Uremic Toxin Lanthionine Interferes with the Transsulfuration Pathway, Angiogenetic Signaling and Increases Intracellular Calcium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
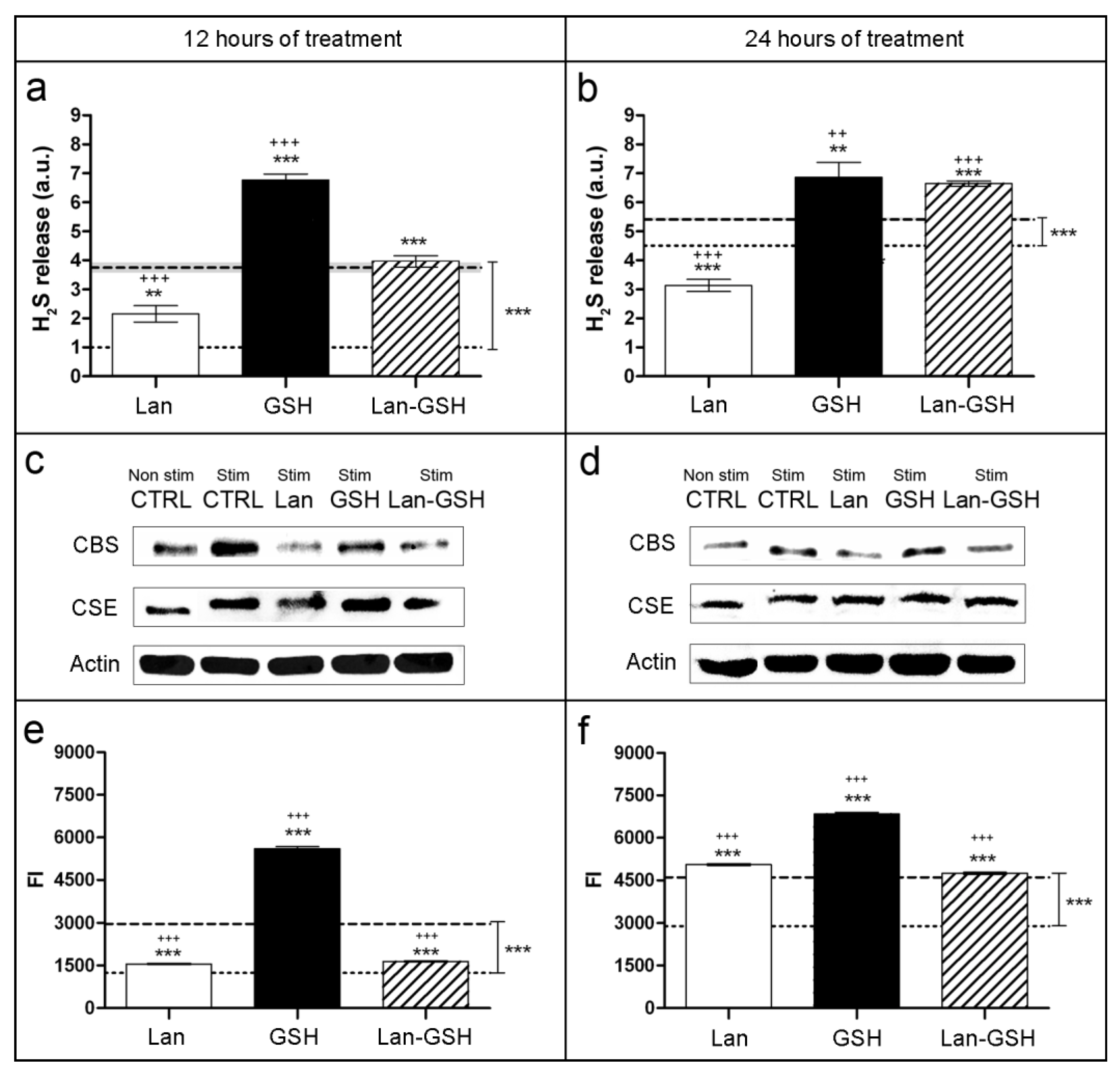
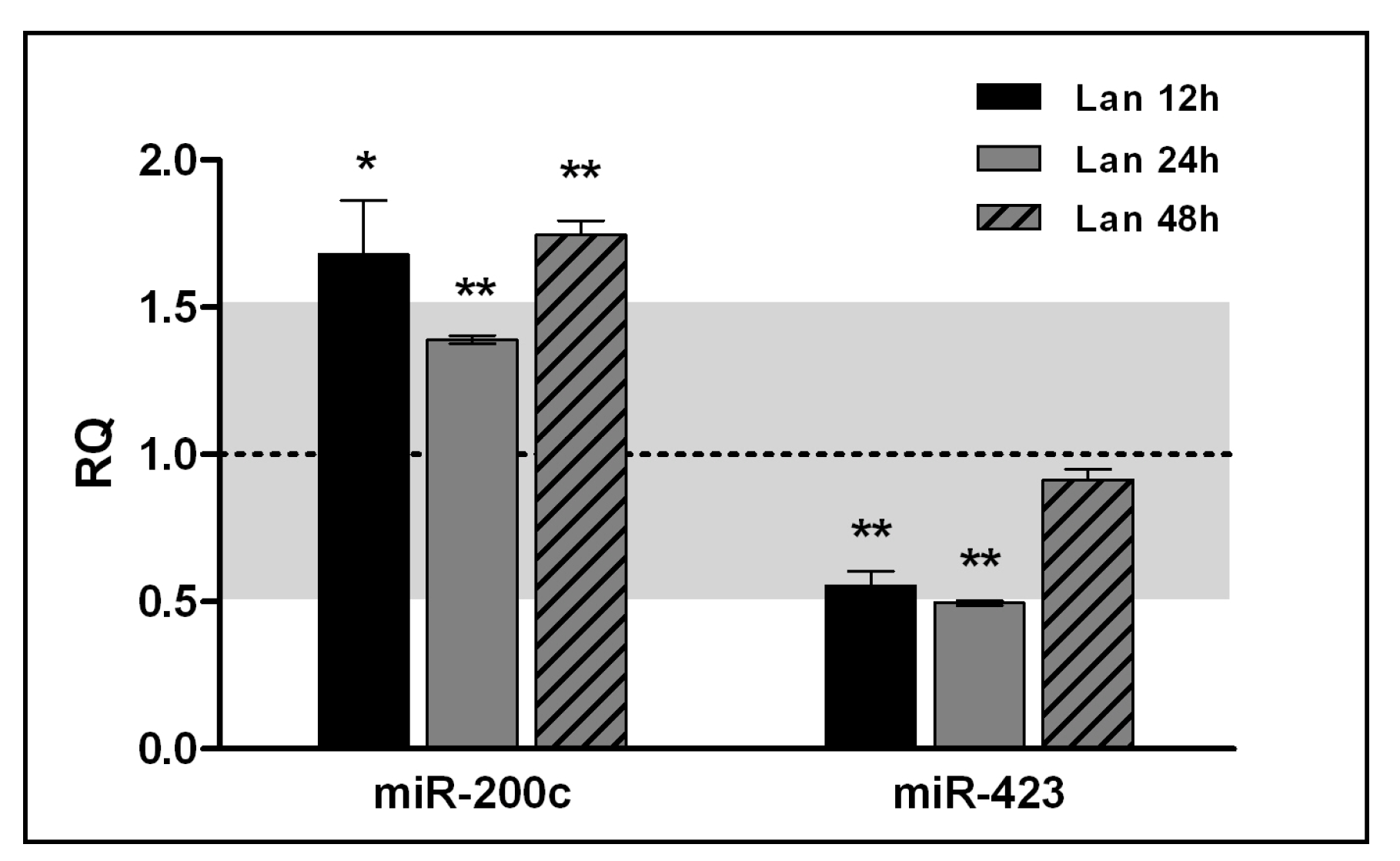
2.1. Lanthionine Decreases H2S Release and Sulfane Sulfur Cell Content as well as CBS Protein Levels
2.2. Lanthionine Hampers CBS Glutathionylation
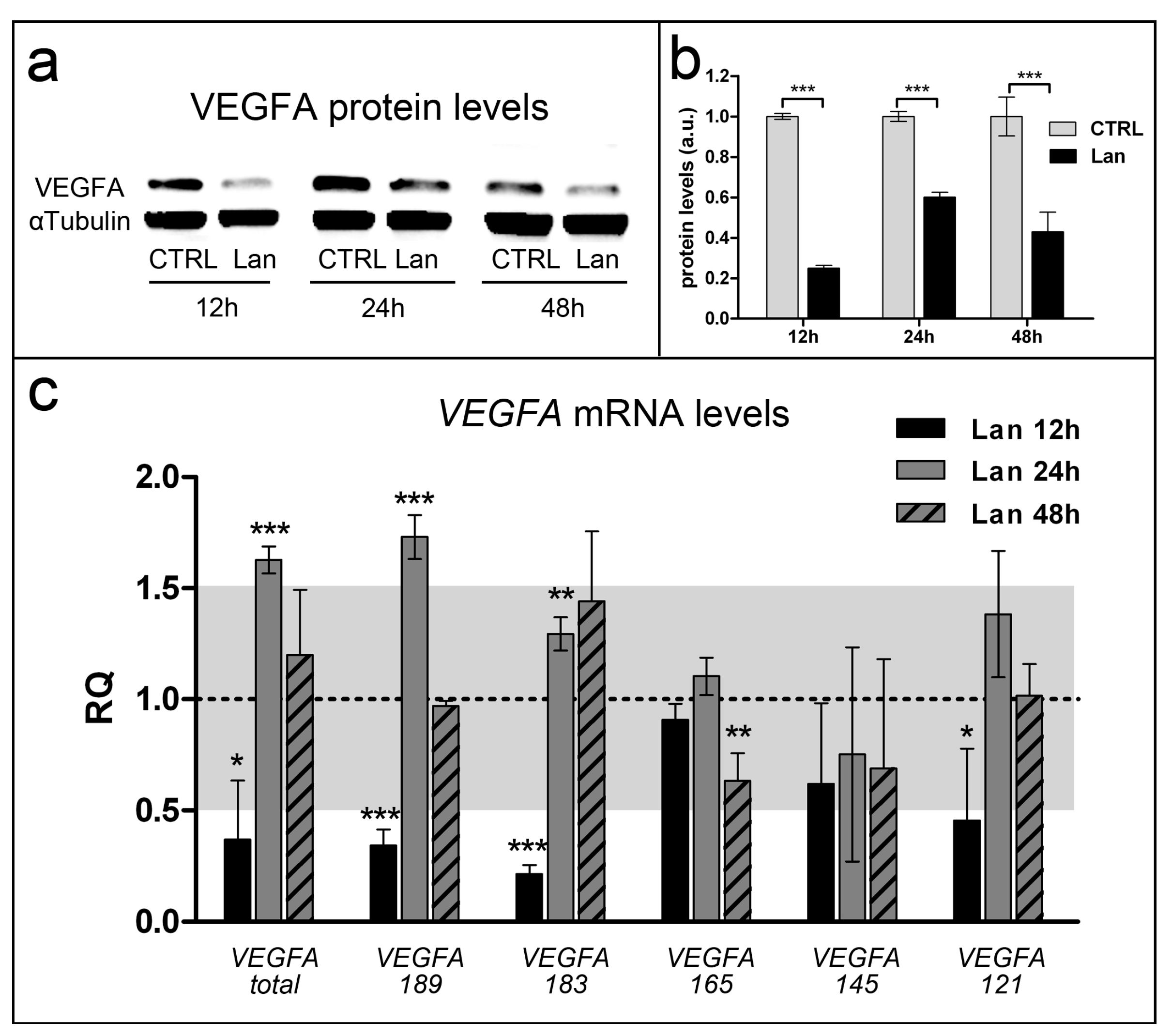
2.3. Lanthionine is Involved in Fundamental Angiogenic Pathways
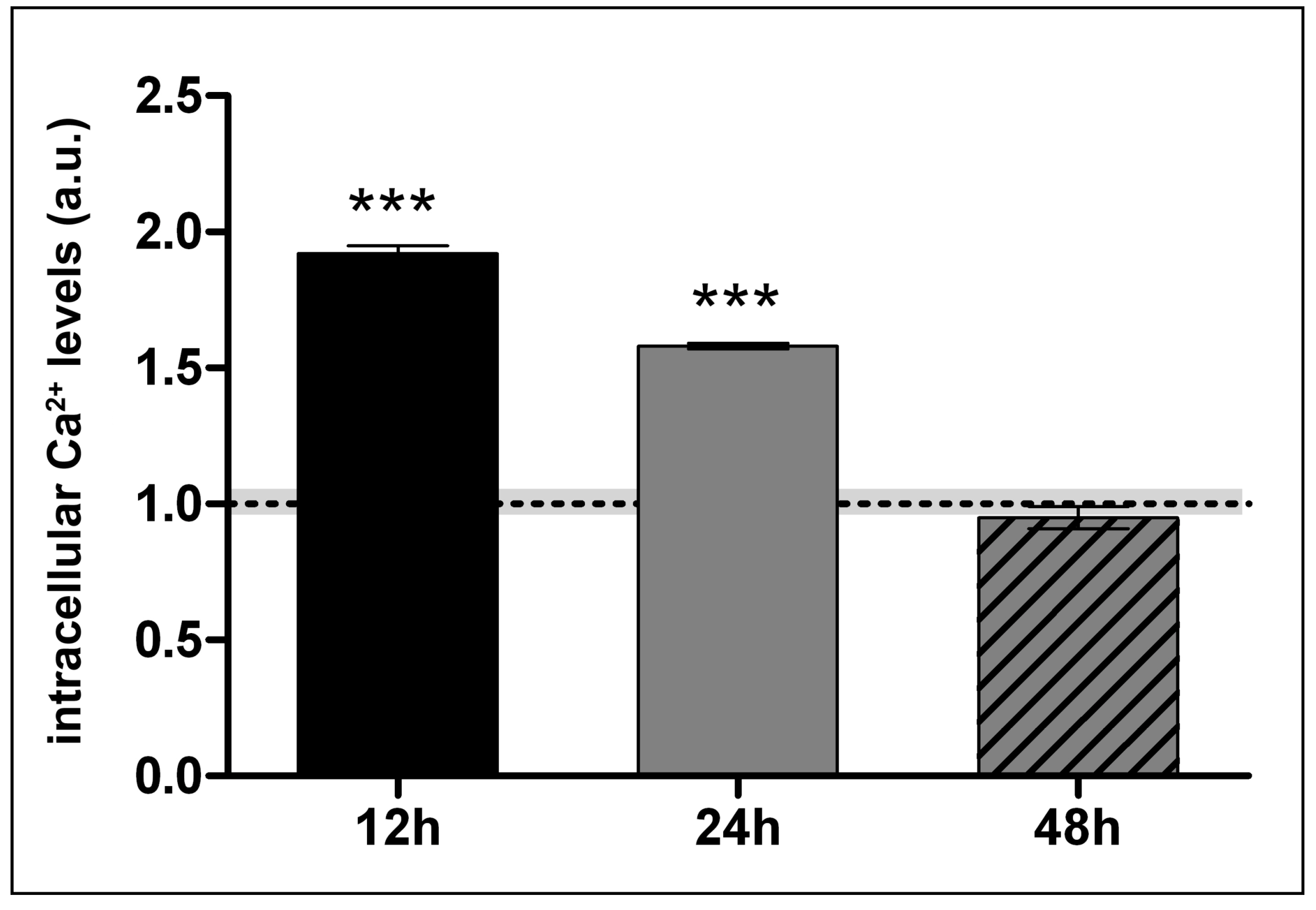
2.4. Lanthionine Increases Intracellular Calcium Levels
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. H2S Detection and Quantitation
4.3. Sulfane Sulfurs Detection
4.4. Measurement of the Intracellular Ca2+ Concentration Ratio
4.5. Protein Extraction
4.6. Immunoprecipitation
4.7. Western Blotting Analysis
4.8. RNA Extraction
4.9. Reverse Transcription, PCR and qPCR
4.10. miRNA Profiling
4.11. Bioinformatics Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AdoMet | S-adenosyl-L-methionine |
| B6 | Pyridoxalphosphate |
| Ca2+ | Calcium |
| CBS | Cystathionine-β-synthase |
| CSE | Cystathionine-γ-lyase |
| CTRL | Control |
| Cys | Cysteine |
| GSH | Glutathione |
| H2S | Hydrogen sulfide |
| Lan | Lanthionine |
| LAST | Lead Acetate Strip Test |
| qPCR | quantitative Polimerase Chain Reaction |
| SSP4 | Sulfane Sulfur Probe 4 |
| VEGF | Vascular Endothelial Growth Factor |
References
- Kimura, H. Hydrogen sulfide and polysulfides as biological mediators. Molecules 2014, 19, 16146–16157. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen sulfide and polysulfides as signaling molecules. Nitric Oxide 2015, 47, S6. [Google Scholar] [CrossRef] [Green Version]
