biol. 2010, 10, 1–10. [Google Scholar] [CrossRef]
- Mohr, T.J.; Mammarelia, N.D.; Hoff, T.; Woffenden, B.J.; Jelesko, J.G.; McDowell, J.M. The Arabidopsis downy mildew resistance gene RPP8 is induced by pathogens and salicylic acid and is regulated by W box cis elements. Mol. Plant Microbe Interact. 2010, 23, 1303–1315. [Google Scholar] [CrossRef]
- Fairbanks, G.; Steck, T.L.; Wallach, D.F.H. Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 1971, 10, 2606–2617. [Google Scholar] [CrossRef]
- Jordaan, J.; Simpson, C.; Brady, D.; Gardiner, N.S.; Gerber, I.B. Emulsion-Derived Particles. U.S. Patent 2013149730 A1, 13 June 2013. [Google Scholar]
- Tejada-Simon, M.V.; Pestka, J.J. Production of polyclonal antibody against ergosterol hemisuccinate using Freund’s and Titermax adjuvants. J. Food Protect. 1998, 61, 1060–1063. [Google Scholar] [CrossRef]
- Switzer, R.C., III; Merril, C.R.; Shifrin, S. A highly sensitive silver stain for detecting proteins and peptides in polyacrylamide gels. Anal. Biochem. 1979, 98, 231–237. [Google Scholar] [CrossRef]
- Blum, H.; Beier, H.; Gross, H.J. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]
- UniProt Knowledgebase (UniprotKB) Database. Available online: https://www.uniprot.org/ (accessed on 2017–2018).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Protein Name | Accession No. | Biological GO Term | Molecular GO Term | Calculated Mass a (M + H) | Mass Error b (ppm) | Byonic ™ Score c | |Log Prob|d |
|---|---|---|---|---|---|---|---|---|
| Perception and signaling (17) | ||||||||
| A5 | Calcium-dependent lipid-binding (CaLB domain) family protein At3g61050 | Q9LEX1 | Response Signaling | DNA-binding | 1214.699 | −0.6 | 422.1 | 8.18 |
| A7 | Non-lysosomal glucosylceramidase At4g10060 | F4JLJ2 | Lipid Metabolism | Glycosidase | 1294.627 | −1.9 | 395.8 | 7.88 |
| A10 | G-type lectin S-receptor-like serine/threonine-protein kinase CES101 At3g16030 | Q9LW83 | Perception Response | Transferase | 1113.626 | −0.6 | 350.0 | 3.23 |
| A5 | Nicalin At3g44330 | Q9M292 | Signaling | --- | 1142.642 | 0.4 | 335.6 | 5.34 |
| A7, A12 | Cysteine-rich receptor-like protein kinase 41 At4g00970 | O23081 | Signaling | Transferase | 973.531 | 0.3 | 328.0 | 1.53 |
| A3 | Axi 1 protein-like protein At2g44500 | O64884 | Biosynthesis Metabolism | Transferase | 928.535 | −2.9 | 289.7 | 2.72 |
| A7 | Cysteine-rich receptor-like protein kinase 10 At4g23180 | Q8GYA4 | Signaling | Transferase | 1223.667 | 0.0 | 285.9 | 6.63 |
| A7 | PQQ_DH domain-containing protein At5g11560 | F4JXW9 | Biosynthesis | --- | 992.541 | 1.2 | 251.0 | 5.58 |
| A8 | Probable serine/threonine-protein kinase At4g35230 | Q944A7 | Defense | Transferase | 1269.741 | −2.3 | 236.2 | 6.31 |
| A4 | 14-3-3-like protein GF14 epsilon At1g22300 | P48347 | Signaling | Protein binding | 1229.580 | −1.5 | 230.0 | 5.62 |
| A9 | Phosphoinositide phospholipase C 2 At3g08510 | Q39033 | Defense | Hydrolase | 996.645 | −0.5 | 228.4 | 4.97 |
