Transcriptional Regulation Factors of the Human Mitochondrial Aspartate/Glutamate Carrier Gene, Isoform 2 (SLC25A13): USF1 as Basal Factor and FOXA2 as Activator in Liver Cells

 , ,
, ,
Abstract
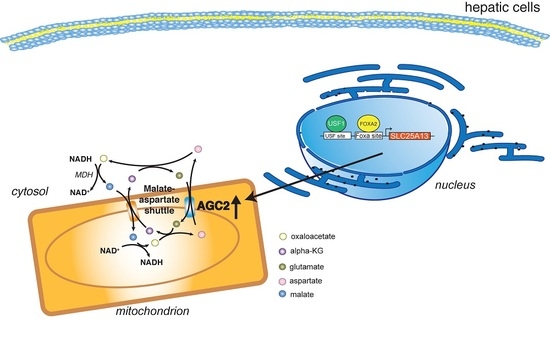
:
1. Introduction
2. Results
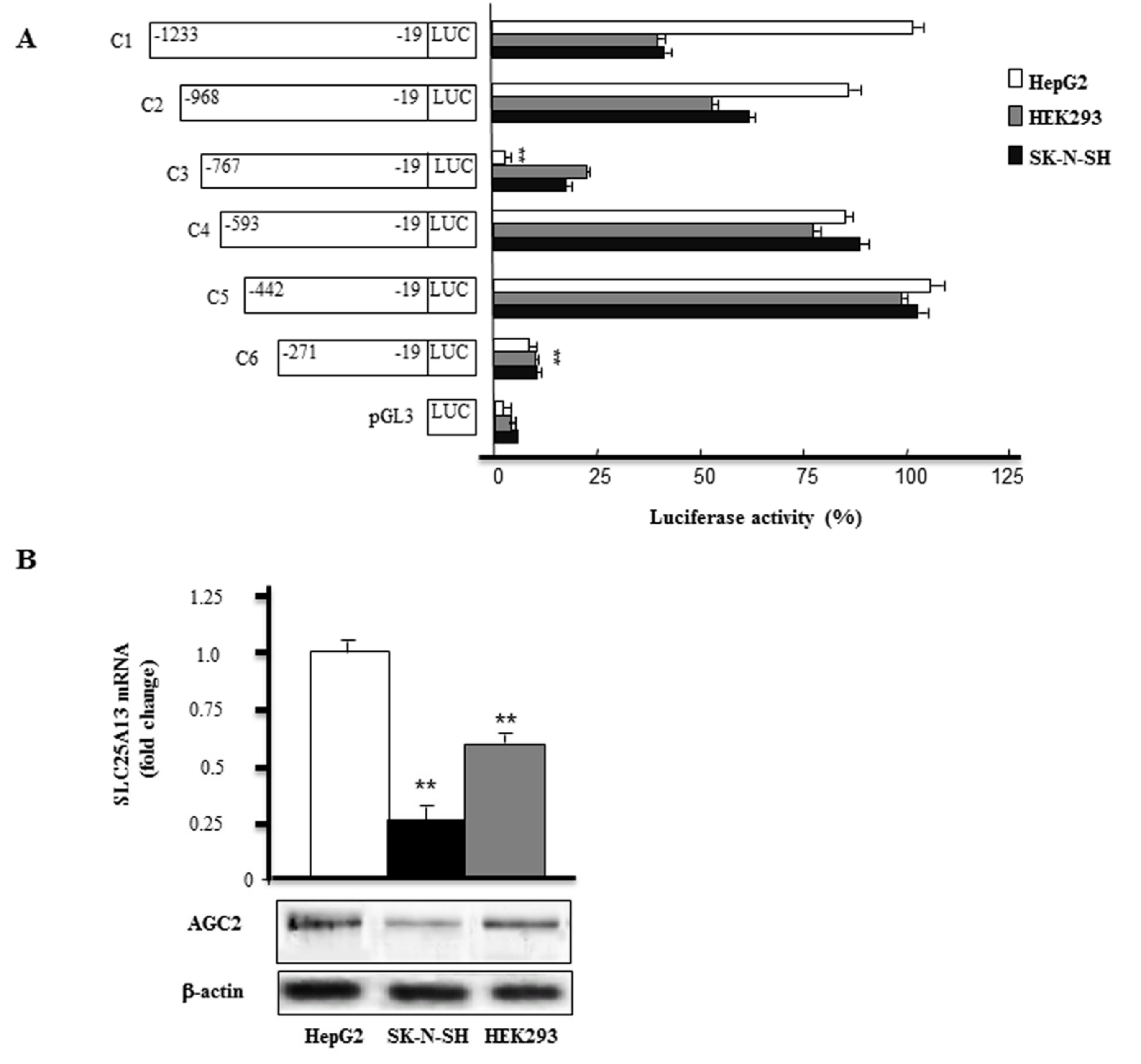
2.1. Screening of SLC25A13 Gene Promoter Activity
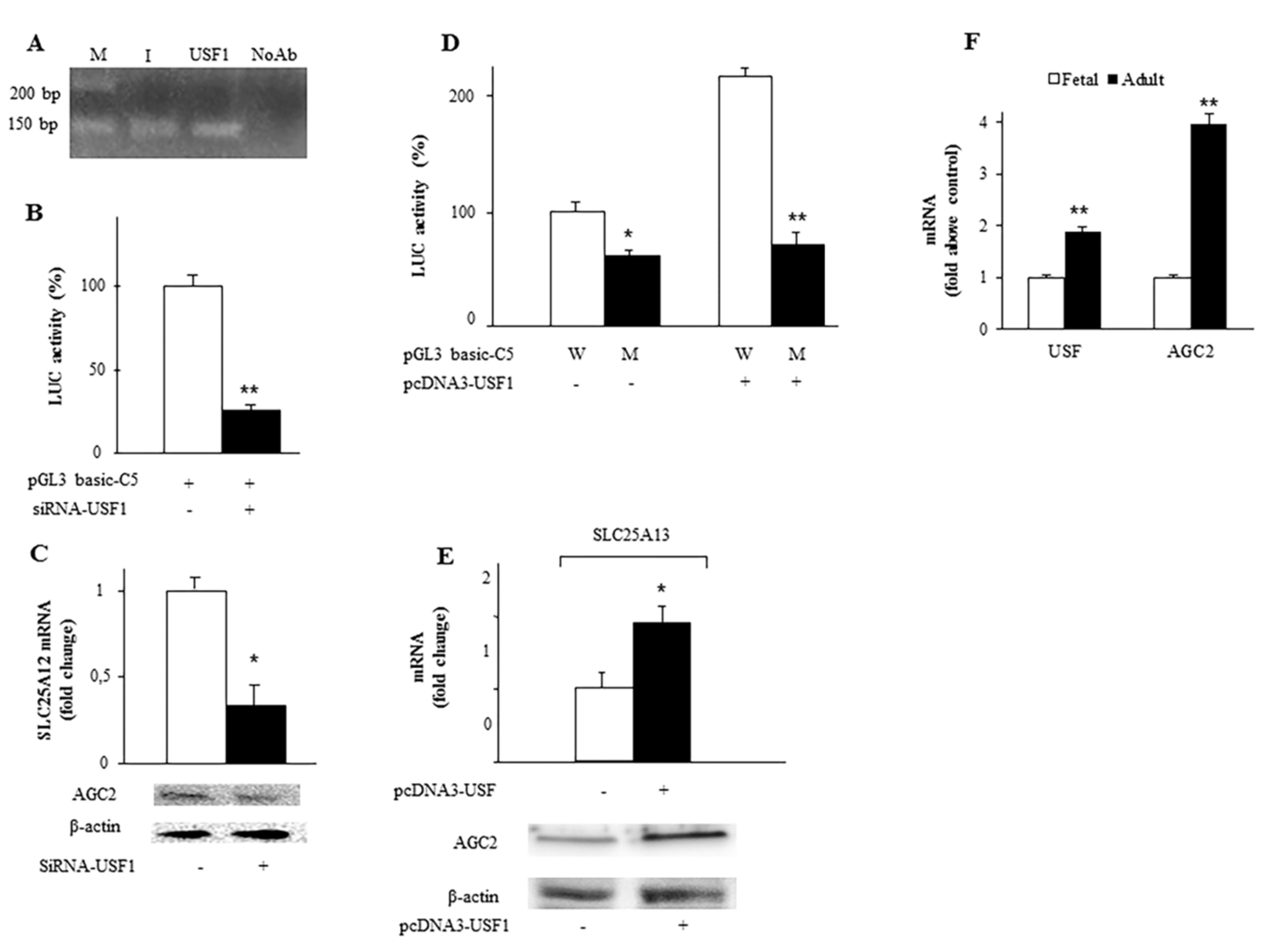
2.2. Identification and Functionality of the USF1 Cis-Element in SLC25A13 Gene Promoter
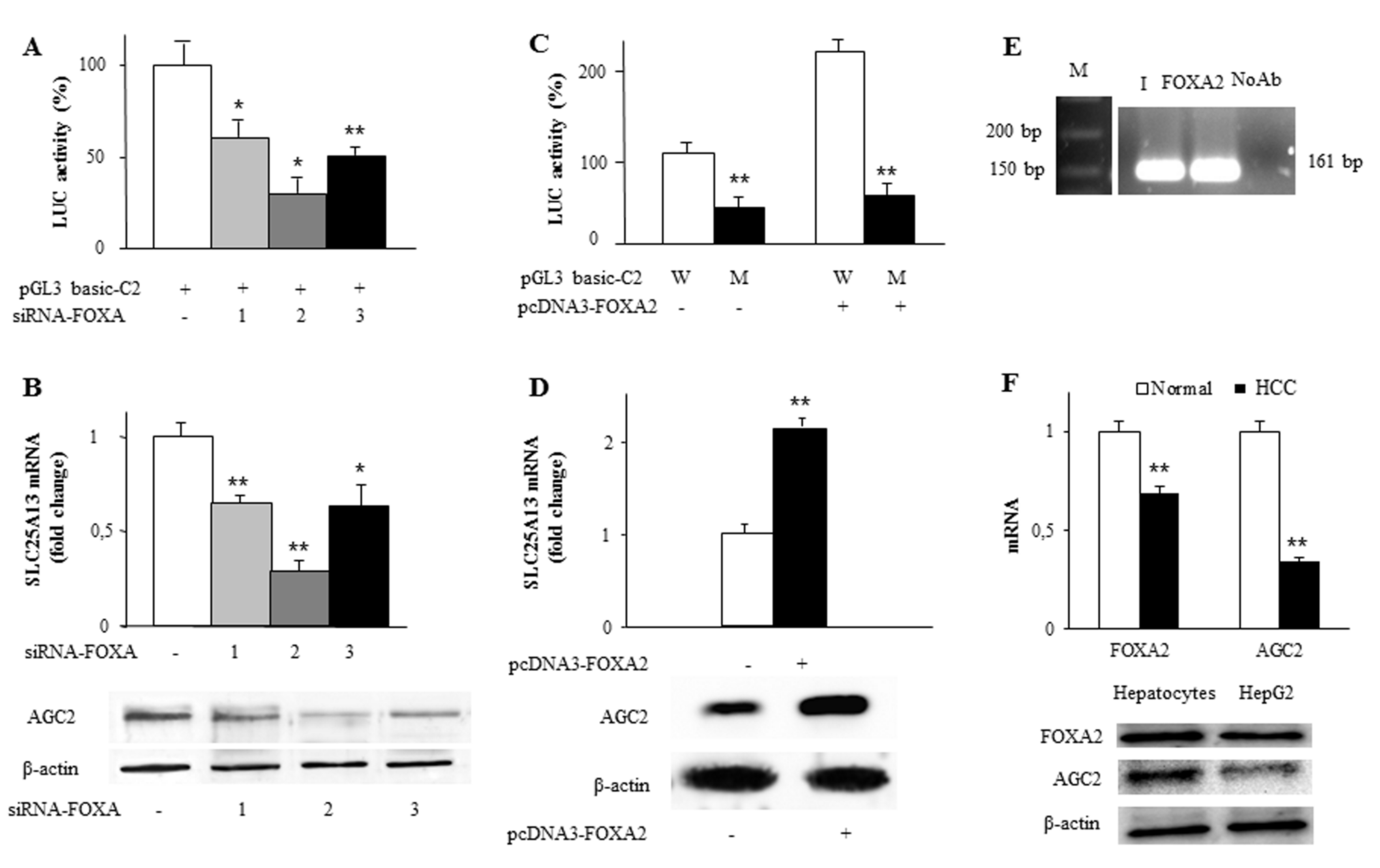
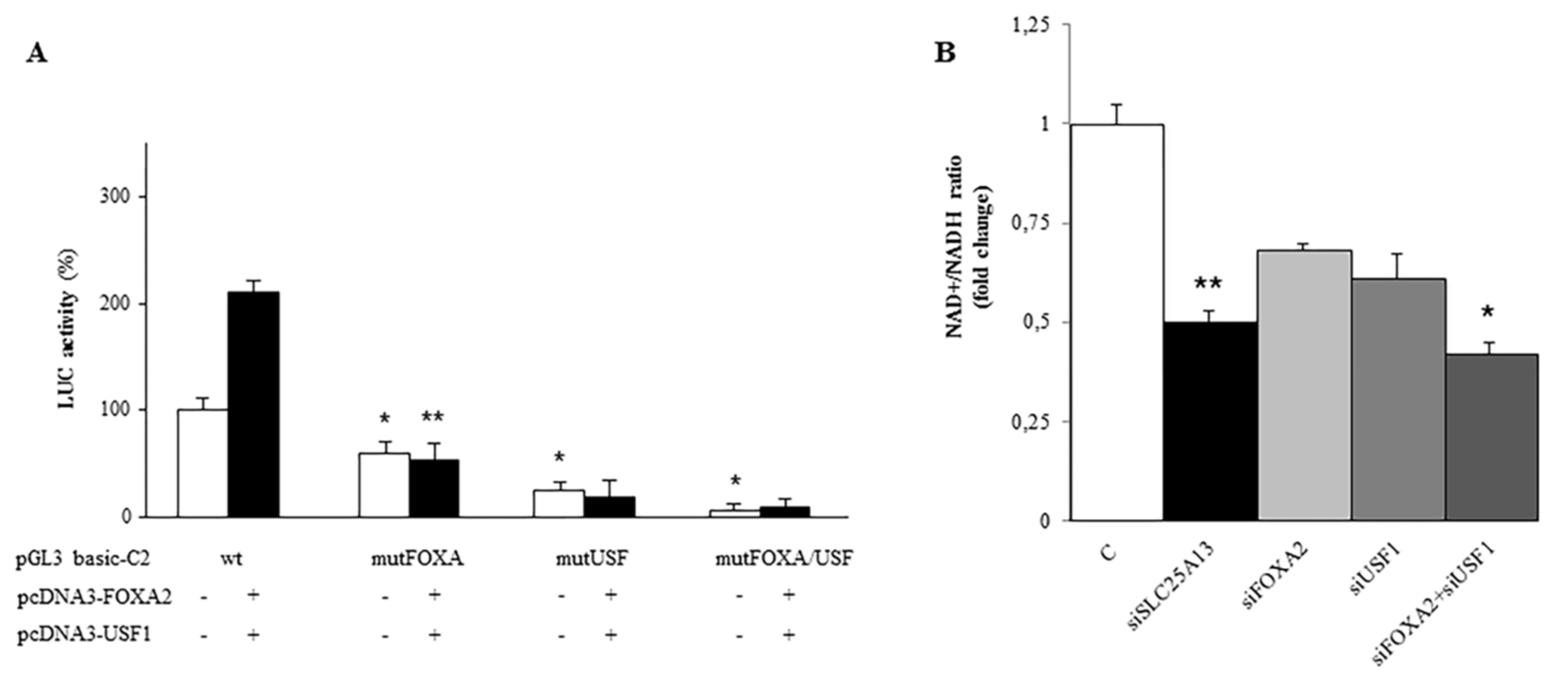
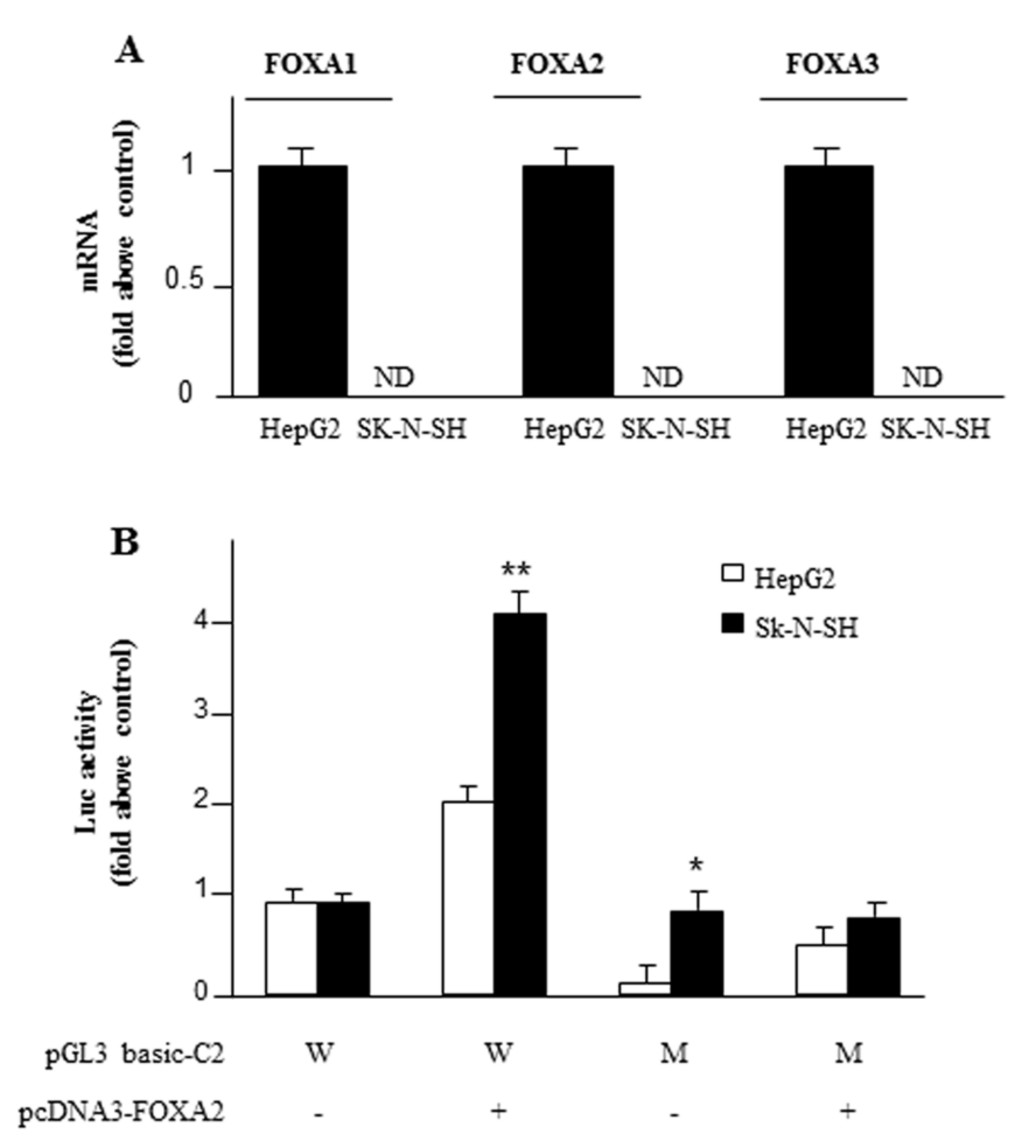
2.3. Identification and Functionality of the FOXA2 Cis-Element in SLC25A13 Gene Promoter
2.4. Exogenous FOXA2 Induces Transcription of SLC25A13 in SK-N-SH Cells
2.5. Synergy between FOXA2 and USF1 in Regulating SLC25A13 Gene Expression and AGC2 Function
3. Discussion
4. Materials and Methods
4.1. Construction of Plasmids.
4.2. Cell Culture, RNA Interference and Transient Transfection.
4.3. Reverse Transcriptase-PCR and Real-Time PCR
4.4. Western Blotting
4.5. Chromatin Immunoprecipitation
4.6. Intracellular NAD+/NADH Quantification
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Palmieri, F. The mitochondrial transporter family slc25: Identification, properties and physiopathology. Mol. Aspects Med. 2013, 34, 465–484. [Google Scholar] [CrossRef]
- Palmieri, F. Mitochondrial transporters of the slc25 family and associated diseases: A review. J. Inherit. Metab. Dis. 2014, 37, 565–575. [Google Scholar] [CrossRef]
- Palmieri, F.; Pierri, C.L. Mitochondrial metabolite transport. Essays Biochem. 2010, 47, 37–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, F.; Monne, M. Discoveries, metabolic roles and diseases of mitochondrial carriers: A review. Biochim. Biophys. Acta 2016, 1863, 2362–2378. [Google Scholar] [CrossRef]
- Palmieri, L.; Pardo, B.; Lasorsa, F.M.; del Arco, A.; Kobayashi, K.; Iijima, M.; Runswick, M.J.; Walker, J.E.; Saheki, T.; Satrustegui, J.; et al. Citrin and aralar1 are Ca(2+)-stimulated aspartate/glutamate transporters in mitochondria. EMBO J. 2001, 20, 5060–5069. [Google Scholar] [CrossRef]
- Indiveri, C.; Kramer, R.; Palmieri, F. Reconstitution of the malate/aspartate shuttle from mitochondria. J. Biol. Chem. 1987, 262, 15979–15983. [Google Scholar] [PubMed]
- Palmieri, F. The mitochondrial transporter family (slc25): Physiological and pathological implications. Pflugers Arch. 2004, 447, 689–709. [Google Scholar] [CrossRef]
