miR-129-5p and miR-130a-3p Regulate VEGFR-2 Expression in Sensory and Motor Neurons during Development

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. VEGFR-2 Expression in Immature and Mature Sensory Neurons in Drgs
2.2. VEGFR-2 Expression in Immature and Mature Motor Neurons of the Spinal Cord
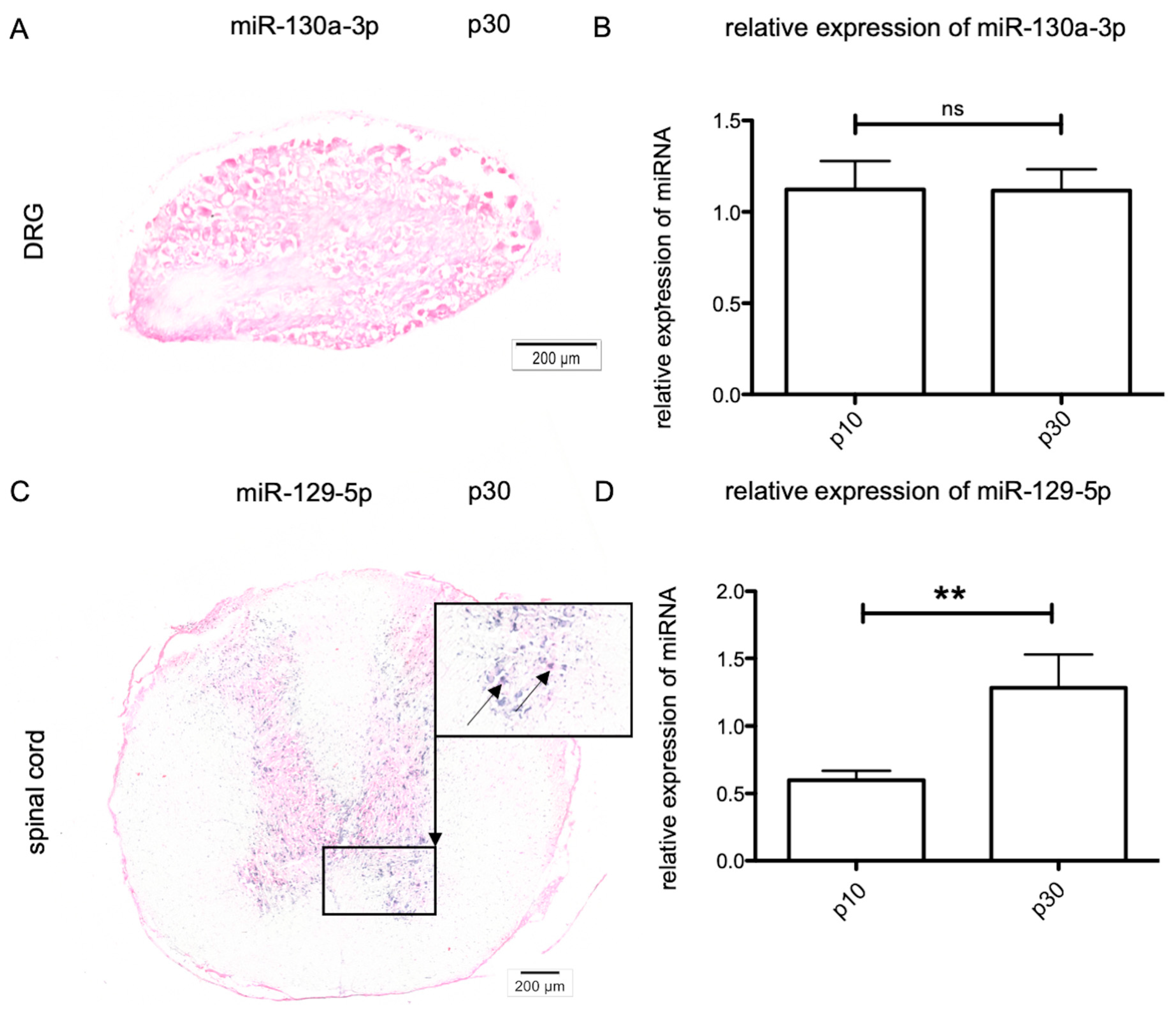
2.3. Age-Dependent MiRNA Expression in Sensory Neurons
2.4. Age-Dependent miRNA Expression in Lower Motor Neurons
2.5. Impact of Mir-130a-3p in Sensory Neurons
2.6. Impact of Mir-129a-5p in Lower Motor Neurons
3. Discussion
3.1. Age-Dependent VEGFR-2 Expression in Sensory DRG Neurons and Spinal Cord Motor Neurons
3.2. miRNA Expression in DRG Neurons and Spinal Cord Motor Neurons
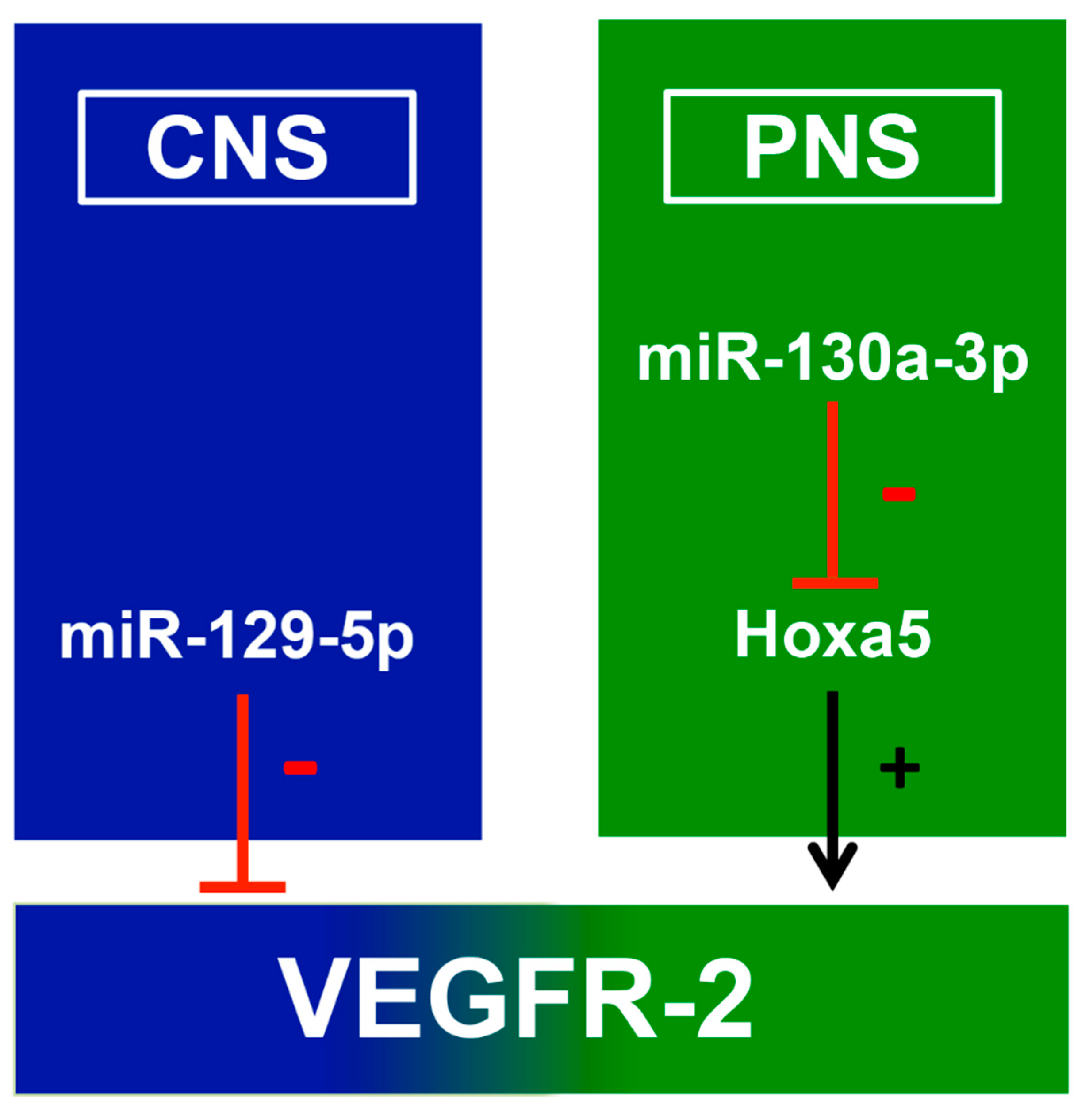
3.3. The Influence of miRNA on Kdr Expression
3.4. Mir-129-5p and Mir-130a-3p could Be Therapeutic Targets in Neurological Diseases
4. Material and Methods
4.1. Animals and Surgical Procedures
4.2. Immunohistochemistry
4.3. Western Blot
4.4. Cryosections for in Situ Hybridization
4.5. In Situ Hybridization
4.6. mRNA- and Total RNA Extraction, Reverse Transcription, and RT-Qpcr
4.7. Cell Culture
4.8. Transfection
4.9. Statistical Analysis of RT-qPCR
5. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Senger, D.R.; Galli, S.J.; Dvorak, A.M.; Perruzzi, C.A.; Harvey, V.S.; Dvorak, H.F. Tumor cells secrete a vascular permeability factor that promotes accumulation of ascites fluid. Science 1983, 219, 983–985. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Ruhrberg, C.; Bautch, V.L. Neurovascular development and links to disease. Cell. Mol. Life Sci. 2013, 70, 1675–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gora-Kupilas, K.; Josko, J. The neuroprotective function of vascular endothelial growth factor (VEGF). Folia Neuropathol. 2005, 43, 31–39. [Google Scholar]
- Foehring, D.; Brand-Saberi, B.; Theiss, C. VEGF-induced growth cone enhancement is diminished by inhibiting tyrosine-residue 1214 of VEGFR-2. Cells Tissues Organs 2012, 196, 195–205. [Google Scholar] [CrossRef]
- Roy, H.; Bhardwaj, S.; Yla-Herttuala, S. Biology of vascular endothelial growth factors. FEBS Lett. 2006, 580, 2879–2887. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.; Dunk, C.; Ahmad, S.; Khaliq, A. Regulation of placental vascular endothelial growth factor (VEGF) and placenta growth factor (PIGF) and soluble Flt-1 by oxygen—A review. Placenta 2000, 21 (Suppl. A), S16–S24. [Google Scholar] [CrossRef]