| A7 | AMP deaminase At2g38280 | O80452 | Response | Hydrolase | 1123.563 | 0.9 | 224.0 | 4.69 |
| A10 | Probable inactive leucine-rich repeat receptor-like protein kinase At3g03770 | Q8LFN2 | Signaling | Kinase | 1041.515 | 0.4 | 217.2 | 1.30 |
| A13 | Mitogen-activated protein kinase 8 At1g18150 | Q9LM33 | Signaling | Kinase | 1028.537 | 0.4 | 200.2 | 8.87 |
| A7 | Putative leucine-rich repeat receptor-like serine/threonine-protein kinase At2g24130 | Q9ZUI0 | Signaling | Transferase | 1149.626 | 2.2 | 174.2 | 1.02 |
| A7 | Leucine-rich repeat receptor-like protein kinase At2g01210 | Q9ZU46 | Signaling | Transferase | 870.541 | 0.1 | 164.6 | 0.9 |
| A7 | Receptor-like kinase TMK4 At3g23750 | Q9LK43 | Signaling | Kinase | 1020.572 | 0.6 | 121.5 | 1.15 |
| Membrane trafficking and transport (16) | ||||||||
| A5 | V-type proton ATPase subunit B2 At4g38510 | Q9SZN1 | Transport | Hydrolase | 1563.801 | −1.4 | 574.5 | 9.38 |
| A7 | Patellin-1 At1g72150 | Q56WK6 | Growth | Lipid binding | 1231.689 | −0.7 | 515.8 | 7.93 |
| A3 | Ras-related protein RABE1c At3g46060 | P28186 | Signaling Transport | GTPase | 1071.641 | −0.9 | 412.5 | 8.36 |
| A7 | ATPase 1, plasma membrane-type At2g18960 | P20649 | Transport | Translocase | 1040.574 | 0.5 | 401.7 | 7.98 |
| A6 | Ras-related protein RABA1g At3g15060 | Q9LK99 | Signaling Transport | GTPase | 1043.610 | −0.1 | 384.7 | 8.14 |
| A7 | Clathrin heavy chain 1 At3g11130 | Q0WNJ6 | Transport | Clathrin binding | 992.578 | 0.5 | 289.6 | 5.65 |
| A3, A7 | Probable aquaporin PIP1-5 At4g23400 | Q8LAA6 | Transport | Water transport | 1049.599 | −0.5 | 288.9 | 6.62 |
| A5 | Aquaporin PIP1-2 At2g45960 | Q06611 | Transport | Water transport | 1033.604 | −0.7 | 282.3 | 6.57 |
| A7 | CSC1-like protein ERD4 At1g30360 | Q9C8G5 | Transport | Ion channel | 1251.612 | 0.5 | 271.5 | 7.51 |
| A4 | Probable ADP, ATP carrier protein At5g56450 | Q9FM86 | Transport | ATP:ADP transport | 1021.531 | −0.3 | 254.1 | 5.24 |
| A3 | Ras-related protein RABA1e At4g18430 | O49513 | Signaling Transport | GTPase | 1274.612 | −1.4 | 240.9 | 7.40 |
| A8 | Aquaporin TIP1-2 At3g26520 | Q41963 | Transport | Water transport | 1980.030 | 0.0 | 239.9 | 6.69 |
| A4, A5, A8 | Aquaporin PIP2-1 At3g53420 | P43286 | Transport | Water transport | 1069.568 | 0.2 | 215.3 | 5.98 |
| A5 | Probable aquaporin PIP2-6 At2g39010 | Q9ZV07 | Transport | Water transport | 1311.669 | −0.8 | 214.5 | 1.65 |
| A7 | Exocyst complex component SEC3A At1g47550 | Q9SX85 | Transport | GTP-Rho binding | 1015.578 | −1.7 | 183.9 | 1.26 |
| A1 | Aluminum-activated malate transporter 6 At2g17470 | Q9SHM1 | Transport | Malate transporter | 1606.832 | 2.8 | 40 | 1.29 |
| Defense (6) | ||||||||
| A5 | Trans-cinnamate 4-monooxygenase At2g30490 | P92994 | Biosynthesis Defense | Monooygenase activity | 1271.721 | −0.3 | 377.3 | 8.02 |
| A9 | Protein BONZAI 2 At5g07300 | Q5S1W2 | Response | Phospholipid binding | 1199.663 | 0.2 | 340.2 | 7.66 |