- Chiku, T.; Padovani, D.; Zhu, W.; Singh, S.; Vitvitsky, V.; Banerjee, R. H2S Biogenesis by Human Cystathionine γ-Lyase Leads to the Novel Sulfur Metabolites Lanthionine and Homolanthionine and Is Responsive to the Grade of Hyperhomocysteinemia. J. Biol. Chem. 2009, 284, 11601–11612. [Google Scholar] [CrossRef] [PubMed]
- Majtan, T.; Krijt, J.; Sokolová, J.; Křížková, M.; Ralat, M.A.; Kent, J.; Gregory, J.F., III; Kožich, V.; Kraus, J.P. Biogenesis of Hydrogen Sulfide and Thioethers by Cystathionine Beta-Synthase. Antioxid. Redox Signal. 2018, 28, 311–323. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Enzymology of H2S biogenesis, decay and signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Perna, A.F.; Di Nunzio, A.; Amoresano, A.; Pane, F.; Fontanarosa, C.; Pucci, P.; Vigorito, C.; Cirillo, G.; Zacchia, M.; Trepiccione, F.; et al. Divergent behavior of hydrogen sulfide pools and of the sulfur metabolite lanthionine, a novel uremic toxin, in dialysis patients. Biochimie 2016, 126, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Perna, A.F.; Anishchenko, E.; Vigorito, C.; Zacchia, M.; Trepiccione, F.; D’Aniello, S.; Ingrosso, D. Zebrafish, a Novel Model System to Study Uremic Toxins: The Case for the Sulfur Amino Acid Lanthionine. Int. J. Mol. Sci. 2018, 19, 1323. [Google Scholar] [CrossRef] [PubMed]