- Ruch, C.; Skiniotis, G.; Steinmetz, M.O.; Walz, T.; Ballmer-Hofer, K. Structure of a VEGF-VEGF receptor complex determined by electron microscopy. Nat. Struct. Mol. Biol. 2007, 14, 249–250. [Google Scholar] [CrossRef]
- Selvaraj, S.K.; Giri, R.K.; Perelman, N.; Johnson, C.; Malik, P.; Kalra, V.K. Mechanism of monocyte activation and expression of proinflammatory cytochemokines by placenta growth factor. Blood 2003, 102, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Hattori, K.; Heissig, B.; Wu, Y.; Dias, S.; Tejada, R.; Ferris, B.; Hicklin, D.J.; Zhu, Z.; Bohlen, P.; Witte, L.; et al. Placental growth factor reconstitutes hematopoiesis by recruiting VEGFR1(+) stem cells from bone-marrow microenvironment. Nat. Med. 2002, 8, 841–849. [Google Scholar] [CrossRef]
- Dhondt, J.; Peeraer, E.; Verheyen, A.; Nuydens, R.; Buysschaert, I.; Poesen, K.; Van Geyte, K.; Beerens, M.; Shibuya, M.; Haigh, J.J.; et al. Neuronal FLT1 receptor and its selective ligand VEGF-B protect against retrograde degeneration of sensory neurons. FASEB J. 2011, 25, 1461–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, J.; Huang, X.; Li, Y. The methods and mechanisms to differentiate endothelial-like cells and smooth muscle cells from mesenchymal stem cells for vascularization in vaginal reconstruction. Mol. Biotechnol. 2018, 60, 396–411. [Google Scholar] [CrossRef]
- Castaneda-Cabral, J.L.; Beas-Zarate, C.; Gudino-Cabrera, G.; Urena-Guerrero, M.E. Glutamate neonatal excitotoxicity modifies VEGF-A, VEGF-B, VEGFR-1 and VEGFR-2 protein expression profiles during postnatal development of the cerebral cortex and hippocampus of male rats. J. Mol. Neurosci. 2017, 63, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Veikkola, T.; Jussila, L.; Makinen, T.; Karpanen, T.; Jeltsch, M.; Petrova, T.V.; Kubo, H.; Thurston, G.; McDonald, D.M.; Achen, M.G.; et al. Signalling via vascular endothelial growth factor receptor-3 is sufficient for lymphangiogenesis in transgenic mice. EMBO J. 2001, 20, 1223–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, M.C.; Cunningham, A.M. Developmental expression of vascular endothelial growth factor receptor 3 and vascular endothelial growth factor C in forebrain. Neuroscience 2015, 303, 544–557. [Google Scholar] [CrossRef] [PubMed]
- Zachary, I. Neuroprotective role of vascular endothelial growth factor: Signalling mechanisms, biological function, and therapeutic potential. Neurosignals 2005, 14, 207–221. [Google Scholar] [CrossRef]