| A3 | Temperature-induced lipocalin-1 At5g58070 | Q9FGT8 | Response | Storage protein | 1110.531 | −0.5 | 329.0 | 7.88 |
| A7 | Disease resistance protein RPP8 At5g43470 | Q8W4J9 | Defense | ATP:ADP binding | 1140.557 | −2.1 | 267.7 | 6.39 |
| A4 | Hypersensitive-induced response protein 3 At3g01290 | Q9SRH6 | Response | --- | 949.547 | −1.6 | 237.0 | 5.91 |
| A4 | Uncharacterized protein (LOW PSII ACCUMULATION-like protein) At4g28740 | F4JM22 | Chloroplast | --- | 995.600 | -0.1 | 131.8 | 1.24 |
| Structure (1) | ||||||||
| A2 | Putative clathrin assembly protein At1g14910 | P94017 | Transport | Clathrin binding | 1314.742 | −1.3 | 122.2 | 1.22 |
| Unknown (8) | ||||||||
| A11 | Triacylglycerol lipase-like 1 At1g45200 | Q8L7S1 | Metabolism | Hydrolase | 1222.622 | −0.2 | 336.6 | 6.54 |
| A11 | TNF receptor associated factor (TRAF)-like family protein At1g58270/F19C14_8 | Q9SLV3 | Signal transduction | ---- | 1434.722 | −0.8 | 286.2 | 6.92 |
| A7 | Uncharacterized protein At4g16180 | F4JLQ2 | --- | --- | 1293.669 | −2.2 | 235.3 | 5.38 |
| A1 | Putative uncharacterized protein At3g19340 | Q8RWC3 | --- | Aminopeptidase | 1219.632 | −0.9 | 229.5 | 5.94 |
| A12 | Putative uncharacterized protein F14P22.240 At3g58650 | Q9M2F2 | Growth | --- | 472.288 | −1.1 | 160.4 | 1.04 |
| A12 | Putative uncharacterized protein F3A4.21 At3g50130 | Q9SN05 | --- | --- | 472.288 | −1.1 | 160.4 | 0.92 |
| A2 | Uncharacterized protein At4g38260 | F4JTM0 | --- | --- | 1245.520 | −5.2 | 142.3 | 0.98 |
| A2 | EMB|CAB72473.1 At5g22560 | Q9FK83 | --- | --- | 1467.731 | 0.1 | 133.6 | 1.21 |
| Sample No. | Protein Name | Accession No. | Biological GO Term | Molecular GO Term | Calculated Mass a (M + H) | Mass Error b (ppm) | Byonic ™ Score c | |Log Prob| d |
|---|---|---|---|---|---|---|---|---|
| Perception and signaling (10) | ||||||||
| B7 | Probable serine/threonine-protein kinase At4g35230 | Q944A7 | Signaling | Transferase | 1269.741 | 0.1 | 480.6 | 7.29 |
| B4 | At2g34560 protein (P-loop containing nucleoside triphosphate hydrolase) At2g34560 | B9DGC0 | Transport | ATPactivity | 1156.672 | −0.8 | 401.3 | 8.95 |
| B3 | Aspartyl aminopeptidase At5g60160/f15|12_20 | Q9LST0 | Biosynthesis | Metalloaminopeptidase | 1148.679 | 0.3 | 382.0 | 8.65 |
| B8 | Probable protein phosphatase 2C 20 At2g20630 | Q9SIU8 | Signaling | Hydrolase | 1288.711 | −0.1 | 363.1 | 7.51 |
| B6 | Abscisic acid receptor PYL1 At5g46790 | Q8VZS8 | Signaling | Receptor | 1442.760 | −0.5 | 357.2 | 9.01 |
| B7 | Phosphotidylinositol 4-kinase alpha 1 At1g49340 | Q9SXA1 | Signaling | Kinase | 1964.041 | 0.1 | 345.5 | 6.85 |
| B2 | Protein SGT1 homolog B At4g11260 | Q9SUT5 | Signaling | --- | 1435.709 | −1.4 | 351.9 | 8.86 |
| B1 | Fasciclin-like arabinogalactan protein 7 At2g04780 | Q9SJ81 | Biosynthesis | --- | 981.500 | 0.9 | 322.0 | 6.49 |
| B2 | 1-Phosphotidylinositol-3-phosphate 5-kinase FAB1A At4g33240 | Q0WUR5 | Signaling | Kinase | 1470.816 | −1.8 | 303.8 | 7.75 |