- Bibli, S.I.; Luck, B.; Zukunft, S.; Wittig, J.; Chen, W.; Xian, M.; Papapetropoulos, A.; Hu, J.; Fleming, I. A selective and sensitive method for quantification of endogenous polysulfide production in biological samples. Redox Biol. 2018, 18, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S signals through protein S-sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [PubMed]
- Vitvitsky, V.; Yadav, P.K.; An, S.; Seravalli, J.; Cho, U.S.; Banerjee, R. Structural and Mechanistic Insights into Hemoglobin-catalyzed Hydrogen Sulfide Oxidation and the Fate of Polysulfide Products. J. Biol. Chem. 2017, 292, 5584–5592. [Google Scholar] [CrossRef] [PubMed]
- Anishchenko, E.; Vigorito, C.; Mele, L.; Lombari, P.; Perna, A.; Ingrosso, D. Novel Applications of Lead Acetate and Flow Cytometry Methods for Detection of Sulfur-Containing Molecules. Methods Protoc. 2019, 2, 13. [Google Scholar] [CrossRef]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [Green Version]
- Niu, W.N.; Yadav, P.K.; Adamec, J.; Banerjee, R. S-glutathionylation enhances human cystathionine β-synthase activity under oxidative stress conditions. Antioxid. Redox Signal. 2015, 22, 350–361. [Google Scholar] [CrossRef]
- Pavkovic, M.; Robinson-Cohen, C.; Chua, A.S.; Nicoara, O.; Cárdenas-González, M.; Bijol, V.; Ramachandran, K.; Hampson, L.; Pirmohamed, M.; Antoine, D.J.; et al. Detection of Drug-Induced Acute Kidney Injury in Humans Using Urinary KIM-1, miR-21, -200c, and -423. Toxicol. Sci. 2016, 152, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, K.; Saikumar, J.; Bijol, V.; Koyner, J.L.; Qian, J.; Betensky, R.A.; Waikar, S.S.; Vaidya, V.S. Human miRNome profiling identifies microRNAs differentially present in the urine after kidney injury. Clin. Chem. 2013, 59, 1742–1752. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.H.; Wang, H.; Chen, L.; Klein, J.D.; Sands, J.M. Urea Transporter B and MicroRNA-200c Differ in Kidney Outer Versus Inner Medulla Following Dehydration. Am. J. Med. Sci. 2016, 352, 296–301. [Google Scholar] [CrossRef]