- Lasorsa, F.M.; Pinton, P.; Palmieri, L.; Fiermonte, G.; Rizzuto, R.; Palmieri, F. Recombinant expression of the Ca(2+)-sensitive aspartate/glutamate carrier increases mitochondrial ATP production in agonist-stimulated Chinese hamster ovary cells. J. Biol. Chem. 2003, 278, 38686–38692. [Google Scholar] [CrossRef] [PubMed]
- Thangaratnarajah, C.; Ruprecht, J.J.; Kunji, E.R. Calcium-induced conformational changes of the regulatory domain of human mitochondrial aspartate/glutamate carriers. Nat. Commun. 2014, 5, 5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiermonte, G.; De Leonardis, F.; Todisco, S.; Palmieri, L.; Lasorsa, F.M.; Palmieri, F. Identification of the mitochondrial atp-mg/pi transporter. Bacterial expression, reconstitution, functional characterization and tissue distribution. J. Biol. Chem. 2004, 279, 30722–30730. [Google Scholar] [CrossRef]
- Cavero, S.; Traba, J.; Del Arco, A.; Satrustegui, J. The calcium-dependent atp-mg/pi mitochondrial carrier is a target of glucose-induced calcium signalling in saccharomyces cerevisiae. Biochem. J. 2005, 392, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Monne, M.; Miniero, D.V.; Obata, T.; Daddabbo, L.; Palmieri, L.; Vozza, A.; Nicolardi, M.C.; Fernie, A.R.; Palmieri, F. Functional characterization and organ distribution of three mitochondrial atp-mg/pi carriers in arabidopsis thaliana. Biochim. Biophys. Acta 2015, 1847, 1220–1230. [Google Scholar] [CrossRef] [PubMed]
- Monne, M.; Daddabbo, L.; Giannossa, L.C.; Nicolardi, M.C.; Palmieri, L.; Miniero, D.V.; Mangone, A.; Palmieri, F. Mitochondrial atp-mg/phosphate carriers transport divalent inorganic cations in complex with atp. J. Bioenerg. Biomembr. 2017, 49, 369–380. [Google Scholar] [CrossRef]
- Iijima, M.; Jalil, A.; Begum, L.; Yasuda, T.; Yamaguchi, N.; Xian Li, M.; Kawada, N.; Endou, H.; Kobayashi, K.; Saheki, T. Pathogenesis of adult-onset type ii citrullinemia caused by deficiency of citrin, a mitochondrial solute carrier protein: Tissue and subcellular localization of citrin. Adv. Enzyme Regul. 2001, 41, 325–342. [Google Scholar] [CrossRef]
- Del Arco, A.; Morcillo, J.; Martinez-Morales, J.R.; Galian, C.; Martos, V.; Bovolenta, P.; Satrustegui, J. Expression of the aspartate/glutamate mitochondrial carriers aralar1 and citrin during development and in adult rat tissues. Eur. J. Biochem. 2002, 269, 3313–3320. [Google Scholar] [CrossRef] [Green Version]
- Wibom, R.; Lasorsa, F.M.; Tohonen, V.; Barbaro, M.; Sterky, F.H.; Kucinski, T.; Naess, K.; Jonsson, M.; Pierri, C.L.; Palmieri, F.; et al. Agc1 deficiency associated with global cerebral hypomyelination. N. Engl. J. Med. 2009, 361, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Falk, M.J.; Li, D.; Gai, X.; McCormick, E.; Place, E.; Lasorsa, F.M.; Otieno, F.G.; Hou, C.; Kim, C.E.; Abdel-Magid, N.; et al. Agc1 deficiency causes infantile epilepsy, abnormal myelination and reduced n-acetylaspartate. JIMD Rep. 2014, 14, 77–85. [Google Scholar] [PubMed]