| B8 | Plasma membrane-associated cation-binding protein 1 At4g20260 | Q96262 | Response | Ion binding | 1146.641 | 0.0 | 281.2 | 7.00 |
| Membrane trafficking and transport (16) | ||||||||
| B2 | V-type proton ATPase subunit B3 At1g20260 | Q8W4E2 | Transport | Hydrolase | 1563.801 | −1.9 | 442.2 | 9.76 |
| B8 | Alpha-soluble NSF attachment protein 2 At3g56190 | Q9SPE6 | Transport | --- | 1259.684 | −0.8 | 426.3 | 8.35 |
| B6 | Ras-related protein RABA1d At4g18800 | Q9SN35 | Signaling | GTPase | 1043.610 | 0.0 | 414.7 | 7.96 |
| B4, B7 | Patellin-2 At1g22530 | Q56ZI2 | Transport | Lipid-binding | 1520.784 | −0.7 | 391.9 | 9.24 |
| B4, B7 | Patellin-1 At1g72150 | Q56WK6 | Transport | Lipid-binding | 1231.689 | −2.0 | 372.2 | 5.53 |
| B2 | Clathrin light chain 3 At3g51890 | F4J5M9 | Transport | Clathrin binding | 855.530 | 0.0 | 363.4 | 4.96 |
| B6 | Ras-related protein RABA5b At3g07410 | Q9SRS5 | Signaling | GTPase | 1071.641 | −0.9 | 357.3 | 8.16 |
| B3 | SNAP25 homologous protein SNAP33 At5g61210 | Q9S7P9 | Transport | SNAP receptor | 1302.715 | −1.3 | 352.6 | 6.82 |
| B1, B4, B8 | V-type ATPase catalytic subunit A At1g78900 | O23654 | Transport | Hydrolase | 1019.552 | −1.6 | 338.6 | 5.50 |
| B7 | Sugar transport protein 7 At4g02050 | O04249 | Transport | Transmembrane transporter | 1006.469 | 0.9 | 338.1 | 6.27 |
| B3 | Auxin transport protein BIG At3g02260 | Q9SRU2 | Signaling | Zinc binding | 589.356 | −1.4 | 331.2 | 5.39 |
| B3 | Protein NETWORKED 1C At4g02710 | Q9ZQX8 | --- | Actin binding | 478.251 | 0.1 | 311.4 | 6.18 |
| B3 | ABC transporter C family member 8 At3g21250 | Q8LGU1 | Transport | Translocase | 530.330 | −0.3 | 303.2 | 5.82 |
| B8 | Syntaxin-71 At3g09740 | Q9SF29 | Transport | SNAP receptor | 1081.636 | 0.7 | 299.7 | 7.30 |
| B4, B7 | Flotillin-like protein 1 At5g25250 | Q501E6 | Transport | --- | 1526.909 | −1.8 | 273.6 | 6.93 |
| Defense (9) | ||||||||
| B2, B4, B7 | Jacalin-related lectin 35 At3g16470 | O04309 | Perception Response | Carbohydrate binding | 1469.763 | −2.5 | 518.7 | 9.21 |
| B6 | Aluminium induced protein with YGL and LRDR motifs At5g19140 | Q94BR2 | --- | --- | 1439.738 | −1.6 | 420.4 | 8.77 |
| B6, B8 | At3g11930 protein (Adenine nucleotide alpha hydrolases-like) At3g11930 | B9DG73 | --- | Hydrolase | 1189.631 | −2.2 | 380.6 | 8.02 |
| B4 | Callose synthase 9 At3g07160 | Q9SFU6 | Biosynthesis Defense | Transferase | 557.402 | −0.9 | 334.8 | 2.44 |
| B8 | Hypersensitive-induced response protein 4 At5g51570 | Q9FHM7 | Defense Signaling | --- | 1466.764 | 1.7 | 345.6 | 8.36 |
| B8 | Binding partner of ACD (accelerated cell death)11 1 At5g16840 | Q9LFD5 | Signaling | RNA-binding | 1132.621 | −1.0 | 332.1 | 7.69 |
| B5, B8 | Hypersensitive-induced response protein 2 At1g69840 | Q9CAR7 | Defense Signaling | Kinase binding | 871.500 | −1.3 | 327.2 | 4.57 |