- Herrfurth, L.; Theis, V.; Matschke, V.; May, C.; Marcus, K.; Theiss, C. Morphological plasticity of emerging purkinje cells in response to exogenous VEGF. Front. Mol. Neurosci. 2017, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Olbrich, L.; Foehring, D.; Happel, P.; Brand-Saberi, B.; Theiss, C. Fast rearrangement of the neuronal growth cone’s actin cytoskeleton following VEGF stimulation. Histochem. Cell Biol. 2013, 139, 431–445. [Google Scholar] [CrossRef]
- Dumpich, M.; Mannherz, H.G.; Theiss, C. VEGF signaling regulates cofilin and the Arp2/3-complex within the axonal growth cone. Curr. Neurovasc. Res. 2015, 12, 293–307. [Google Scholar] [CrossRef]
- Theis, V.; Theiss, C. VEGF—A stimulus for neuronal development and regeneration in the CNS and PNS. Curr. Protein Pept. Sci. 2018, 19, 589–597. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xia, W.; Yang, Y.; Mason, A.S.; Lei, X.; Ma, Z. Characterization and evolution of conserved MicroRNA through duplication events in date palm (Phoenix dactylifera). PLoS ONE 2013, 8, e71435. [Google Scholar] [CrossRef] [PubMed]
- Davis-Dusenbery, B.N.; Hata, A. Mechanisms of control of microRNA biogenesis. J. Biochem. 2010, 148, 381–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Sayed, D.; Abdellatif, M. MicroRNAs in development and disease. Physiol. Rev. 2011, 91, 827–887. [Google Scholar] [CrossRef]
- Pieczora, L.; Stracke, L.; Vorgerd, M.; Hahn, S.; Theiss, C.; Theis, V. Unveiling of miRNA expression patterns in purkinje cells during development. Cerebellum 2017, 16, 376–387. [Google Scholar] [CrossRef]
- Soufi-Zomorrod, M.; Hajifathali, A.; Kouhkan, F.; Mehdizadeh, M.; Rad, S.M.; Soleimani, M. MicroRNAs modulating angiogenesis: MiR-129-1 and miR-133 act as angio-miR in HUVECs. Tumour Biol. 2016, 37, 9527–9534. [Google Scholar] [CrossRef]
- Mujahid, S.; Nielsen, H.C.; Volpe, M.V. MiR-221 and miR-130a regulate lung airway and vascular development. PLoS ONE 2013, 8, e55911. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.L.; Li, S.Y.; Shang, F. Effect of microRNA-129-5p targeting HMGB1-RAGE signaling pathway on revascularization in a collagenase-induced intracerebral hemorrhage rat model. Biomed. Pharm. 2017, 93, 238–244. [Google Scholar] [CrossRef]
- Gerber, H.P.; Dixit, V.; Ferrara, N. Vascular endothelial growth factor induces expression of the antiapoptotic proteins Bcl-2 and A1 in vascular endothelial cells. J. Biol. Chem. 1998, 273, 13313–13316. [Google Scholar] [CrossRef] [Green Version]