- Saheki, T.; Kobayashi, K.; Iijima, M.; Horiuchi, M.; Begum, L.; Jalil, M.A.; Li, M.X.; Lu, Y.B.; Ushikai, M.; Tabata, A.; et al. Adult-onset type ii citrullinemia and idiopathic neonatal hepatitis caused by citrin deficiency: Involvement of the aspartate glutamate carrier for urea synthesis and maintenance of the urea cycle. Mol. Genet. Metab. 2004, 81 (Suppl. 1), S20–S26. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Bang Lu, Y.; Xian Li, M.; Nishi, I.; Hsiao, K.J.; Choeh, K.; Yang, Y.; Hwu, W.L.; Reichardt, J.K.; Palmieri, F.; et al. Screening of nine slc25a13 mutations: Their frequency in patients with citrin deficiency and high carrier rates in asian populations. Mol. Genet. Metab. 2003, 80, 356–359. [Google Scholar] [CrossRef]
- Fiermonte, G.; Soon, D.; Chaudhuri, A.; Paradies, E.; Lee, P.J.; Krywawych, S.; Palmieri, F.; Lachmann, R.H. An adult with type 2 citrullinemia presenting in Europe. N. Engl. J. Med. 2008, 358, 1408–1409. [Google Scholar] [CrossRef]
- Jalil, M.A.; Begum, L.; Contreras, L.; Pardo, B.; Iijima, M.; Li, M.X.; Ramos, M.; Marmol, P.; Horiuchi, M.; Shimotsu, K.; et al. Reduced n-acetylaspartate levels in mice lacking aralar, a brain- and muscle-type mitochondrial aspartate-glutamate carrier. J. Biol. Chem. 2005, 280, 31333–31339. [Google Scholar] [CrossRef]
- Infantino, V.; Dituri, F.; Convertini, P.; Santarsiero, A.; Palmieri, F.; Todisco, S.; Mancarella, S.; Giannelli, G.; Iacobazzi, V. Epigenetic upregulation and functional role of the mitochondrial aspartate/glutamate carrier isoform 1 in hepatocellular carcinoma. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Iacobazzi, V.; Infantino, V.; Costanzo, P.; Izzo, P.; Palmieri, F. Functional analysis of the promoter of the mitochondrial phosphate carrier human gene: Identification of activator and repressor elements and their transcription factors. Biochem. J. 2005, 391, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Convertini, P.; Infantino, V.; Bisaccia, F.; Palmieri, F.; Iacobazzi, V. Role of foxa and sp1 in mitochondrial acylcarnitine carrier gene expression in different cell lines. Biochem. Biophys. Res. Commun. 2011, 404, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Iacobazzi, V.; Menga, A.; Avantaggiati, M.L.; Palmieri, F. A key role of the mitochondrial citrate carrier (slc25a1) in tnfalpha- and ifngamma-triggered inflammation. Biochim. Biophys. Acta 2014, 1839, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Convertini, P.; Cucci, L.; Panaro, M.A.; Di Noia, M.A.; Calvello, R.; Palmieri, F.; Iacobazzi, V. The mitochondrial citrate carrier: A new player in inflammation. Biochem. J. 2011, 438, 433–436. [Google Scholar] [CrossRef]
