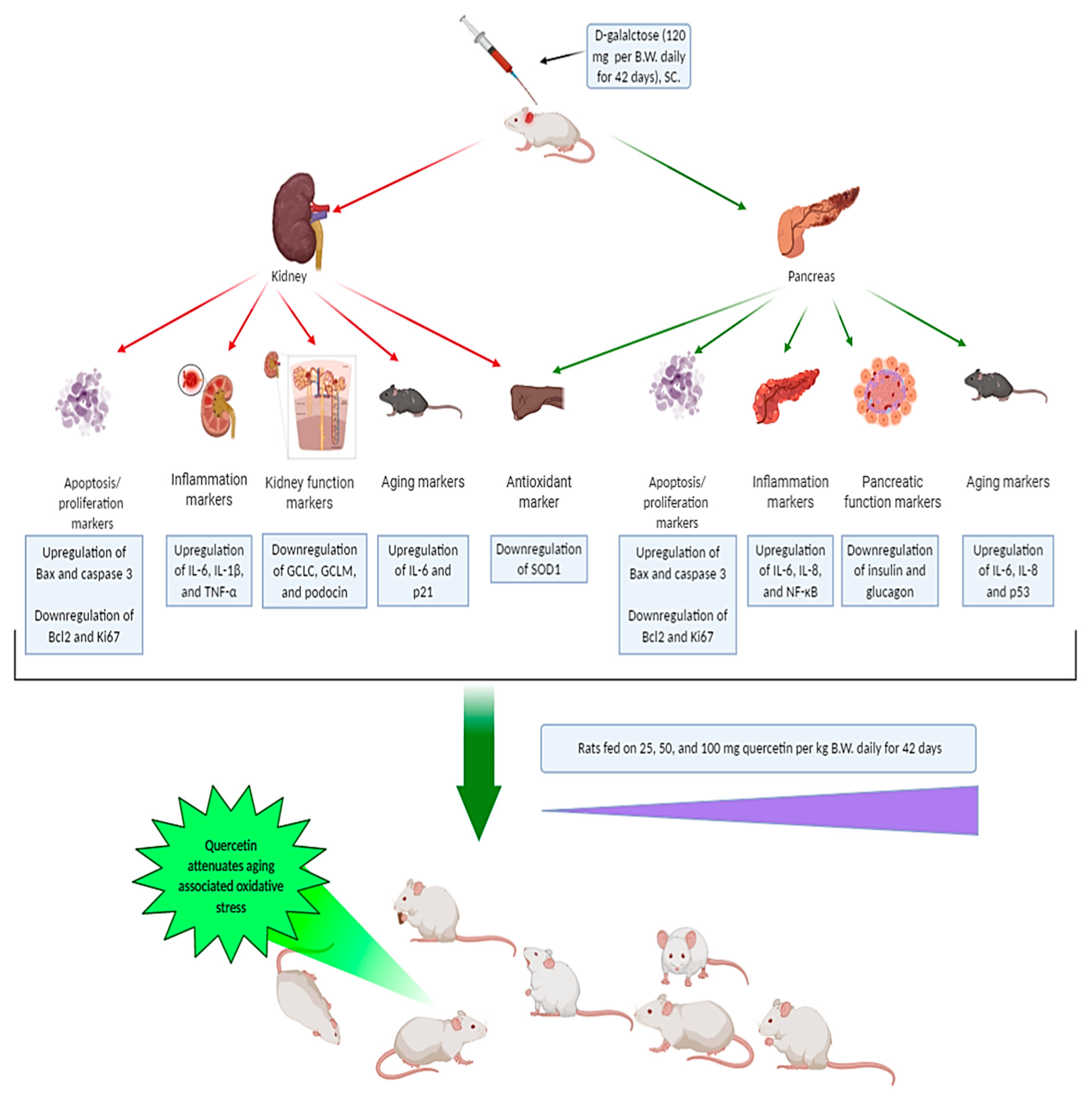
Quercetin Attenuates Pancreatic and Renal D-Galactose-Induced Aging-Related Oxidative Alterations in Rats

 ,
, 
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
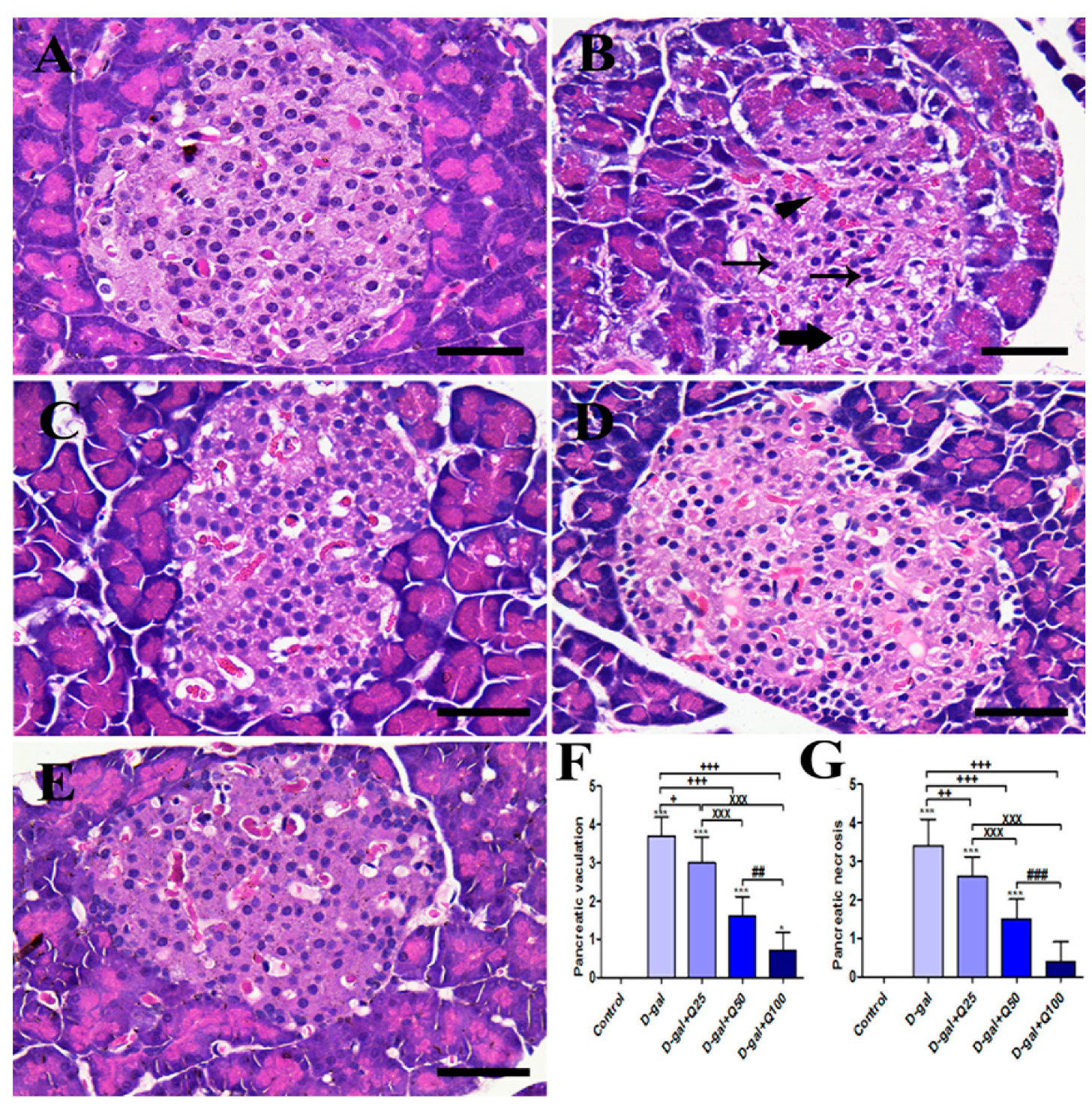
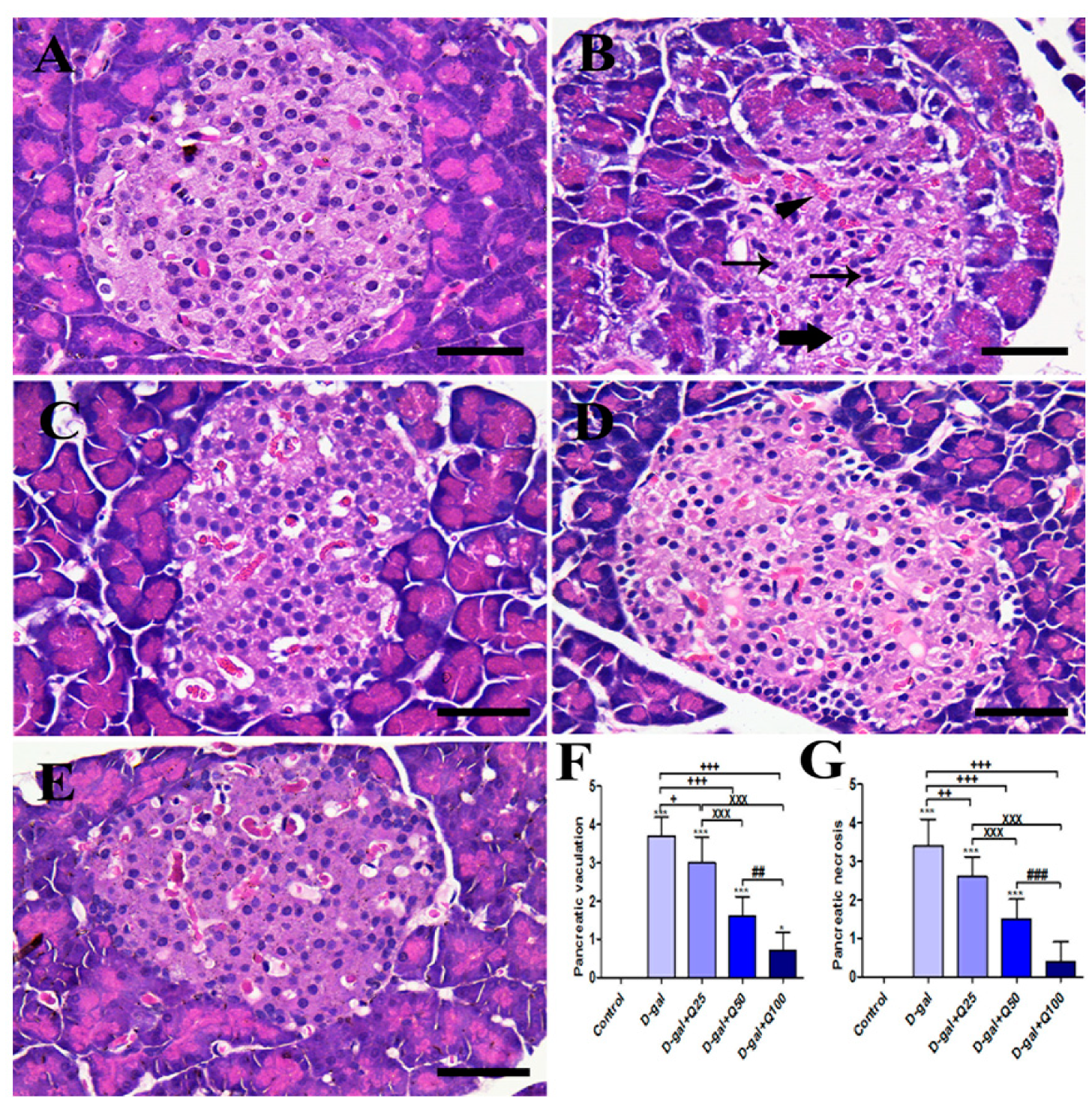
2.1. Pancreatic Histomorphology
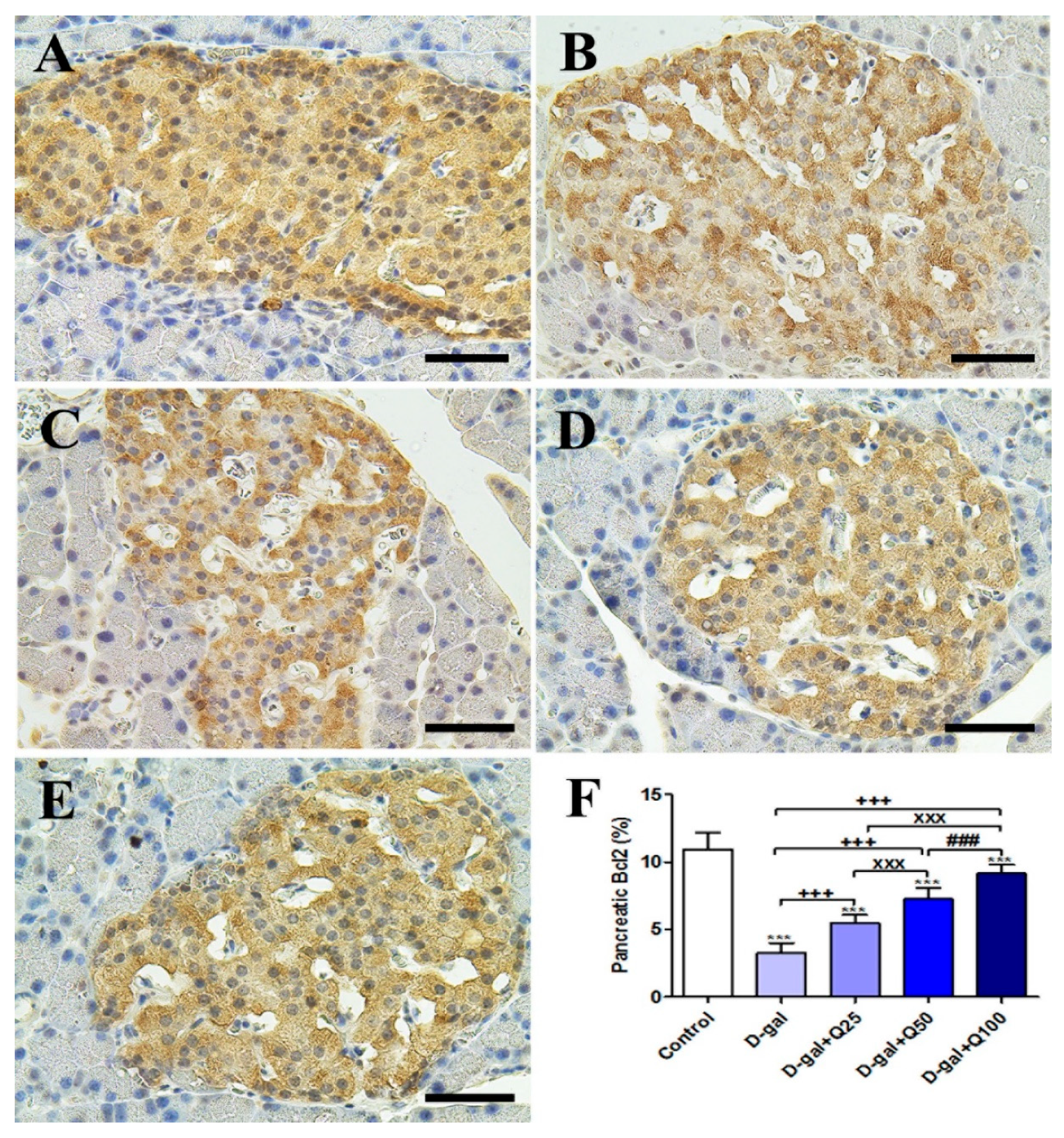
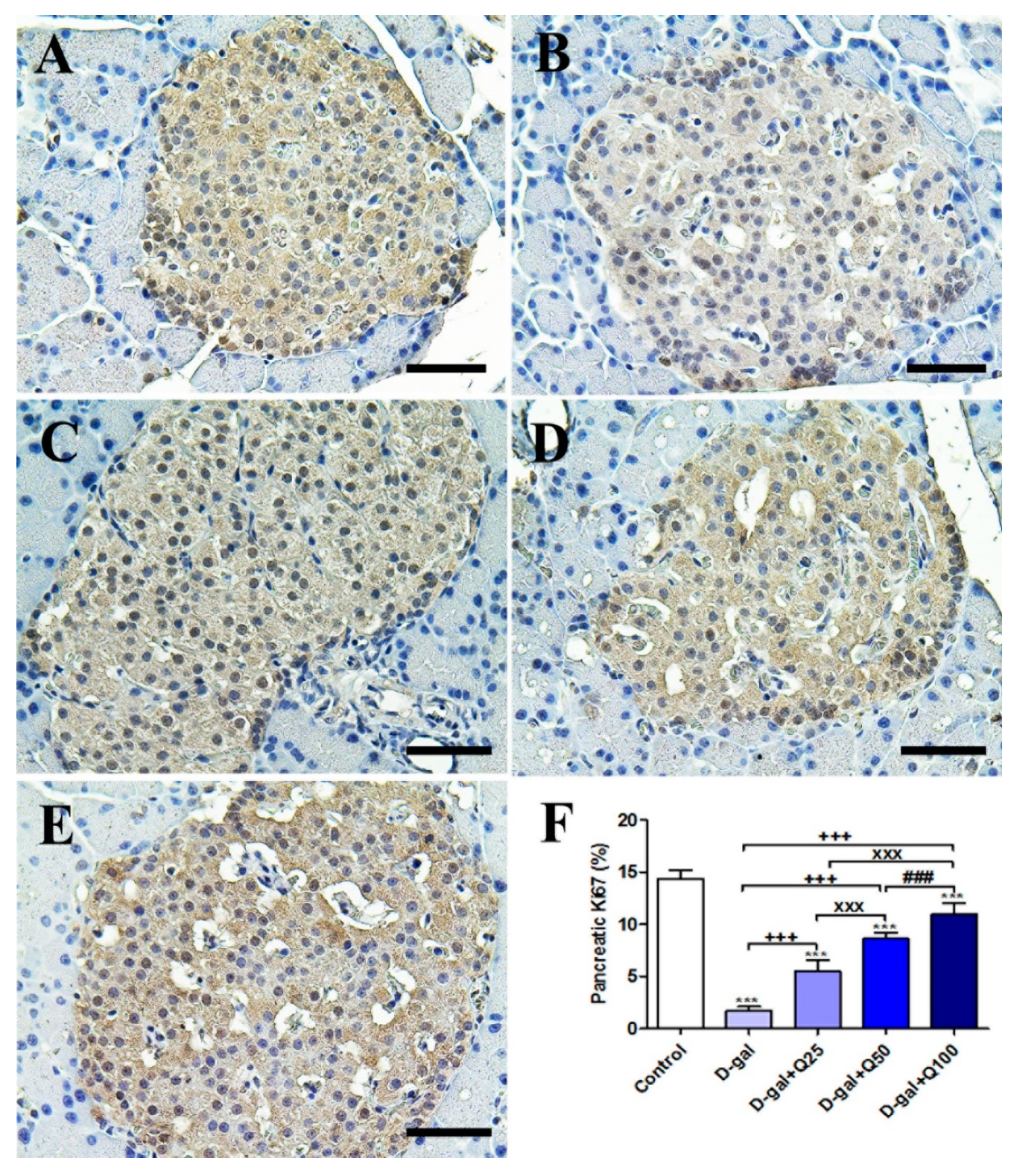
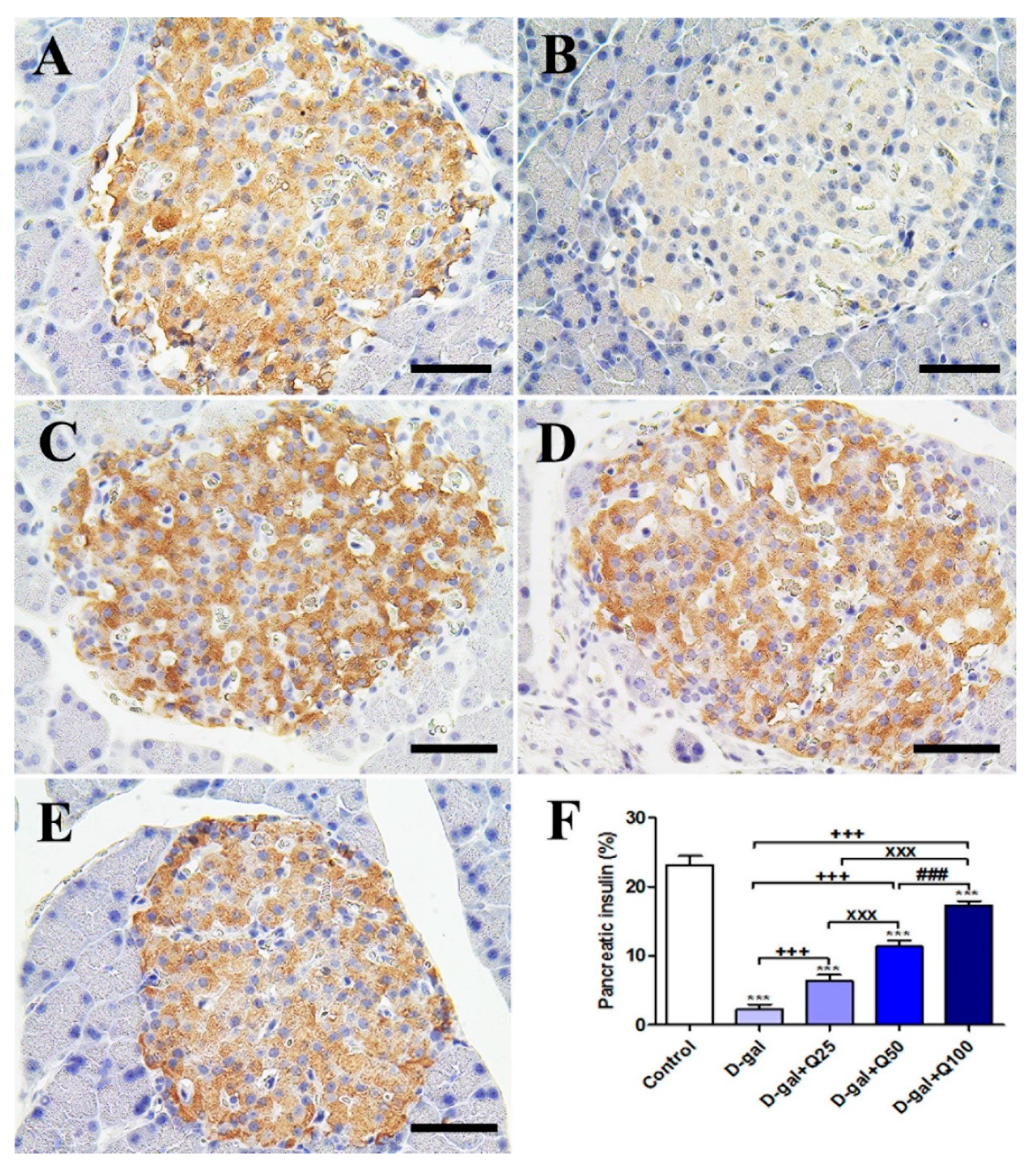
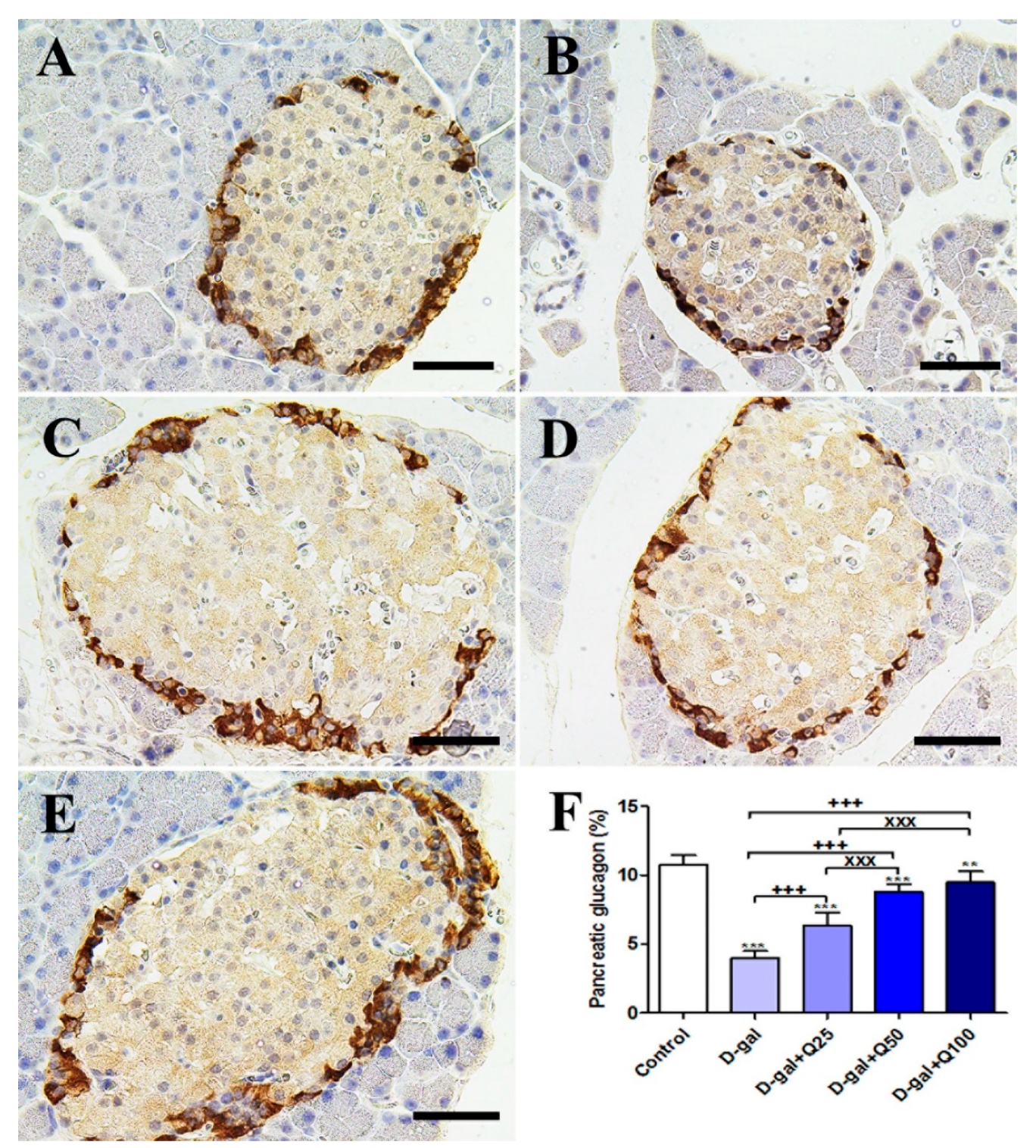
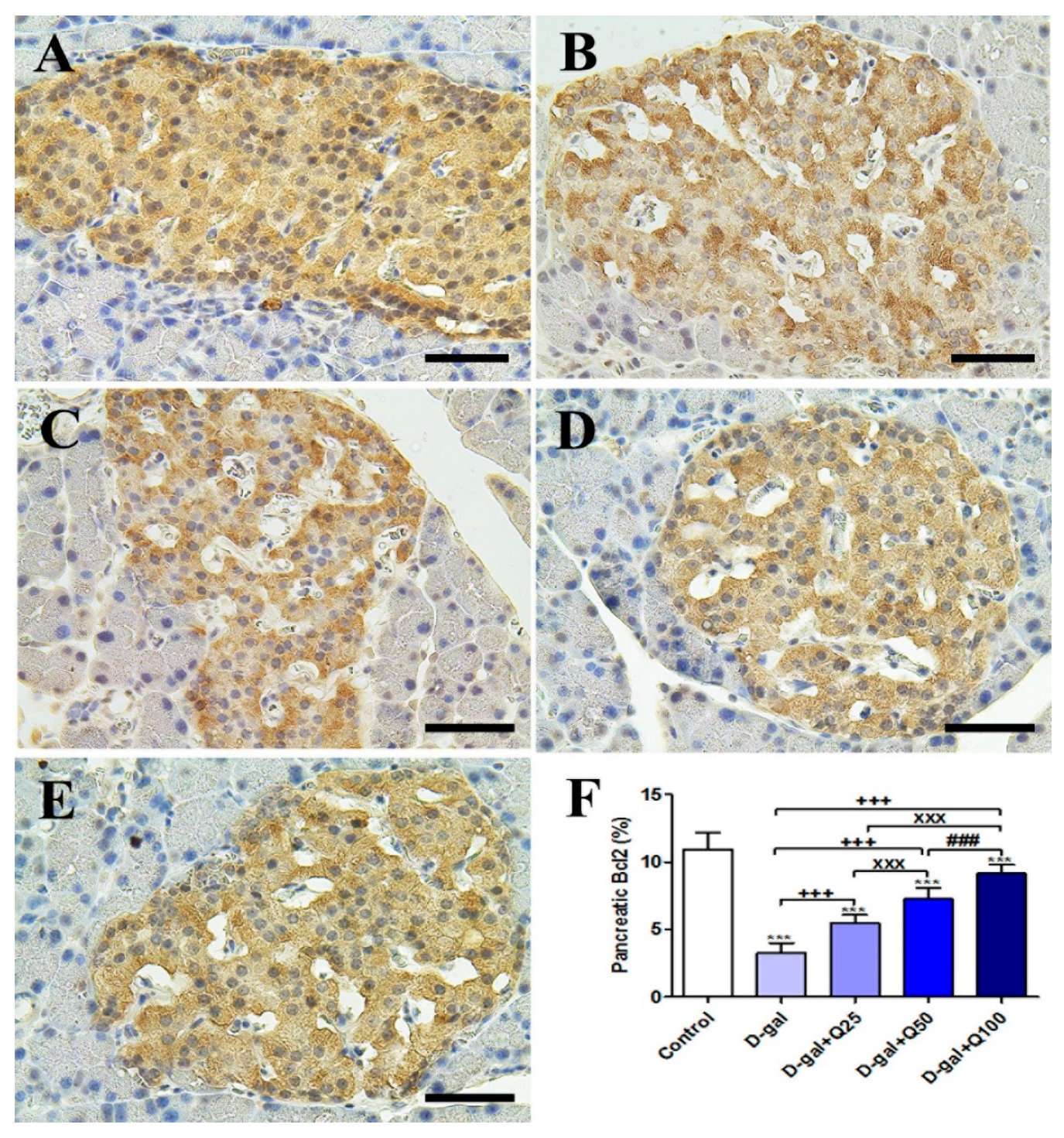
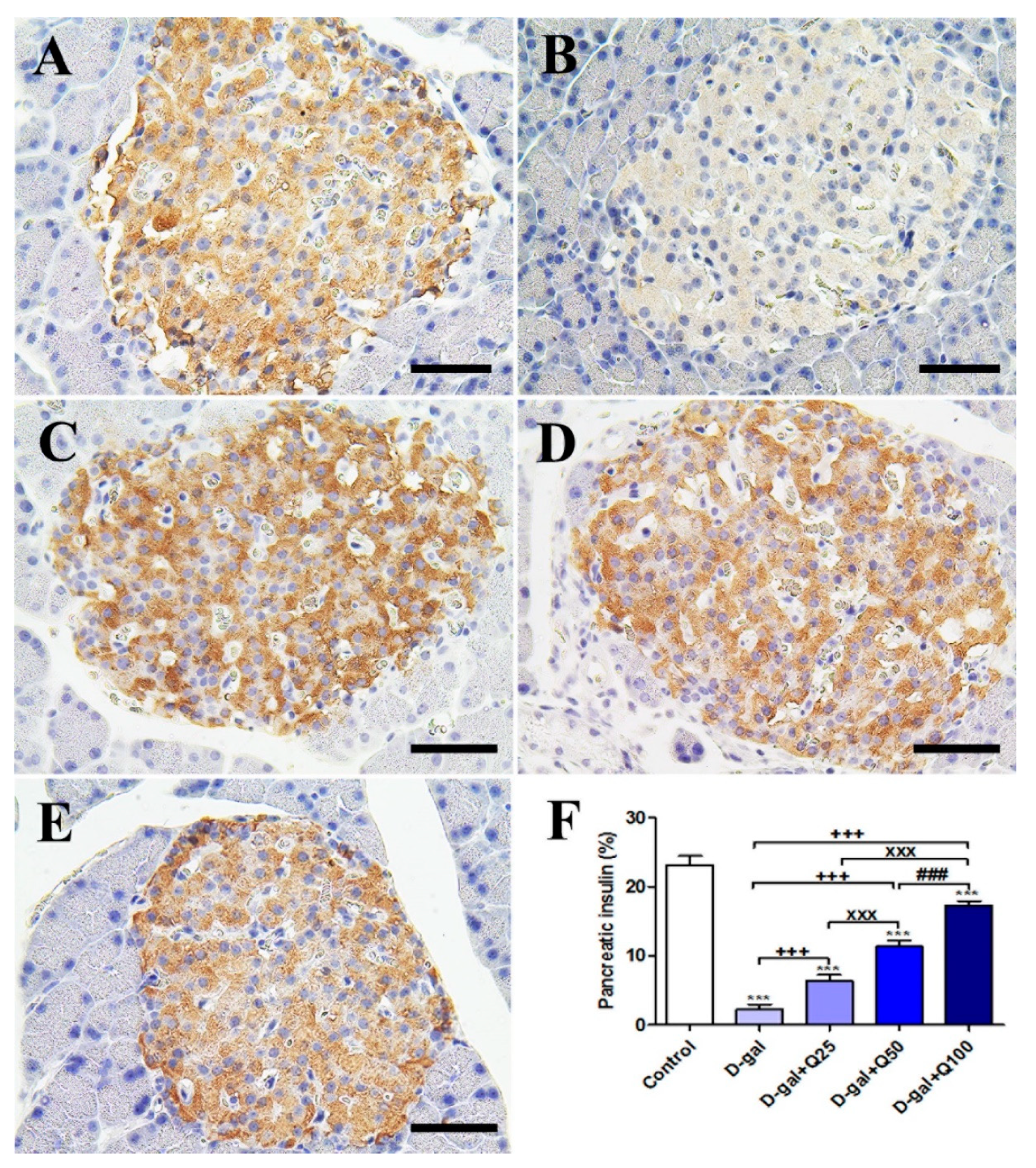
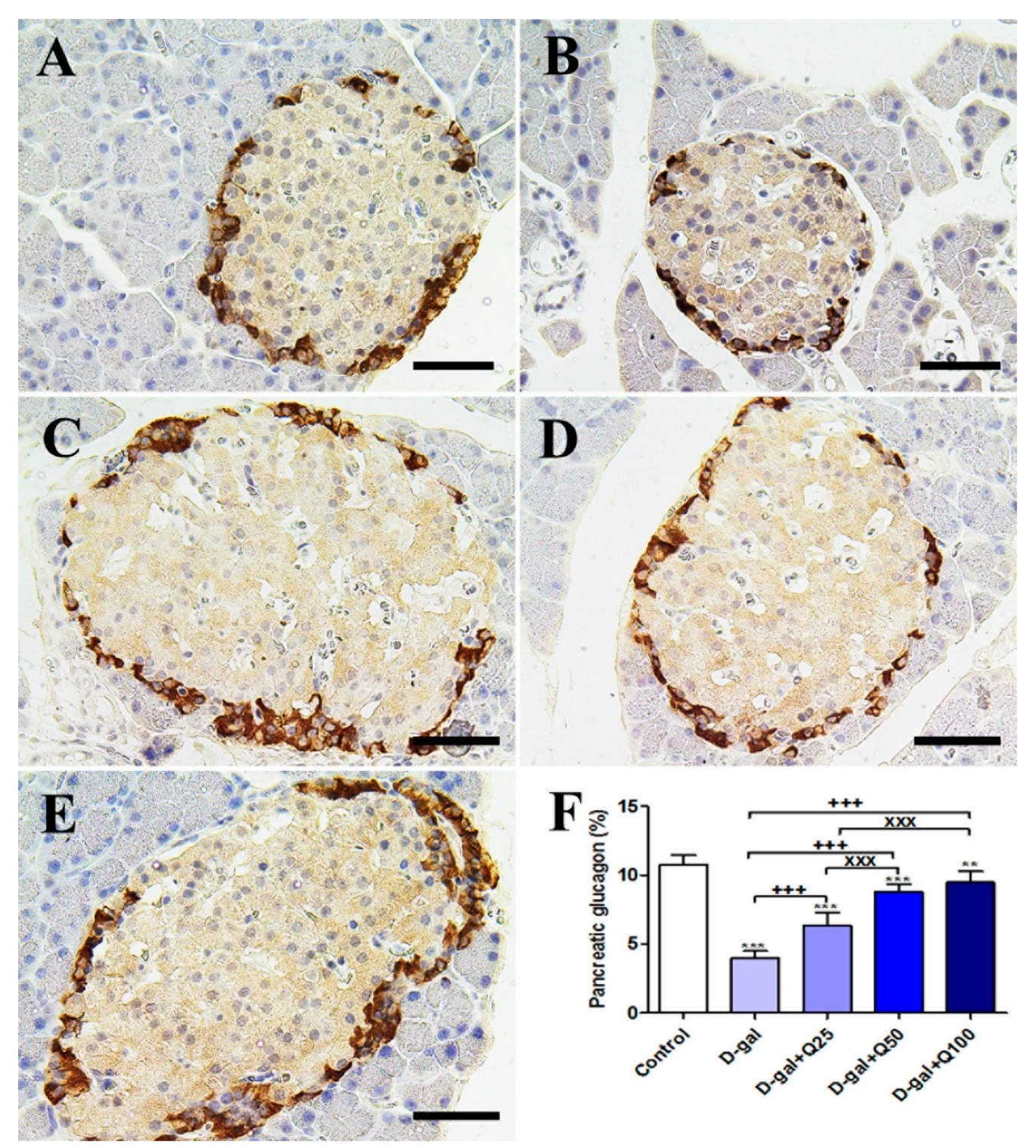
2.2. Pancreatic Immunohistochemistry
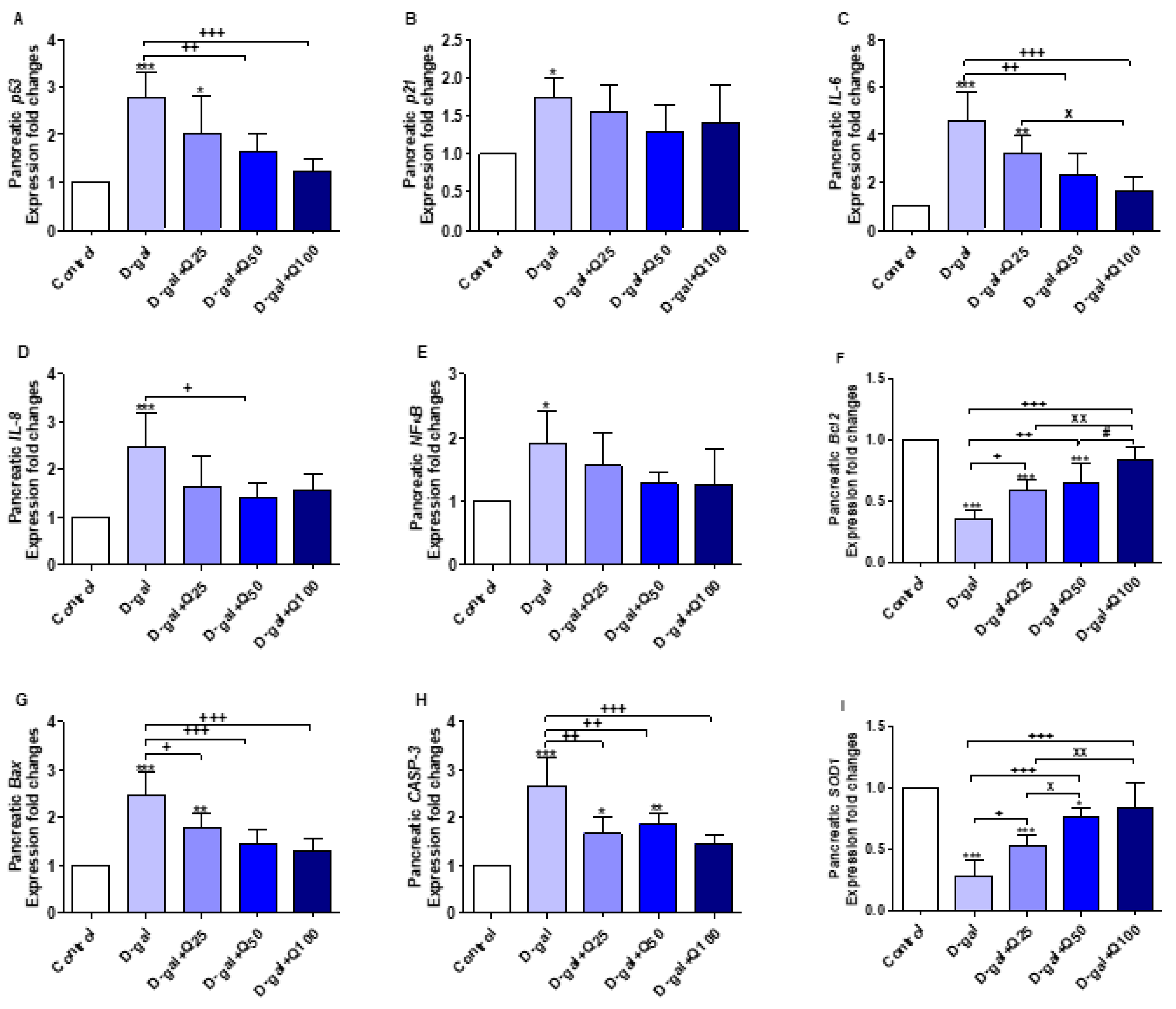
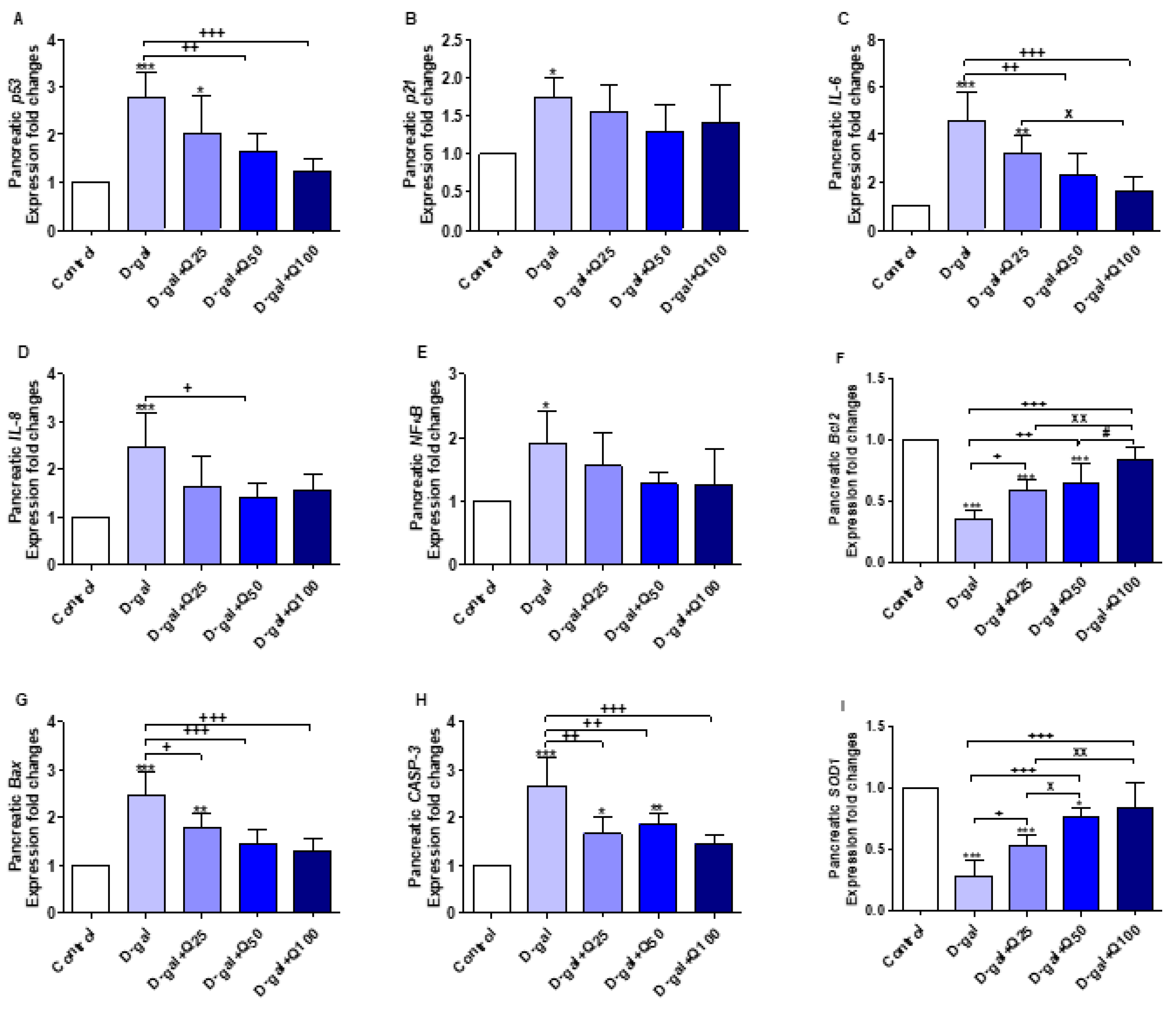
2.3. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) of Pancreatic Aging, Apoptotic, Anti-Inflammatory and Antioxidant Markers
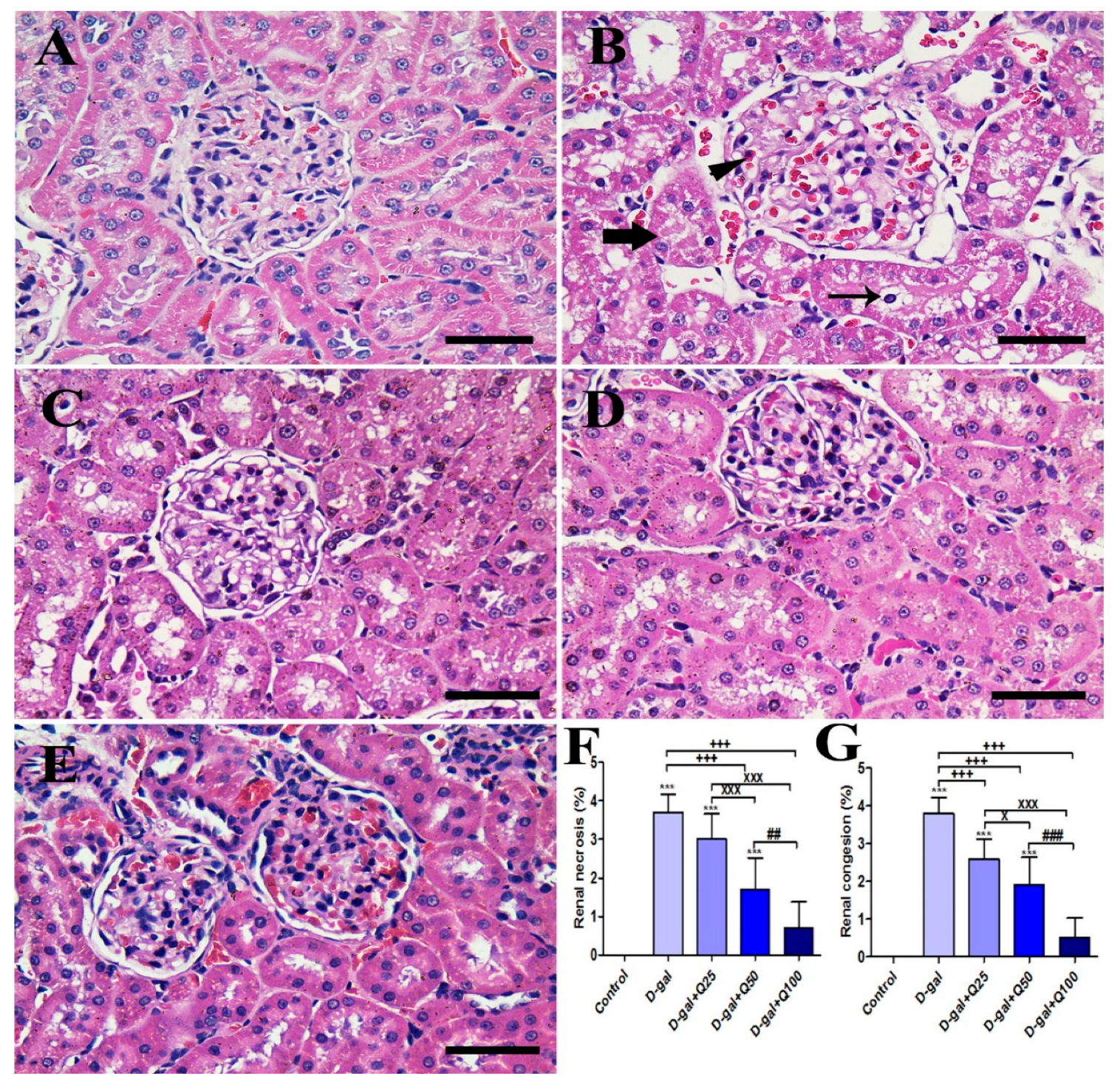
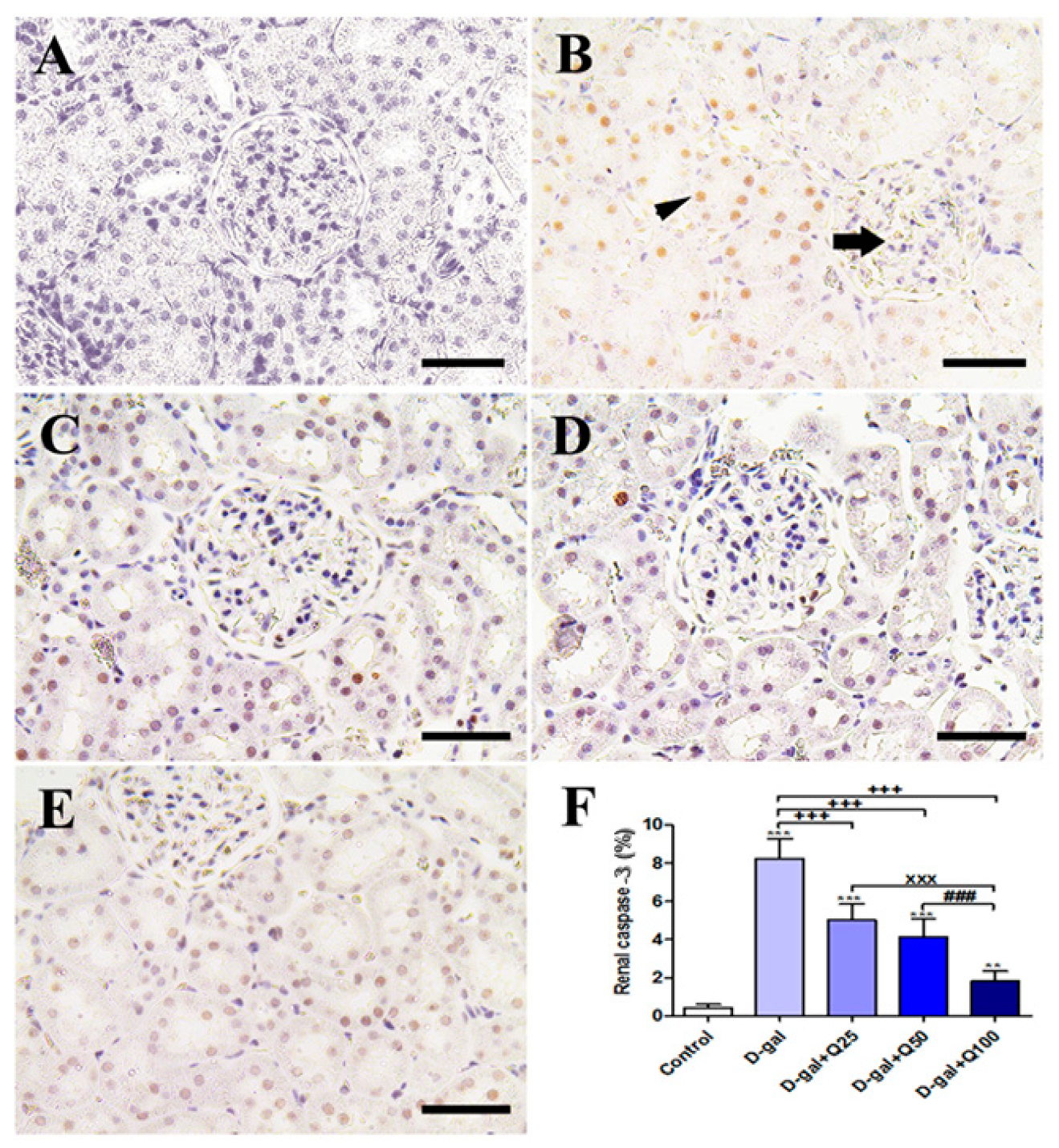
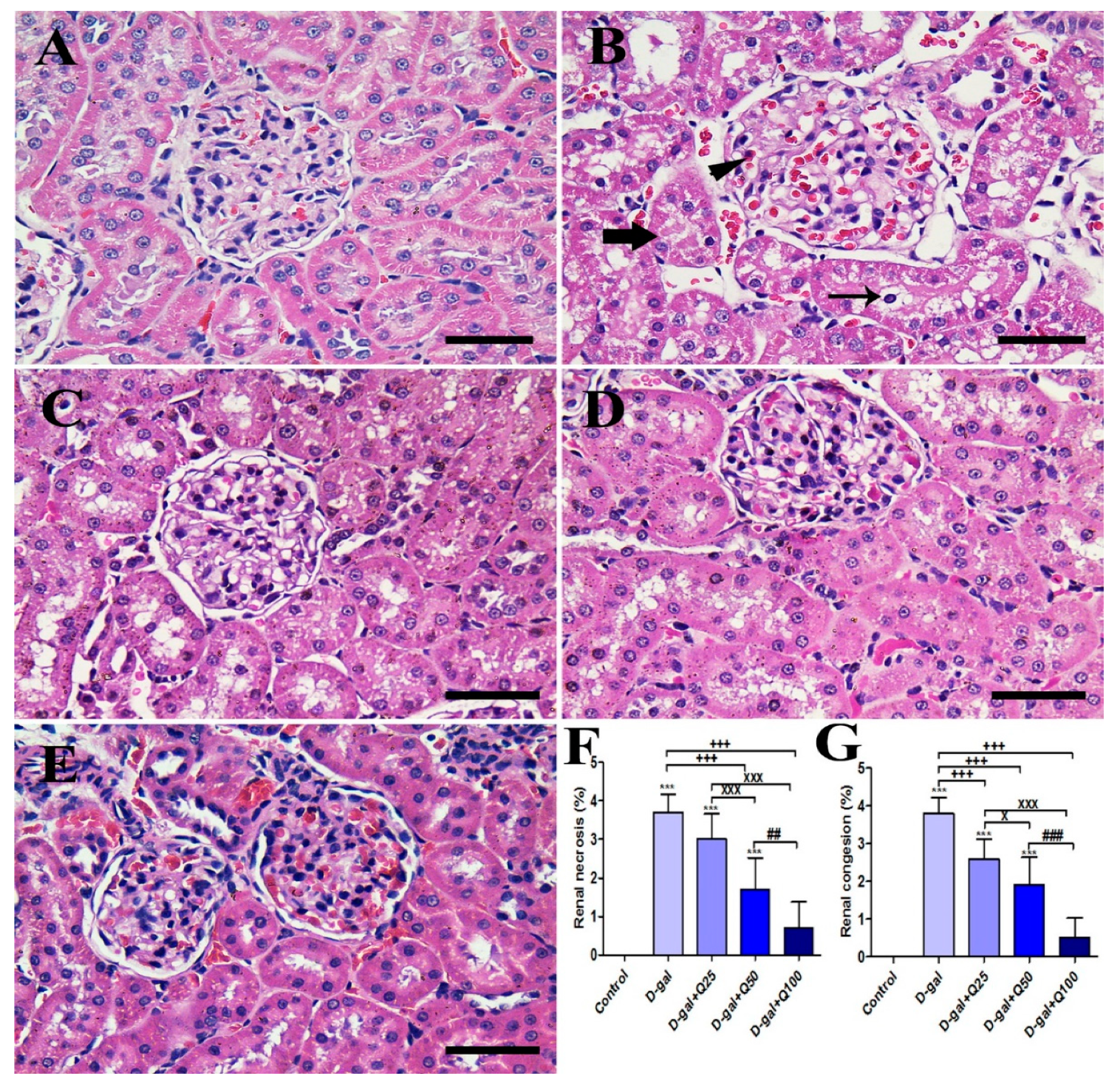
2.4. Renal Histomorphology
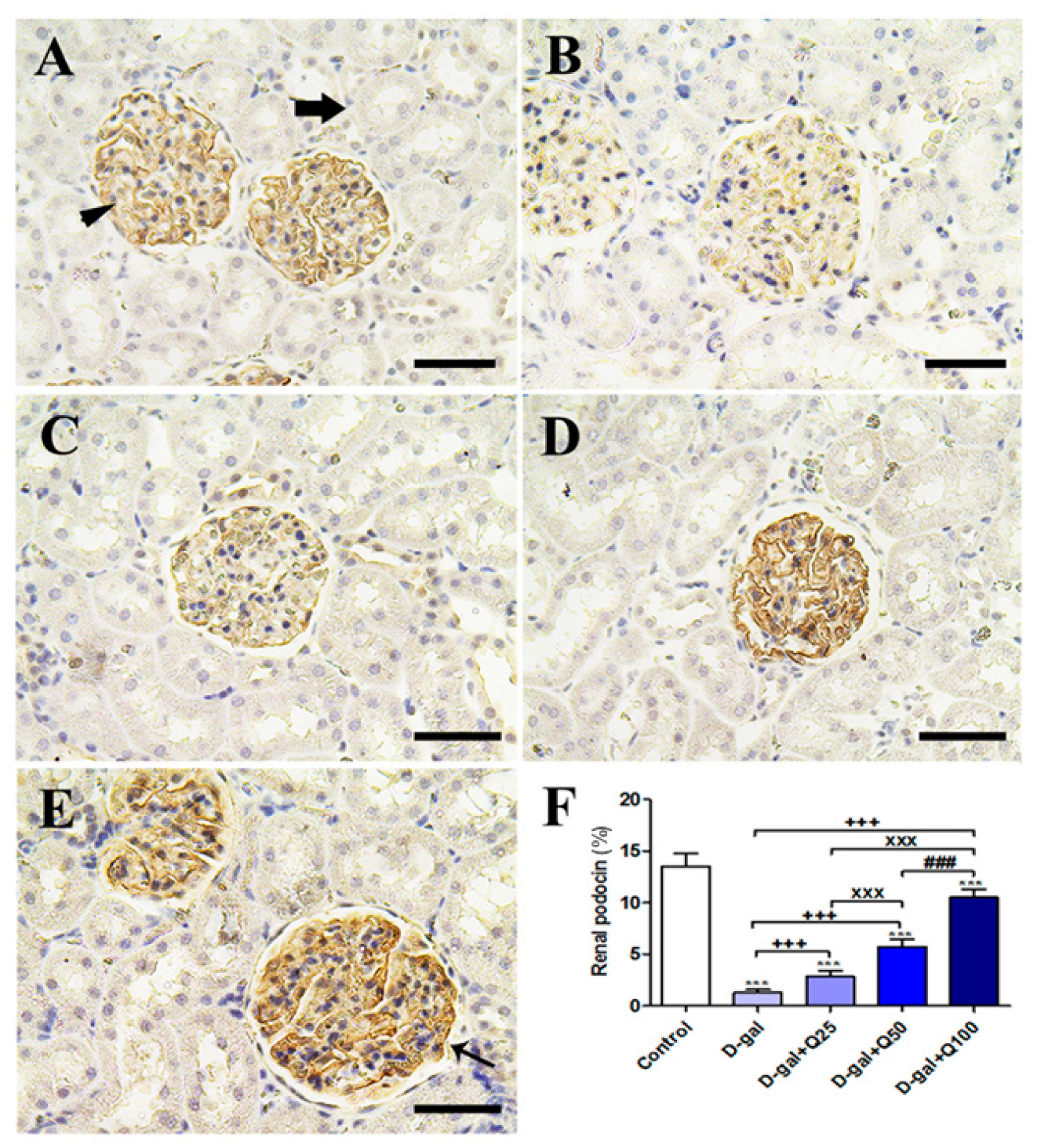
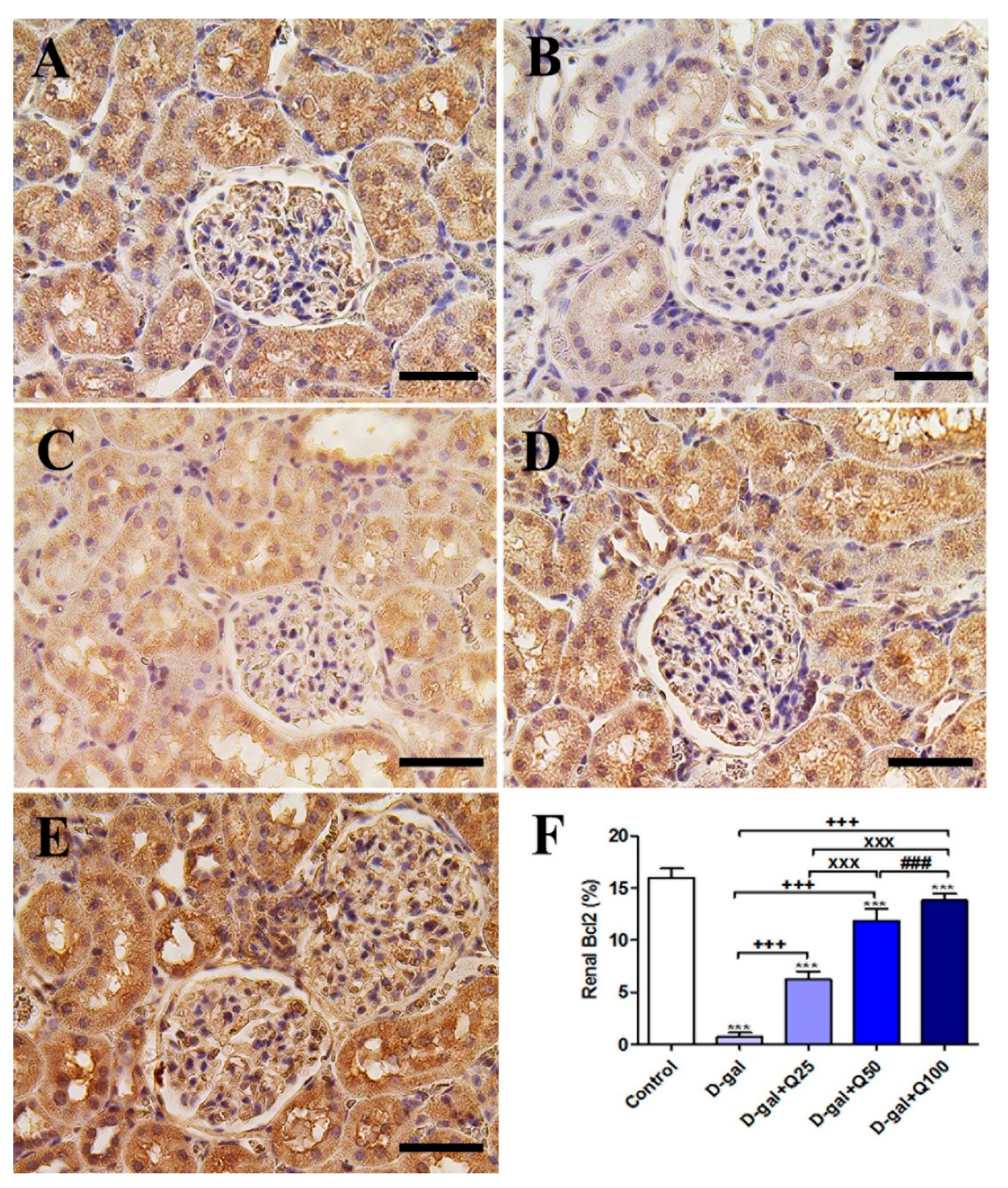
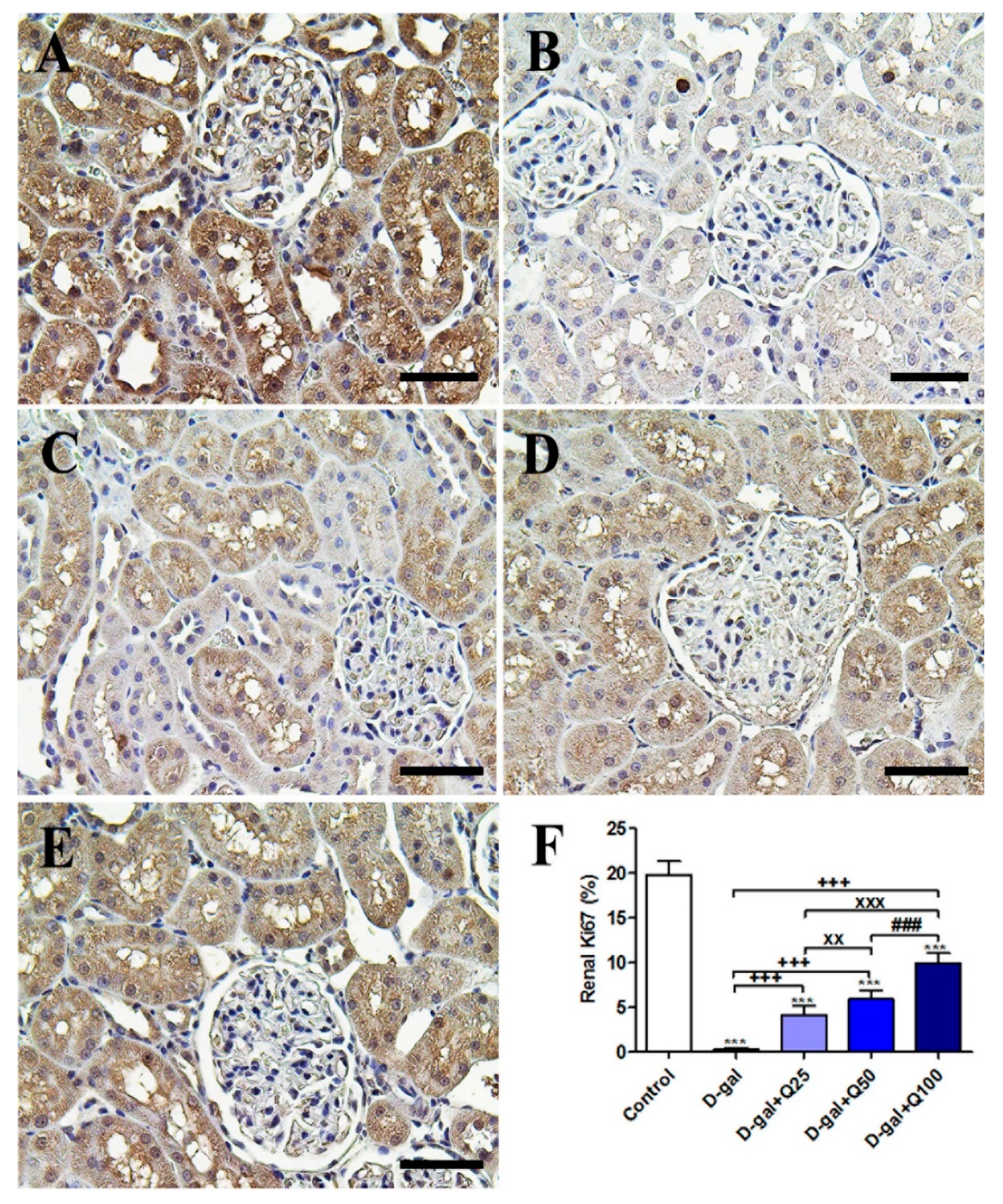
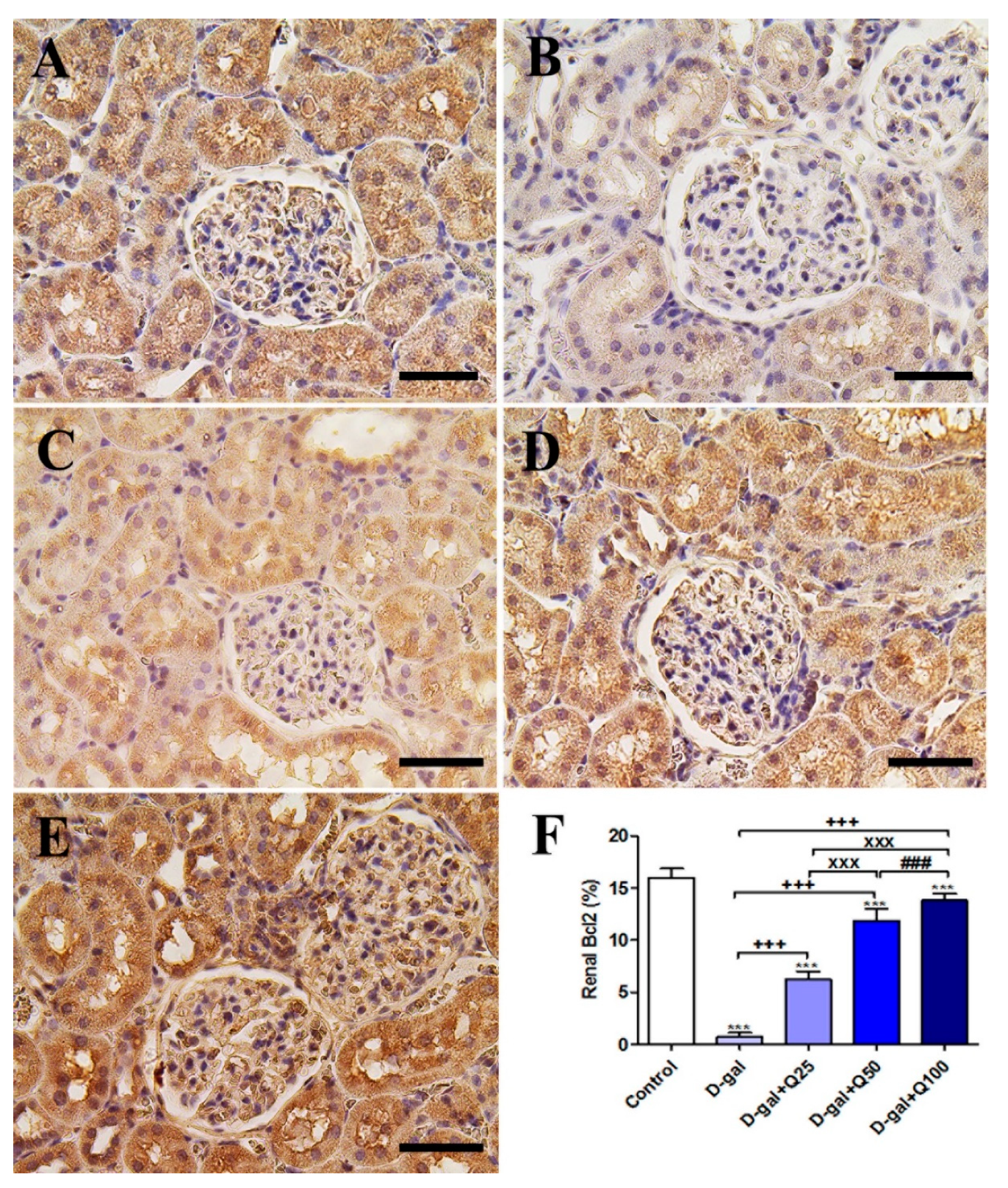
2.5. Renal Immunohistochemistry
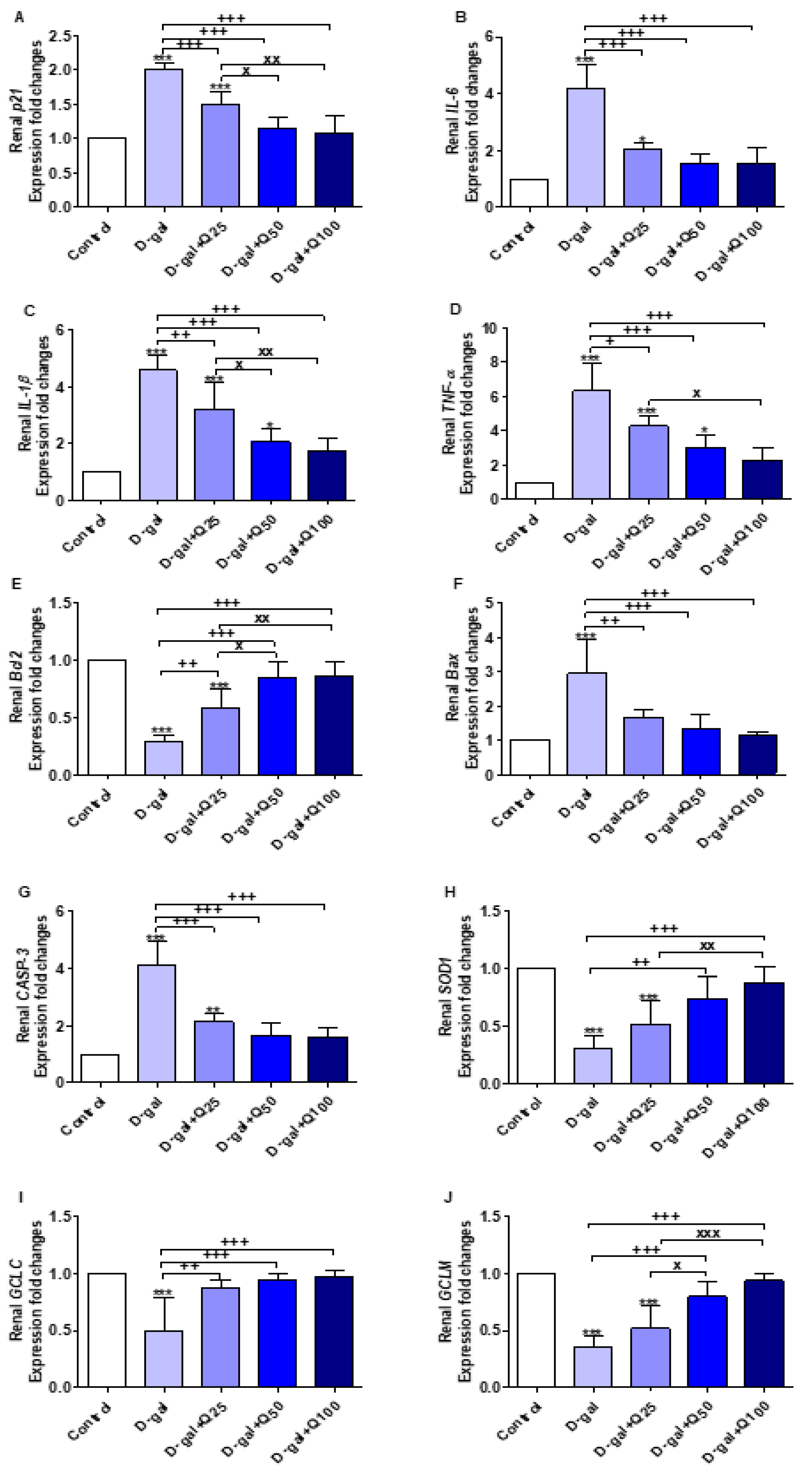
2.6. RT-PCR of Renal Aging, Apoptotic, Anti-Inflammatory, Antioxidant and Functional Markers
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Design
4.3. Sampling
4.4. Histopathological Assessment
4.5. Immunohistochemistry
4.6. RNA Extraction and RT-PCR
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Bax. | Bcl-2-associated X protein |
| Bcl2 | B-cell lymphoma 2 |
| CASP-3 | Caspase-3 |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GCLC | Glutamate-cysteine ligase catalytic subunit |
| GCLM | Glutamate-cysteine ligase regulatory subunit |
| GSH | Reduced glutathione |
| IL-1β | Interleukin-1beta |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| MDA | Malondialdehyde |