- Sondell, M.; Lundborg, G.; Kanje, M. Vascular endothelial growth factor has neurotrophic activity and stimulates axonal outgrowth, enhancing cell survival and Schwann cell proliferation in the peripheral nervous system. J. Neurosci. 1999, 19, 5731–5740. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Fukuoka, S.; Karagianni, N.; Guaiquil, V.H.; Rosenblatt, M.I. Vascular endothelial growth factor promotes anatomical and functional recovery of injured peripheral nerves in the avascular cornea. FASEB J. 2013, 27, 2756–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanojlovic, M.; Pang, X.; Lin, Y.; Stone, S.; Cvetanovic, M.; Lin, W. Inhibition of vascular endothelial growth factor receptor 2 exacerbates loss of lower motor neurons and axons during experimental autoimmune encephalomyelitis. PLoS ONE 2016, 11, e0160158. [Google Scholar] [CrossRef] [PubMed]
- Sondell, M.; Kanje, M. Postnatal expression of VEGF and its receptor flk-1 in peripheral ganglia. Neuroreport 2001, 12, 105–108. [Google Scholar] [CrossRef]
- Losordo, D.W.; Isner, J.M. Estrogen and angiogenesis: A review. Arter. Thromb. Vasc. Biol. 2001, 21, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Reynders, K.; Wauters, E.; Moisse, M.; Decaluwé, H.; De Leyn, P.; Peeters, S.; Lambrecht, M.; Nackaerts, K.; Dooms, C.; Janssens, W.; et al. RNA-sequencing in non-small cell lung cancer shows gene downregulation of therapeutic targets in tumor tissue compared to non-malignant lung tissue. Radiat. Oncol. 2018, 13, 131. [Google Scholar] [CrossRef]
- Petersen, C.P.; Bordeleau, M.-E.; Pelletier, J.; Sharp, P.A. Short RNAs repress translation after initiation in mammalian cells. Mol. Cell 2006, 21, 533–542. [Google Scholar] [CrossRef]
- Hu, Z.; Li, Z. miRNAs in synapse development and synaptic plasticity. Curr. Opin. Neurobiol. 2017, 45, 24–31. [Google Scholar] [CrossRef]
- Madelaine, R.; Sloan, S.A.; Huber, N.; Notwell, J.H.; Leung, L.C.; Skariah, G.; Halluin, C.; Pasca, S.P.; Bejerano, G.; Krasnow, M.A.; et al. MicroRNA-9 couples brain neurogenesis and angiogenesis. Cell Rep. 2017, 20, 1533–1542. [Google Scholar] [CrossRef] [Green Version]
- Rinchetti, P.; Rizzuti, M.; Faravelli, I.; Corti, S. MicroRNA metabolism and dysregulation in amyotrophic lateral sclerosis. Mol. Neurobiol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Di, G.; Dong, M.; Qu, M.; Zhao, X.; Duan, H.; Hu, X.; Liu, T.; Zhou, Q.; Shi, W.; et al. Epithelium-derived miR-204 inhibits corneal neovascularization. Exp. Eye Res. 2018, 167, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Chen, F.S.; Tan, W.F.; Fang, B.; Zhang, Z.L.; Ma, H. Elevated microRNA-129-5p level ameliorates neuroinflammation and blood-spinal