- Convertini, P.; Menga, A.; Andria, G.; Scala, I.; Santarsiero, A.; Castiglione Morelli, M.A.; Iacobazzi, V.; Infantino, V. The contribution of the citrate pathway to oxidative stress in down syndrome. Immunology 2016, 149, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Menga, A.; Palmieri, E.M.; Cianciulli, A.; Infantino, V.; Mazzone, M.; Scilimati, A.; Palmieri, F.; Castegna, A.; Iacobazzi, V. Slc25a26 overexpression impairs cell function via mtdna hypermethylation and rewiring of methyl metabolism. FEBS J. 2017, 284, 967–984. [Google Scholar] [CrossRef]
- Fiore, C.; Trezeguet, V.; Le Saux, A.; Roux, P.; Schwimmer, C.; Dianoux, A.C.; Noel, F.; Lauquin, G.J.; Brandolin, G.; Vignais, P.V. The mitochondrial adp/atp carrier: Structural, physiological and pathological aspects. Biochimie 1998, 80, 137–150. [Google Scholar] [CrossRef]
- Dolce, V.; Scarcia, P.; Iacopetta, D.; Palmieri, F. A fourth adp/atp carrier isoform in man: Identification, bacterial expression, functional characterization and tissue distribution. FEBS Lett. 2005, 579, 633–637. [Google Scholar] [CrossRef]
- Villarroya, F.; Peyrou, M.; Giralt, M. Transcriptional regulation of the uncoupling protein-1 gene. Biochimie 2017, 134, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Menga, A.; Iacobazzi, V.; Infantino, V.; Avantaggiati, M.L.; Palmieri, F. The mitochondrial aspartate/glutamate carrier isoform 1 gene expression is regulated by creb in neuronal cells. Int. J. Biochem. Cell Biol. 2015, 60, 157–166. [Google Scholar] [CrossRef]
- Anantharaman, A.; Lin, I.J.; Barrow, J.; Liang, S.Y.; Masannat, J.; Strouboulis, J.; Huang, S.; Bungert, J. Role of helix-loop-helix proteins during differentiation of erythroid cells. Mol. Cell. Biol. 2011, 31, 1332–1343. [Google Scholar] [CrossRef]
- Li, N.; Seetharam, B. A 69-base pair fragment derived from human transcobalamin ii promoter is sufficient for high bidirectional activity in the absence of a tata box and an initiator element in transfected cells. Role of an e box in transcriptional activity. J. Biol. Chem. 1998, 273, 28170–28177. [Google Scholar] [CrossRef] [PubMed]
- Kaestner, K.H. The hepatocyte nuclear factor 3 (hnf3 or foxa) family in metabolism. Trends Endocrinol. Metab. 2000, 11, 281–285. [Google Scholar] [CrossRef]