| NF-κB | Nuclear factor-kappa B |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| ROS | Reactive oxygen species |
| SOD1 | Cu, Zn-superoxide dismutase |
| TNF-α | Tumor necrosis factor-alpha |
References
- Chen, W.K.; Tsai, Y.L.; Shibu, M.A.; Shen, C.Y.; Chang-Lee, S.N.; Chen, R.J.; Yao, C.H.; Ban, B.; Kuo, W.W.; Huang, C.Y. Exercise training augments Sirt1-signaling and attenuates cardiac inflammation in D-galactose induced-aging rats. Aging 2018, 10, 4166. [Google Scholar] [CrossRef] [PubMed]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, Cell Senescence, and Novel Molecular Mechanisms in Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 3565127. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Zhou, Y.; Gao, L.; Qin, X.; Du, G. Advances in the study of the rat model of aging induced by D-galactose. Acta Pharm. Sin. 2017, 52, 347–354. [Google Scholar]
- Ma, Y.; Ma, B.; Shang, Y.; Yin, Q.; Wang, D.; Xu, S.; Hong, Y.; Hou, X.; Liu, X. Flavonoid-Rich Ethanol Extract from the Leaves of Diospyros kaki Attenuates D-Galactose-Induced Oxidative Stress and Neuroinflammation-Mediated Brain Aging in Mice. Oxidative Med. Cell. Longev. 2018, 2018, 8938207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. Biomed Res. Int. 2019, 2019, 8748253. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef] [Green Version]
- Boots, A.W.; Haenen, G.R.M.M.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Bansal, M.; Singh, N.; Pal, S.; Dev, I.; Ansari, K.M. Chapter Three—Chemopreventive Role of Dietary Phytochemicals in Colorectal Cancer. In Advances in Molecular Toxicology; Elsevier: Washington, DC, USA, 2018; Volume 12, pp. 69–121. [Google Scholar]
- El-Far, A.H.; Ahmed, H.A.; Shaheen, H.M. Dietary Supplementation of Phoenix dactylifera Seeds Enhances Performance, Immune Response, and Antioxidant Status in Broilers. Oxidative Med. Cell. Longev. 2016, 2016, 5454963. [Google Scholar] [CrossRef] [Green Version]