- Szeto, C.-C. Urine miRNA in nephrotic syndrome. Clin. Chim. Acta 2014, 436, 308–313. [Google Scholar] [CrossRef]
- Lv, L.L.; Cao, Y.; Liu, D.; Xu, M.; Liu, H.; Tang, R.N.; Ma, K.L.; Liu, B.C. Isolation and Quantification of MicroRNAs from Urinary Exosomes/Microvesicles for Biomarker Discovery. Int. J. Biol. Sci. 2013, 9, 1021–1031. [Google Scholar] [CrossRef] [Green Version]
- Sataranatarajan, K.; Feliers, D.; Mariappan, M.M.; Lee, H.J.; Lee, M.J.; Day, R.T.; Yalamanchili, H.B.; Choudhury, G.G.; Barnes, J.L.; Van Remmen, H.; et al. Molecular events in matrix protein metabolism in the aging kidney. Aging Cell 2012, 11, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.P.; Liu, L.S.; Chen, C.B.; Zhou, J.; Zheng, Y.T.; Wang, X.P.; Han, M.; Wang, C.X. MicroRNA-423-5p facilitates hypoxia/reoxygenation-induced apoptosis in renal proximal tubular epithelial cells by targeting GSTM1 via endoplasmic reticulum stress. Oncotarget 2017, 8, 82064–82077. [Google Scholar] [CrossRef] [Green Version]
- Koo, T.; Cho, B.J.; Kim, D.H.; Park, J.M.; Choi, E.J.; Kim, H.H.; Lee, D.J.; Kim, I.A. MicroRNA-200c increases radiosensitivity of human cancer cells with activated EGFR-associated signaling. Oncotarget 2017, 8, 65457–65468. [Google Scholar] [CrossRef] [Green Version]