- Monaghan, A.P.; Kaestner, K.H.; Grau, E.; Schutz, G. Postimplantation expression patterns indicate a role for the mouse forkhead/hnf-3 alpha, beta and gamma genes in determination of the definitive endoderm, chordamesoderm and neuroectoderm. Development 1993, 119, 567–578. [Google Scholar] [PubMed]
- Wang, J.; Zhu, C.P.; Hu, P.F.; Qian, H.; Ning, B.F.; Zhang, Q.; Chen, F.; Liu, J.; Shi, B.; Zhang, X.; et al. Foxa2 suppresses the metastasis of hepatocellular carcinoma partially through matrix metalloproteinase-9 inhibition. Carcinogenesis 2014, 35, 2576–2583. [Google Scholar] [CrossRef]
- Kadesch, T. Consequences of heteromeric interactions among helix-loop-helix proteins. Cell Growth Differ. 1993, 4, 49–55. [Google Scholar]
- Roy, A.L.; Meisterernst, M.; Pognonec, P.; Roeder, R.G. Cooperative interaction of an initiator-binding transcription initiation factor and the helix-loop-helix activator usf. Nature 1991, 354, 245–248. [Google Scholar] [CrossRef]
- Corre, S.; Galibert, M.D. USF as a key regulatory element of gene expression. Med. Sci. 2006, 22, 62–67. [Google Scholar]
- Van Deursen, D.; van Leeuwen, M.; Vaulont, S.; Jansen, H.; Verhoeven, A.J. Upstream stimulatory factors 1 and 2 activate the human hepatic lipase promoter via e-box dependent and independent mechanisms. Biochim. Biophys. Acta 2009, 1791, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Bidder, M.; Shao, J.S.; Charlton-Kachigian, N.; Loewy, A.P.; Semenkovich, C.F.; Towler, D.A. Osteopontin transcription in aortic vascular smooth muscle cells is controlled by glucose-regulated upstream stimulatory factor and activator protein-1 activities. J. Biol. Chem. 2002, 277, 44485–44496. [Google Scholar] [CrossRef] [PubMed]
- Weigert, C.; Brodbeck, K.; Sawadogo, M.; Haring, H.U.; Schleicher, E.D. Upstream stimulatory factor (usf) proteins induce human tgf-beta1 gene activation via the glucose-response element-1013/-1002 in mesangial cells: Up-regulation of usf activity by the hexosamine biosynthetic pathway. J. Biol. Chem. 2004, 279, 15908–15915. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Skorczewski, J.; Feng, X.; Mei, L.; Murphy-Ullrich, J.E. Glucose up-regulates thrombospondin 1 gene transcription and transforming growth factor-beta activity through antagonism of cgmp-dependent protein kinase repression via upstream stimulatory factor 2. J. Biol. Chem. 2004, 279, 34311–34322. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Liu, S.; Nikolic, D.; Wang, S. High glucose levels upregulate upstream stimulatory factor 2 gene transcription in mesangial cells. J. Cell. Biochem. 2008, 103, 1952–1961. [Google Scholar] [CrossRef] [PubMed]