- El-Far, A.H.; Oyinloye, B.E.; Sepehrimanesh, M.; Allah, M.A.G.; Abu-Reidah, I.; Shaheen, H.M.; Razeghian-Jahromi, I.; Alsenosy, A.A.; Noreldin, A.E.; Al Jaouni, S.K.; et al. Date Palm (Phoenix dactylifera): Novel Findings and Future Directions for Food and Drug Discovery. Curr. Drug Discov. Technol. 2019, 16, 2–10. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Heijnen, C.G.M.; Haenen, G.R.M.M.; Oostveen, R.M.; Stalpers, E.M.; Bast, A. Protection of flavonoids against lipid peroxidation: The structure activity relationship revisited. Free Radic. Res. 2002, 36, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darband, S.G.; Sadighparvar, S.; Yousefi, B.; Kaviani, M.; Ghaderi-Pakdel, F.; Mihanfar, A.; Rahimi, Y.; Mobaraki, K.; Majidinia, M. Quercetin attenuated oxidative DNA damage through NRF2 signaling pathway in rats with DMH induced colon carcinogenesis. Life Sci. 2020, 117584. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, N.F.; Eitah, H.E.; Maklad, Y.A.; Gamaleldin, A.A.; Badawi, M.A.; Kenawy, S.A. New combination therapy of gliclazide and quercetin for protection against STZ-induced diabetic rats. Life Sci. 2020, 247. [Google Scholar] [CrossRef] [PubMed]
- Saccol, R.S.P.; da Silveira, K.L.; Manzoni, A.G.; Abdalla, F.H.; de Oliveira, J.S.; Dornelles, G.L.; Barbisan, F.; Passos, D.F.; Casali, E.A.; de Andrade, C.M.; et al. Antioxidant, hepatoprotective, genoprotective, and cytoprotective effects of quercetin in a murine model of arthritis. J. Cell. Biochem. 2019, 121. [Google Scholar] [CrossRef]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for Valid Histopathologic Scoring in Research. Vet. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Chen, J. The Cell-Cycle Arrest and Apoptotic Functions of p53 in Tumor Initiation and Progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef]