| B6 | Dessication responsive protein At2g21620 | Q94II5 | --- | Hydrolase | 980.614 | −0.8 | 293.5 | 6.84 |
| B8 | Hypersensitive-induced response protein 1 At5g62740 | Q9FM19 | Defense Signaling | Kinase-binding | 949.547 | −0.5 | 281.6 | 7.29 |
| Sample No. | Protein Name | Accession No. | Biological GO Term | Molecular GO Term | Calculated Mass a (M + H) | Mass Error b (ppm) | Byonic™ Score c | |Log Prob|d |
|---|---|---|---|---|---|---|---|---|
| Signaling | ||||||||
| A12 | Uncharacterized glycosylphophatidylinositol (GPI)-anchored protein At5g19250 | P59833 | --- | --- | 1910.898 | −3.1 | 464.4 | 8.69 |
| A13 | Binding partner of ACD (accelerated cell death)11 1 At5g16840 | Q9LFD5 | Signaling Response | RNA-binding | 1132.621 | −1.0 | 428.4 | 8.21 |
| A4 | Probable inactive receptor kinase At3g02880 | Q9M8T0 | Response | Receptor | 1426.706 | −3.0 | 392.2 | 6.99 |
| A13 | Uncharacterized GPI-anchored protein At5g19250 | P59833 | --- | --- | 1910.898 | −3.5 | 388.2 | 7.32 |
| A11 | Leucine-rich repeat-containing protein At5g07910 | Q8RWI2 | Response | --- | 1269.727 | −0.5 | 336.8 | 7.18 |
| A4 | Probable inactive receptor kinase At5g16590 | Q9FMD7 | Response | Receptor | 2127.170 | −1.5 | 324.5 | 8.27 |
| A11 | Leucine-rich repeat protein kinase-like protein At1g10850 | Q940B9 | Signaling Response | Kinase | 984.584 | 1.0 | 322.0 | 6.56 |
| A11, A14 | Chitin elicitor receptor kinase 1 At3g21630 | A8R7E6 | Perception Signaling | Kinase | 1132.596 | −0.6 | 313.8 | 6.87 |
| Membrane trafficking and transport | ||||||||
| A1, A5 | Aquaporin PIP2-7 At4g35100 | P93004 | Transport | Water channel | 1312.653 | −1.4 | 541.6 | 7.44 |
| A1 | Aquaporin PIP1-2 At2g45960 | Q06611 | Transport | Water channel | 1017.548 | −0.3 | 509.0 | 6.59 |
| A1 | Aquaporin PIP2-1 At3g53420 | P43286 | Transport | Water channel | 2000.996 | −1.2 | 473.2 | 10.44 |
| A1 | Probable aquaporin PIP1-5 At4g23400 | Q8LAA6 | Transport | Water channel | 1230.632 | −1.1 | 464.3 | 9.05 |
| A8 | Plasma membrane-associated cation-binding protein 1 At4g20260 | Q96262 | Response | Ion-binding | 1425.711 | −2.6 | 418.8 | 8.56 |
| A4 | ATPase 2, plasma membrane-type At4g30190 | P19456 | Transport | Translocase | 1040.574 | −0.2 | 412.0 | 7.60 |
| A6 | At2g34250 protein At2g34250 | O80774 | Transport | Protein transport | 1164.601 | −2.0 | 408.2 | 6.50 |
| A1 | Ras-related protein RABE1c At3g46060 | P28186 | Signaling | GTPase | 1071.641 | −0.7 | 394.8 | 8.40 |
| A9, A12 | CASP-like protein 1D1 At4g15610 | Q9FE29 | --- | --- | 1127.657 | −0.3 | 389.4 | 9.41 |
| A11 | Plasma membrane ATPase At4g30190 | F4JPJ7 | Transport | Translocase | 1040.574 | −0.2 | 381.3 | 8.04 |
| A11 | ATPase 5, plasma membrane-type At2g24520 | Q9SJB3 | Transport | Translocase | 1040.574 | 0.0 | 362.8 | 7.00 |
| A11 | Fasciclin-like arabinogalactan protein 8 At2g45470 | O22126 | --- | --- | 967.484 | −0.8 | 348.8 | 7.71 |
| A11 | Patellin-1 At1g72150 | Q56WK6 | Transport | Lipid-binding | 1078.589 | −1.2 | 326.8 | 7.67 |