- Zygalaki, E.; Kaklamanis, L.; Nikolaou, N.I.; Kyrzopoulos, S.; Houri, M.; Kyriakides, Z.; Lianidou, E.S.; Kremastinos, D.T. Expression profile of total VEGF, VEGF splice variants and VEGF receptors in the myocardium and arterial vasculature of diabetic and non-diabetic patients with coronary artery disease. Clin. Biochem. 2008, 41, 82–87. [Google Scholar] [CrossRef]
- Potenza, D.M.; Guerra, G.; Avanzato, D.; Poletto, V.; Pareek, S.; Guido, D.; Gallanti, A.; Rosti, V.; Munaron, L.; Tanzi, F.; et al. Hydrogen sulphide triggers VEGF-induced intracellular Ca2 signals in human endothelial cells but not in their immature progenitors. Cell Calcium 2014, 56, 225–234. [Google Scholar] [CrossRef]
- Perna, A.F.; Glorieux, G.; Zacchia, M.; Trepiccione, F.; Capolongo, G.; Vigorito, C.; Anishchenko, E.; Ingrosso, D. The role of the intestinal microbiota in uremic solute accumulation: a focus on sulfur compounds. J. Nephrol. 2019. [Google Scholar] [CrossRef]
- Perna, A.F.; Ingrosso, D.; Satta, E.; Lombardi, C.; Acanfora, F.; De Santo, N.G. Homocysteine metabolism in renal failure. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 53–57. [Google Scholar] [CrossRef]
- Perna, A.F. Plasma Protein Aspartyl Damage Is Increased in Hemodialysis Patients: Studies on Causes and Consequences. J. Am. Soc. Nephrol. 2004, 15, 2747–2754. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N. Vascular Endothelial Growth Factor: Basic Science and Clinical Progress. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef] [Green Version]
- Shih, S.C.; Ju, M.; Liu, N.; Mo, J.R.; Ney, J.J.; Smith, L.E.H. Transforming growth factor beta1 induction of vascular endothelial growth factor receptor 1: mechanism of pericyte-induced vascular survival in vivo. Proc. Natl. Acad. Sci. USA 2003, 100, 15859–15864. [Google Scholar] [CrossRef]