- Fujita, K. P53 isoforms in cellular senescence-and ageing-associated biological and physiological functions. Int. J. Mol. Sci. 2019, 20, 6023. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.; Weinberger, B.; Arnold, C.R.; Maier, A.B.; Westendorp, R.G.J.; Grubeck-Loebenstein, B. The effect of chronological age on the inflammatory response of human fibroblasts. Exp. Gerontol. 2012, 47, 749–753. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Qi, M.; Konermann, A.; Zhang, L.; Jin, F.; Jin, Y. The p53/miR-17/Smurf1 pathway mediates skeletal deformities in an age-related model via inhibiting the function of mesenchymal stem cells. Aging 2015, 7, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Cai, Z.; Wang, W.; Wei, M.; Si, X.; Shang, Y.; Yang, Z.; Li, T.; Guo, H.; Li, S. Piperine regulates glycogen synthase kinase-3β-related signaling and attenuates cognitive decline in D-galactose-induced aging mouse model. J. Nutr. Biochem. 2020, 75, 108261. [Google Scholar] [CrossRef]
- Zhong, J.; Wang, F.; Wang, Z.; Shen, C.; Zheng, Y.; Ma, F.; Zhu, T.; Chen, L.; Tang, Q.; Zhu, J. Aloin attenuates cognitive impairment and inflammation induced by D-galactose via down-regulating ERK, p38 and NF-κB signaling pathway. Int. Immunopharmacol. 2019, 72, 48–54. [Google Scholar] [CrossRef]
- Yu, Y.; Bai, F.; Wang, W.; Liu, Y.; Yuan, Q.; Qu, S.; Zhang, T.; Tian, G.; Li, S.; Li, D.; et al. Fibroblast growth factor 21 protects mouse brain against D-galactose induced aging via suppression of oxidative stress response and advanced glycation end products formation. Pharmacol. Biochem. Behav. 2015, 133, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Xu, M.; Huang, J.; Chen, L.; Xia, J.; Chen, X.; Jiang, R.; Wang, L.; Wang, Y. The protective effect of Ginsenoside Rg1 on aging mouse pancreas damage induced by D-galactose. Exp. Ther. Med. 2017, 14, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.Z.; Lin, Y.J.; Li, K.J.; Zhang, G.L.; Zhao, Y.F.; Wang, M.M.; Wei, J.B.; Wei, J.; Hu, G. Effects of rhein lysinate on D-galactose-induced aging mice. Exp. Ther. Med. 2016, 11, 303–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Yu, Y.H.; Wang, S.T.; Ren, J.; Camer, D.; Hua, Y.Z.; Zhang, Q.; Huang, J.; Xue, D.L.; Zhang, X.F.; et al. Chlorogenic acid protects d -galactose-induced liver and kidney injury via antioxidation and anti-inflammation effects in mice. Pharm. Biol. 2016, 54, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- El-Horany, H.E.-S.; Gaballah, H.H.; Helal, D.S. Berberine ameliorates renal injury in a rat model of D-galactose-induced aging through a PTEN/Akt-dependent mechanism. Arch. Physiol. Biochem. 2020, 126, 157–165. [Google Scholar] [CrossRef]
- Nobakht-Haghighi, N.; Rahimifard, M.; Baeeri, M.; Rezvanfar, M.A.; Moini Nodeh, S.; Haghi-Aminjan, H.; Hamurtekin, E.; Abdollahi, M. Regulation of aging and oxidative stress pathways in aged pancreatic islets using alpha-lipoic acid. Mol. Cell. Biochem. 2018, 449, 267–276. [Google Scholar] [CrossRef]
- Kassem, M.; Niazi, Z.R.; Abbas, M.; El Habhab, A.; Kreutter, G.; Khemais-Benkhiat, S.; Auger, C.; Antal, M.C.; Schini-Kerth, V.B.; Toti, F.; et al. Senescence of pancreas in middle-aged rats with normal vascular function. Ann. Transplant. 2017, 22, 177–186. [Google Scholar] [CrossRef]
- Riccillo, F.L.; Bracamonte, M.I.; Cónsole, G.M.; Gómez Dumm, C.L.A. Histomorphological and quantitative immunohistochemical changes in the rat pancreas during aging. Biocell 2004, 28, 127–134. [Google Scholar]
- Ahangarpour, A.; Oroojan, A.A.; Badavi, M. Exendin-4 protects mice from D-galactose-induced hepatic and pancreatic dysfunction. Pathobiol. Aging Age-Relat. Dis. 2018, 8, 1418593. [Google Scholar] [CrossRef] [Green Version]