| A1 | F-box/LRR-repeat protein At3g60040 | Q8GWI2 | Response | --- | 784.529 | −0.3 | 309.1 | 1.05 |
| A9 | ABC transporter G family member 41 At4g15215 | Q7PC83 | Transport | ATP-binding | 543.386 | −0.9 | 301.8 | 4.38 |
| A6 | CSC1-like protein ERD4 At1g30360 | Q9C8G5 | Transport | Ion channel | 1583.850 | 2.7 | 301.4 | 7.40 |
| Defense response | ||||||||
| A5 | Hypersensitive-induced response protein 3 At3g01290 | Q9SRH6 | Defense Signaling Response | --- | 1519.775 | −1.6 | 489.0 | 8.32 |
| A6 | Syntaxin-121 At3g11820 | Q9ZSD4 | Defense | SNAP receptor | 1329.701 | −1.4 | 439.9 | 7.40 |
| A10 | NDR1/HIN1-like protein 3 At5g06320 | Q9FNH6 | Defense | --- | 1496.843 | −1.8 | 394.3 | 8.73 |
| A10 | Protein BONZAI 2 At5g07300 | Q5S1W2 | Defense | Phospholipid-binding | 1060.615 | 0.0 | 372.4 | 7.25 |
| A11, A14 | Remorin At2g45820 | O80837 | --- | --- | 617.409 | −1.8 | 358.2 | 4.82 |
| A11, A14 | Blue copper protein At5g20230 | Q07488 | Transport | Electron transfer | 1425.664 | −0.4 | 337.8 | 9.34 |
| Unknown | ||||||||
| A11 | At1g55160/T7N22.11 At1g55160 | Q9C542 | --- | --- | 1174.631 | −0.2 | 384.7 | 8.38 |
| A11 | At3g08600/F17014_7 At3g08600 | Q9C9Z6 | --- | --- | 903.453 | −0.2 | 352.4 | 7.93 |
| A14 | Expressed protein At2g18690 | Q9ZV49 | --- | --- | 1115.606 | −1.0 | 319.2 | 6.03 |
| Sample No. | Protein Name | Accession No. | Biological GO Term | Molecular GO Term | Calculated Mass a (M + H) | Mass Error b (ppm) | Byonic™ Score c | |Log Prob|d |
|---|---|---|---|---|---|---|---|---|
| A7 | Ras-related protein RABG1 At5g39620 | Q948K6 | Signaling | GTP-binding | 1071.641 | 0.0 | 515.00 | 6.92 |
| A4 | Aquaporin PIP1-2 At2g45960 | Q06611 | Transport | Water channel | 1033.604 | −1.2 | 336.3 | 6.55 |
| A10 | 1-Phosphotidylinositol-3-phosphate-5-kinase FAB1B At3g14270 | Q9LUM0 | --- | Kinase | 956.480 | −0.5 | 328.4 | 6.19 |
| A6 | Ras-related protein RABE1c At3g46060 | P28186 | Signaling | GTP-binding | 1164.590 | 0.6 | 319.2 | 6.34 |
| A1 | Aquaporin PIP2-1 At3g53420 | P43286 | Transport | Water channel | 1069.568 | 0.4 | 284.5 | 5.82 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khoza, T.G.; Dubery, I.A.; Piater, L.A. Identification of Candidate Ergosterol-Responsive Proteins Associated with the Plasma Membrane of Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 1302. https://doi.org/10.3390/ijms20061302
Khoza TG, Dubery IA, Piater LA. Identification of Candidate Ergosterol-Responsive Proteins Associated with the Plasma Membrane of Arabidopsis thaliana. International Journal of Molecular Sciences. 2019; 20(6):1302. https://doi.org/10.3390/ijms20061302
Chicago/Turabian StyleKhoza, Thembisile G., Ian A. Dubery, and Lizelle A. Piater. 2019. "Identification of Candidate Ergosterol-Responsive Proteins Associated with the Plasma Membrane of Arabidopsis thaliana" International Journal of Molecular Sciences 20, no. 6: 1302. https://doi.org/10.3390/ijms20061302