cord barrier damage after ischemia-reperfusion by inhibiting HMGB1 and the TLR3-cytokine pathway. J. Neuroinflammation 2017, 14, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajman, M.; Metge, F.; Fiore, R.; Khudayberdiev, S.; Aksoy-Aksel, A.; Bicker, S.; Ruedell Reschke, C.; Raoof, R.; Brennan, G.P.; Delanty, N.; et al. A microRNA-129-5p/Rbfox crosstalk coordinates homeostatic downscaling of excitatory synapses. EMBO J. 2017, 36, 1770–1787. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Huang, W.; Lu, Q.; Chen, H.; Liu, J.; Hong, B. miR24 regulates angiogenesis in gliomas. Mol. Med. Rep. 2018, 18, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Zhou, S.; Wang, D.; He, W.; Li, J.; Zhang, S. MicroRNA-205 is downregulated in hepatocellular carcinoma and inhibits cell growth and metastasis via directly targeting vascular endothelial growth factor A. Oncol. Lett. 2018, 16, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- Kontarakis, Z.; Rossi, A.; Ramas, S.; Dellinger, M.T.; Stainier, D.Y.R. Mir-126 is a conserved modulator of lymphatic development. Dev. Biol. 2018, 437, 120–130. [Google Scholar] [CrossRef]
- Lu, B.; Liu, G.L.; Yu, F.; Li, W.J.; Xiang, X.X.; Xiao, H.Z. MicroRNA16/VEGFR2/p38/NFkappaB signaling pathway regulates cell growth of human pituitary neoplasms. Oncol. Rep. 2018, 39, 1235–1244. [Google Scholar] [CrossRef] [Green Version]
- Pengcheng, S.; Ziqi, W.; Luyao, Y.; Xiangwei, Z.; Liang, L.; Yuwei, L.; Lechen, L.; Wanhai, X. MicroRNA-497 suppresses renal cell carcinoma by targeting VEGFR-2 in ACHN cells. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [Green Version]
- Salinas-Vera, Y.M.; Marchat, L.A.; Garcia-Vazquez, R.; Gonzalez de la Rosa, C.H.; Castaneda-Saucedo, E.; Tito, N.N.; Flores, C.P.; Perez-Plasencia, C.; Cruz-Colin, J.L.; Carlos-Reyes, A.; et al. Cooperative multi-targeting of signaling networks by angiomiR-204 inhibits vasculogenic mimicry in breast cancer cells. Cancer Lett. 2018, 432, 17–27. [Google Scholar] [CrossRef]
- Horst, C.H.; Titze-de-Almeida, R.; Titze-de-Almeida, S.S. The involvement of Eag1 potassium channels and miR-34a in rotenone-induced death of dopaminergic SH-SY5Y cells. Mol. Med. Rep. 2017, 15, 1479–1488. [Google Scholar] [CrossRef] [PubMed]