- Kenawy, S.; Hegazy, R.; Hassan, A.; El-Shenawy, S.; Gomaa, N.; Zaki, H.; Attia, A. Involvement of insulin resistance in D-galactose-induced age-related dementia in rats: Protective role of metformin and saxagliptin. PLoS ONE 2017, 12, e0183565. [Google Scholar] [CrossRef] [PubMed]
- Sumida, Y.; Shima, T.; Shirayama, K.; Misaki, M.; Miyaji, K. Effects of hexoses and their derivatives on glucagon secretion from isolated perfused rat pancreas. Horm. Metab. Res. 1994, 26, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Bonegio, R.; Lieberthal, W. Role of apoptosis in the pathogenesis of acute renal failure. Curr. Opin. Nephrol. Hypertens. 2002, 11, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, C.S.; Min, J.; Lee, S.H.; Jung, Y.S. A high-fat diet increases oxidative renal injury and protein glycation in D-galactose-induced aging rats and its prevention by Korea red Ginseng. J. Nutr. Sci. Vitaminol. 2014, 60, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miner, J.H. Glomerular basement membrane composition and the filtration barrier. Pediatric Nephrol. 2011, 26, 1413–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pätäri-Sampo, A.; Ihalmo, P.; Holthöfer, H. Molecular basis of the glomerular filtration: Nephrin and the emerging protein complex at the podocyte slit diaphragm. Ann. Med. 2006, 38, 483–492. [Google Scholar] [CrossRef]
- Roselli, S.; Gribouval, O.; Boute, N.; Sich, M.; Benessy, F.; Attié, T.; Gubler, M.C.; Antignac, C. Podocin localizes in the kidney to the slit diaphragm area. Am. J. Pathol. 2002, 160, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, V.; Prasad, N.; Jain, M.; Pandey, R. Reduced podocin expression in minimal change disease and focal segmental glomerulosclerosis is related to the level of proteinuria. Clin. Exp. Nephrol. 2013, 17, 811–818. [Google Scholar] [CrossRef]
- Jim, B.; Ghanta, M.; Qipo, A.; Fan, Y.; Chuang, P.Y.; Cohen, H.W.; Abadi, M.; Thomas, D.B.; He, J.C. Dysregulated Nephrin in Diabetic Nephropathy of Type 2 Diabetes: A Cross Sectional Study. PLoS ONE 2012, 7, e36041. [Google Scholar] [CrossRef]