- Sawadogo, M. Multiple forms of the human gene-specific transcription factor usf. Ii. DNA binding properties and transcriptional activity of the purified hela usf. J. Biol. Chem. 1988, 263, 11994–12001. [Google Scholar] [PubMed]
- Ferre-D’Amare, A.R.; Pognonec, P.; Roeder, R.G.; Burley, S.K. Structure and function of the b/hlh/z domain of usf. EMBO J. 1994, 13, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; Kaestner, K.H. The foxa family of transcription factors in development and metabolism. Cell. Mol. Life Sci. 2006, 63, 2317–2328. [Google Scholar] [CrossRef]
- Kaestner, K.H.; Hiemisch, H.; Luckow, B.; Schutz, G. The hnf-3 gene family of transcription factors in mice: Gene structure, cdna sequence and mrna distribution. Genomics 1994, 20, 377–385. [Google Scholar] [CrossRef]
- Wolfrum, C.; Asilmaz, E.; Luca, E.; Friedman, J.M.; Stoffel, M. Foxa2 regulates lipid metabolism and ketogenesis in the liver during fasting and in diabetes. Nature 2004, 432, 1027–1032. [Google Scholar] [CrossRef]
- Wolfrum, C.; Shih, D.Q.; Kuwajima, S.; Norris, A.W.; Kahn, C.R.; Stoffel, M. Role of foxa-2 in adipocyte metabolism and differentiation. J. Clin. Investig. 2003, 112, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Rubins, N.E.; Ahima, R.S.; Greenbaum, L.E.; Kaestner, K.H. Foxa2 integrates the transcriptional response of the hepatocyte to fasting. Cell Metab. 2005, 2, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Le Lay, J.; Kaestner, K.H. The fox genes in the liver: From organogenesis to functional integration. Physiol. Rev. 2010, 90, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, H.J.; Kim, K.T.; Park, Y.Y.; Seong, H.A.; Park, K.C.; Lee, I.K.; Ha, H.; Shong, M.; Park, S.C.; et al. Orphan nuclear receptor small heterodimer partner represses hepatocyte nuclear factor 3/foxa transactivation via inhibition of its DNA binding. Mol. Endocrinol. 2004, 18, 2880–2894. [Google Scholar] [CrossRef]
- Yalley, A.; Schill, D.; Hatta, M.; Johnson, N.; Cirillo, L.A. Loss of interdependent binding by the foxo1 and foxa1/a2 forkhead transcription factors culminates in perturbation of active chromatin marks and binding of transcriptional regulators at insulin-sensitive genes. J. Biol. Chem. 2016, 291, 8848–8861. [Google Scholar] [CrossRef]