- Frangié, C.; Lefaucheur, C.; Medioni, J.; Jacquot, C.; Hill, G.S.; Nochy, D. Renal thrombotic microangiopathy caused by anti-VEGF-antibody treatment for metastatic renal-cell carcinoma. Lancet Oncol. 2007, 8, 177–178. [Google Scholar] [CrossRef]
- Eremina, V.; Cui, S.; Gerber, H.; Ferrara, N.; Haigh, J.; Nagy, A.; Ema, M.; Rossant, J.; Jothy, S.; Miner, J.H.; et al. Vascular endothelial growth factor a signaling in the podocyte-endothelial compartment is required for mesangial cell migration and survival. J. Am. Soc. Nephrol. 2006, 17, 724–735. [Google Scholar] [CrossRef]
- Jingjing, L.; Srinivasan, B.; Roque, R.S. Ectodomain shedding of VEGF183, a novel isoform of vascular endothelial growth factor, promotes its mitogenic activity in vitro. Angiogenesis 2001, 4, 103–112. [Google Scholar] [CrossRef]
- Houck, K.A.; Ferrara, N.; Winer, J.; Cachianes, G.; Li, B.; Leung, D.W. The vascular endothelial growth factor family: identification of a fourth molecular species and characterization of alternative splicing of RNA. Mol. Endocrinol. 1991, 5, 1806–1814. [Google Scholar] [CrossRef]
- Wang, X.-F.; Cui, J.Z.; Prasad, S.S.; Matsubara, J.A. Altered gene expression of angiogenic factors induced by calcium-mediated dissociation of retinal pigment epithelial cells. Invest. Ophthalmol. Vis. Sci. 2005, 46, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Medford, A.R.L.; Douglas, S.K.; Godinho, S.I.H.; Uppington, K.M.; Armstrong, L.; Gillespie, K.M.; van Zyl, B.; Tetley, T.D.; Ibrahim, N.B.N.; Millar, A.B. Vascular Endothelial