- Kelly, D.J.; Aaltonen, P.; Cox, A.J.; Rumble, J.R.; Langham, R.; Panagiotopoulos, S.; Jerums, G.; Holthöfer, H.; Gilbert, R.E. Expression of the slit-diaphragm protein, nephrin, in experimental diabetic nephropathy: Differing effects of anti-proteinuric therapies. Nephrol. Dial. Transplant. 2002, 17, 1327–1332. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.E.N.; Xu, D.P.; Li, H.B.; Kitts, D.D. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.F.; Xu, X.R.; Guo, Y.J.; Xia, E.Q.; Li, S.; Wu, S.; Chen, F.; Ling, W.H.; Li, H. Bin Determination of antioxidant property and their lipophilic and hydrophilic phenolic contents in cereal grains. J. Funct. Foods 2012, 4, 906–914. [Google Scholar] [CrossRef]
- Vaithiyanathan, V.; Mirunalini, S. Assessment of antioxidant potential and acute toxicity studies of whole plant extract of pergularia daemia (Forsk). Toxicol. Int. 2015, 22, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunnick, J.K.; Halley, J.R. Toxicity and carcinogenicity studies of quercetin, a natural component of foods. Toxicol. Sci. 1992, 19, 423–431. [Google Scholar] [CrossRef]
- Lu, J.; Zheng, Y.; Luo, L.; Wu, D.; Sun, D.; Feng, Y. Quercetin reverses D-galactose induced neurotoxicity in mouse brain. Behav. Brain Res. 2006, 171, 251–260. [Google Scholar] [CrossRef]
- Dong, F.; Wang, S.; Wang, Y.; Yang, X.; Jiang, J.; Wu, D.; Qu, X.; Fan, H.; Yao, R. Quercetin ameliorates learning and memory via the Nrf2-ARE signaling pathway in D-galactose-induced neurotoxicity in mice. Biochem. Biophys. Res. Commun. 2017, 491, 636–641. [Google Scholar] [CrossRef]
- Ramana, B.V.; Kumar, V.V.; Krishna, P.N.R.; Kumar, C.S.; Reddy, P.U.M.; Raju, T.N. Effect of quercetin on galactose-induced hyperglycaemic oxidative stress in hepatic and neuronal tissues of Wistar rats. Acta Diabetol. 2006, 43, 135–141. [Google Scholar] [CrossRef]