- Bryzgalov, L.O.; Ershov, N.I.; Ilnitskaya, S.I. Foxa transcription factors determine the amplitude of glucocorticoid induction of tyrosine aminotransferase in mice. Bull. Exp. Biol. Med. 2007, 144, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Huizing, M.; Ruitenbeek, W.; van den Heuvel, L.P.; Dolce, V.; Iacobazzi, V.; Smeitink, J.A.; Palmieri, F.; Trijbels, J.M. Human mitochondrial transmembrane metabolite carriers: Tissue distribution and its implication for mitochondrial disorders. J. Bioenerg. Biomembr. 1998, 30, 277–284. [Google Scholar] [CrossRef]
- Iacobazzi, V.; Convertini, P.; Infantino, V.; Scarcia, P.; Todisco, S.; Palmieri, F. Statins, fibrates and retinoic acid upregulate mitochondrial acylcarnitine carrier gene expression. Biochem. Biophys. Res. Commun. 2009, 388, 643–647. [Google Scholar] [CrossRef]
- Liu, M.; Lee, D.F.; Chen, C.T.; Yen, C.J.; Li, L.Y.; Lee, H.J.; Chang, C.J.; Chang, W.C.; Hsu, J.M.; Kuo, H.P.; et al. Ikkalpha activation of notch links tumorigenesis via foxa2 suppression. Mol. Cell 2012, 45, 171–184. [Google Scholar] [CrossRef]
- Vorvis, C.; Hatziapostolou, M.; Mahurkar-Joshi, S.; Koutsioumpa, M.; Williams, J.; Donahue, T.R.; Poultsides, G.A.; Eibl, G.; Iliopoulos, D. Transcriptomic and crispr/cas9 technologies reveal foxa2 as a tumor suppressor gene in pancreatic cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G1124–G1137. [Google Scholar] [CrossRef]
- Chen, X.; Cheung, S.T.; So, S.; Fan, S.T.; Barry, C.; Higgins, J.; Lai, K.M.; Ji, J.; Dudoit, S.; Ng, I.O.; et al. Gene expression patterns in human liver cancers. Mol. Biol. Cell 2002, 13, 1929–1939. [Google Scholar] [CrossRef] [PubMed]
- Mas, V.R.; Maluf, D.G.; Archer, K.J.; Yanek, K.; Kong, X.; Kulik, L.; Freise, C.E.; Olthoff, K.M.; Ghobrial, R.M.; McIver, P.; et al. Genes involved in viral carcinogenesis and tumor initiation in hepatitis C virus-induced hepatocellular carcinoma. Mol. Med. 2009, 15, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Roessler, S.; Jia, H.L.; Budhu, A.; Forgues, M.; Ye, Q.H.; Lee, J.S.; Thorgeirsson, S.S.; Sun, Z.; Tang, Z.Y.; Qin, L.X.; et al. A unique metastasis gene signature enables prediction of tumor relapse in early-stage hepatocellular carcinoma patients. Cancer Res. 2010, 70, 10202–10212. [Google Scholar] [CrossRef] [PubMed]
- Hanse, E.A.; Ruan, C.; Kachman, M.; Wang, D.; Lowman, X.H.; Kelekar, A. Cytosolic malate dehydrogenase activity helps support glycolysis in actively proliferating cells and cancer. Oncogene 2017, 36, 3915–3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infantino, V.; Iacobazzi, V.; De Santis, F.; Mastrapasqua, M.; Palmieri, F. Transcription of the mitochondrial citrate carrier gene: Role of srebp-1, upregulation by insulin and downregulation by pufa. Biochem. Biophys. Res. Commun. 2007, 356, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Santarsiero, A.; Leccese, P.; Convertini, P.; Padula, A.; Abriola, P.; D’Angelo, S.; Bisaccia, F.; Infantino, V. New insights into behcet’s syndrome metabolic reprogramming: Citrate pathway dysregulation. Mediators Inflamm. 2018, 2018, 1419352. [Google Scholar] [CrossRef]