Growth Factor (VEGF) isoform expression and activity in human and murine lung injury. Respir. Res. 2009, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Azimi-Nezhad, M. Vascular endothelial growth factor from embryonic status to cardiovascular pathology. Rep. Biochem. Mol. Biol. 2014, 2, 59–69. [Google Scholar]
- Bergantino, F.; Guariniello, S.; Raucci, R.; Colonna, G.; De Luca, A.; Normanno, N.; Costantini, S. Structure–fluctuation–function relationships of seven pro-angiogenic isoforms of VEGFA, important mediators of tumorigenesis. Biochim. Biophys. Acta-Proteins Proteom. 2015, 1854, 410–425. [Google Scholar] [CrossRef] [PubMed]
- Tessler, S.; Rockwell, P.; Hicklin, D.; Cohen, T.; Levi, B.Z.; Witte, L.; Lemischka, I.R.; Neufeld, G. Heparin modulates the interaction of VEGF165 with soluble and cell associated flk-1 receptors. J. Biol. Chem. 1994, 269, 12456–12461. [Google Scholar] [PubMed]
- McCullough, P.A.; Agarwal, M.; Agrawal, V. Review article: Risks of coronary artery calcification in chronic kidney disease: do the same rules apply? Nephrology 2009, 14, 428–436. [Google Scholar] [CrossRef]
- Coletta, C.; Papapetropoulos, A.; Erdelyi, K.; Olah, G.; Modis, K.; Panopoulos, P.; Asimakopoulou, A.; Gero, D.; Szabo, C. P23 The angiogenic and vasorelaxant effect of H2S require the endogenous production of NO, while the angiogenic and vasorelaxant effect of NO require the endogenous production of H2S. Nitric Oxide 2012, 27, S22–S23. [Google Scholar] [CrossRef]
- Vicente, J.B.; Malagrinò, F.; Arese, M.; Forte, E.; Sarti, P.; Giuffrè, A. Bioenergetic relevance of hydrogen sulfide and the interplay between gasotransmitters at human cystathionine β-synthase. Biochim. Biophys. Acta 2016, 1857, 1127–1138. [Google Scholar] [CrossRef]