- Roser, A.E.; Caldi Gomes, L.; Halder, R.; Jain, G.; Maass, F.; Tonges, L.; Tatenhorst, L.; Bahr, M.; Fischer, A.; Lingor, P.; et al. miR-182-5p and miR-183-5p act as GDNF mimics in dopaminergic midbrain neurons. Mol. Ther. Nucleic Acids 2018, 11, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Liu, D.Q.; Zheng, C.; Zheng, S.Q.; Liu, M.; Li, X.; Tang, H. miR-200a modulate HUVECs viability and migration. IUBMB Life 2011, 63, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.J.; Kwok, Z.H.; Chew, X.H.; Zhang, B.; Liu, C.; Soong, T.W.; Yang, H.; Tay, Y. A FTH1 gene:pseudogene:microRNA network regulates tumorigenesis in prostate cancer. Nucleic Acids Res. 2018, 46, 1998–2011. [Google Scholar] [CrossRef] [PubMed]
- Sossey-Alaoui, K.; Bialkowska, K.; Plow, E.F. The miR200 family of microRNAs regulates WAVE3-dependent cancer cell invasion. J. Biol. Chem. 2009, 284, 33019–33029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekdash, R.A.; Harrison, N.L. Downregulation of Gabra4 expression during alcohol withdrawal is mediated by specific microRNAs in cultured mouse cortical neurons. Brain Behav. 2015, 5, e00355. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Miao, L.; Mei, Y.; Wu, M. Retinoic acid-induced HOXA5 expression is co-regulated by HuR and miR-130a. Cell. Signal. 2013, 25, 1476–1485. [Google Scholar] [CrossRef]
- Chen, Y.; Gorski, D.H. Regulation of angiogenesis through a microRNA (miR-130a) that down-regulates antiangiogenic homeobox genes GAX and HOXA5. Blood 2008, 111, 1217–1226. [Google Scholar] [CrossRef]
- Rhoads, K.; Arderiu, G.; Charboneau, A.; Hansen, S.L.; Hoffman, W.; Boudreau, N. A role for Hox A5 in regulating angiogenesis and vascular patterning. Lymphat. Res. Biol. 2005, 3, 240–252. [Google Scholar] [CrossRef]
- Silfa-Mazara, F.; Mujahid, S.; Thomas, C.; Vong, T.; Larsson, I.; Nielsen, H.C.; Volpe, M.V. Oxygen differentially affects the hox proteins Hoxb5 and Hoxa5 altering airway branching and lung vascular formation. J. Cell. Commun. Signal. 2014, 8, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Ridolfi, B.; Abdel-Haq, H. Neurodegenerative disorders treatment: The microRNA role. Curr. Gene Ther. 2017, 17, 327–363. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Liu, D.Z.; Jickling, G.C.; Sharp, F.R.; Yin, K.J. MicroRNA-based therapeutics in central nervous system injuries. J. Cereb. Blood Flow Metab. 2018, 38, 1125–1148. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Wang, Y.W.; Xu, S.H.; Qiao, S.; Shu, Q.F.; Du, J.Z.; Li, Y.G.; Liu, X.L. Regulatory effects of the long noncoding RNA RP11543N12.1 and microRNA3243p axis on the neuronal apoptosis induced by the inflammatory reactions of microglia. Int. J. Mol. Med. 2018, 42, 1741–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higaki, S.; Muramatsu, M.; Matsuda, A.; Matsumoto, K.; Satoh, J.I.; Michikawa, M.; Niida, S. Defensive effect of microRNA-200b/c against amyloid-beta peptide-induced toxicity in Alzheimer’s disease models. PLoS ONE 2018, 13, e0196929. [Google Scholar] [CrossRef]