- Infantino, V.; Iacobazzi, V.; Palmieri, F.; Menga, A. Atp-citrate lyase is essential for macrophage inflammatory response. Biochem. Biophys. Res. Commun. 2013, 440, 105–111. [Google Scholar] [CrossRef]
- Convertini, P.; Tramutola, F.; Iacobazzi, V.; Lupattelli, P.; Chiummiento, L.; Infantino, V. Permethylated anigopreissin a inhibits human hepatoma cell proliferation by mitochondria-induced apoptosis. Chem. Biol. Interact. 2015, 237, 1–8. [Google Scholar] [CrossRef]
- Iacobazzi, V.; Infantino, V.; Palmieri, F. Epigenetic mechanisms and sp1 regulate mitochondrial citrate carrier gene expression. Biochem. Biophys. Res. Commun. 2008, 376, 15–20. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| wtFOXA | 5′-TGCTTGTTTATTTATTTTAGTAGG-3′ |
| mutFOXA | 5′-TGCTTGGGGAGGGATGGTAGTAGG-3′ |
| wtUSF | 5′-GCGGGGTCACGTGTCCCTGT-3′ |
| mutUSF | 5′-GCGGGGTACCTGTTCCCTGT-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Convertini, P.; Todisco, S.; De Santis, F.; Pappalardo, I.; Iacobazzi, D.; Castiglione Morelli, M.A.; Fondufe-Mittendorf, Y.N.; Martelli, G.; Palmieri, F.; Infantino, V. Transcriptional Regulation Factors of the Human Mitochondrial Aspartate/Glutamate Carrier Gene, Isoform 2 (SLC25A13): USF1 as Basal Factor and FOXA2 as Activator in Liver Cells. Int. J. Mol. Sci. 2019, 20, 1888. https://doi.org/10.3390/ijms20081888
Convertini P, Todisco S, De Santis F, Pappalardo I, Iacobazzi D, Castiglione Morelli MA, Fondufe-Mittendorf YN, Martelli G, Palmieri F, Infantino V. Transcriptional Regulation Factors of the Human Mitochondrial Aspartate/Glutamate Carrier Gene, Isoform 2 (SLC25A13): USF1 as Basal Factor and FOXA2 as Activator in Liver Cells. International Journal of Molecular Sciences. 2019; 20(8):1888. https://doi.org/10.3390/ijms20081888
Chicago/Turabian StyleConvertini, Paolo, Simona Todisco, Francesco De Santis, Ilaria Pappalardo, Dominga Iacobazzi, Maria Antonietta Castiglione Morelli, Yvonne N. Fondufe-Mittendorf, Giuseppe Martelli, Ferdinando Palmieri, and Vittoria Infantino. 2019. "Transcriptional Regulation Factors of the Human Mitochondrial Aspartate/Glutamate Carrier Gene, Isoform 2 (SLC25A13): USF1 as Basal Factor and FOXA2 as Activator in Liver Cells" International Journal of Molecular Sciences 20, no. 8: 1888. https://doi.org/10.3390/ijms20081888