- Vicente, J.B.; Colaço, H.G.; Malagrinò, F.; Arese, M.; Forte, E.; Sarti, P.; Leandro, P.; Giuffrè, A. S-adenosyl-L-methionine and the Cross-talk between the Bioenergetically Relevant Gasotransmitters H 2 S, NO and CO at Human Cystathionine β-synthase. Biochim. Biophys. Acta-Bioenergetics 2016, 1857, e81–e82. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vigorito, C.; Anishchenko, E.; Mele, L.; Capolongo, G.; Trepiccione, F.; Zacchia, M.; Lombari, P.; Capasso, R.; Ingrosso, D.; Perna, A.F. Uremic Toxin Lanthionine Interferes with the Transsulfuration Pathway, Angiogenetic Signaling and Increases Intracellular Calcium. Int. J. Mol. Sci. 2019, 20, 2269. https://doi.org/10.3390/ijms20092269
Vigorito C, Anishchenko E, Mele L, Capolongo G, Trepiccione F, Zacchia M, Lombari P, Capasso R, Ingrosso D, Perna AF. Uremic Toxin Lanthionine Interferes with the Transsulfuration Pathway, Angiogenetic Signaling and Increases Intracellular Calcium. International Journal of Molecular Sciences. 2019; 20(9):2269. https://doi.org/10.3390/ijms20092269
Chicago/Turabian StyleVigorito, Carmela, Evgeniya Anishchenko, Luigi Mele, Giovanna Capolongo, Francesco Trepiccione, Miriam Zacchia, Patrizia Lombari, Rosanna Capasso, Diego Ingrosso, and Alessandra F. Perna. 2019. "Uremic Toxin Lanthionine Interferes with the Transsulfuration Pathway, Angiogenetic Signaling and Increases Intracellular Calcium" International Journal of Molecular Sciences 20, no. 9: 2269. https://doi.org/10.3390/ijms20092269