- Liang, Z.; Chi, Y.J.; Lin, G.Q.; Luo, S.H.; Jiang, Q.Y.; Chen, Y.K. MiRNA-26a promotes angiogenesis in a rat model of cerebral infarction via PI3K/AKT and MAPK/ERK pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3485–3492. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Z.F.; Li, W.; Hong, H.; Chen, J.; Tian, Y.; Liu, Z.Y. Protective effects of microRNA-330 on amyloid beta-protein production, oxidative stress, and mitochondrial dysfunction in Alzheimer’s disease by targeting VAV1 via the MAPK signaling pathway. J. Cell. Biochem. 2018, 119, 5437–5448. [Google Scholar] [CrossRef]
- Heinrich, T.; Hübner, C.A.; Kurth, I. Isolation and primary cell culture of mouse dorsal root ganglion neurons. Biol.-Protocol. 2016, 6. [Google Scholar] [CrossRef]
- Montoya-Gacharna, J.V.; Sutachan, J.J.; Chan, W.S.; Sideris, A.; Blanck, T.J.; Recio-Pinto, E. Preparation of adult spinal cord motor neuron cultures under serum-free conditions. Methods Mol. Biol. 2012, 846, 103–116. [Google Scholar] [CrossRef]
- Brewer, G.J.; Torricelli, J.R. Isolation and culture of adult neurons and neurospheres. Nat. Protoc. 2007, 2, 1490–1498. [Google Scholar] [CrossRef]
- Rzemieniec, J.; Litwa, E.; Wnuk, A.; Lasoń, W.; Gołaś, A.; Krzeptowski, W.; Kajta, M. Neuroprotective action of raloxifene against hypoxia-induced damage in mouse hippocampal cells depends on ERα but not ERβ or GPR30 signalling. J. Steroid Biochem. Mol. Biol. 2015, 146. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaesel, K.; May, C.; Marcus, K.; Matschke, V.; Theiss, C.; Theis, V. miR-129-5p and miR-130a-3p Regulate VEGFR-2 Expression in Sensory and Motor Neurons during Development. Int. J. Mol. Sci. 2020, 21, 3839. https://doi.org/10.3390/ijms21113839
Glaesel K, May C, Marcus K, Matschke V, Theiss C, Theis V. miR-129-5p and miR-130a-3p Regulate VEGFR-2 Expression in Sensory and Motor Neurons during Development. International Journal of Molecular Sciences. 2020; 21(11):3839. https://doi.org/10.3390/ijms21113839
Chicago/Turabian StyleGlaesel, Kevin, Caroline May, Katrin Marcus, Veronika Matschke, Carsten Theiss, and Verena Theis. 2020. "miR-129-5p and miR-130a-3p Regulate VEGFR-2 Expression in Sensory and Motor Neurons during Development" International Journal of Molecular Sciences 21, no. 11: 3839. https://doi.org/10.3390/ijms21113839