- Atta, M.S.; Almadaly, E.A.E.A.; El-Far, A.H.A.H.; Saleh, R.M.R.M.; Assar, D.H.D.H.; Al Jaouni, S.K.S.K.; Mousa, S.A. Thymoquinone defeats diabetes-induced testicular damage in rats targeting antioxidant, inflammatory and aromatase expression. Int. J. Mol. Sci. 2017, 18, 919. [Google Scholar] [CrossRef]
- Zhu, J.; Mu, X.; Zeng, J.; Xu, C.; Liu, J.; Zhang, M.; Li, C.; Chen, J.; Li, T.; Wang, Y. Ginsenoside Rg1 Prevents Cognitive Impairment and Hippocampus Senescence in a Rat Model of D-Galactose-Induced Aging. PLoS ONE 2014, 9, e101291. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.Y.; Cui, L.; Wang, X.Z.; Zhang, L.; Zhu, D.; Zhou, X.R.; Hao, L.R. Quercetin Attenuates Vascular Calcification through Suppressed Oxidative Stress in Adenine-Induced Chronic Renal Failure Rats. Biomed Res. Int. 2017, 2017. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Layton, C. The Hematoxylin and eosin. In Theory Practice of Histological Techniques, 7th ed.; Suvarna, S.K., Layton, C., Bancroft, J.D., Eds.; Churchill Livingstone: Philadelphia, PA, USA, 2013. [Google Scholar]
- Noreldin, A.E.; Sogabe, M.; Yamano, Y.; Uehara, M.; Mahdy, M.A.A.; Elnasharty, M.A.; Sayed-Ahmed, A.; Warita, K.; Hosaka, Y.Z. Spatial distribution of osteoblast activating peptide in the rat stomach. Acta Histochem. 2016, 118, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Noreldin, A.E.; Elewa, Y.H.A.; Kon, Y.; Warita, K.; Hosaka, Y.Z. Immunohistochemical localization of osteoblast activating peptide in the mouse kidney. Acta Histochem. 2018, 120, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Sysel, A.M.; Valli, V.E.; Nagle, R.B.; Bauer, J.A. Immunohistochemical quantification of the vitamin B12 transport protein (TCII), cell surface receptor (TCII-R) and Ki-67 in human tumor xenografts. Anticancer Res. 2013, 33, 4203–4212. [Google Scholar]
- Vis, A.E.N.; Kranse, R.; Nigg, A.L.; Van Der Kwast, T.H. Quantitative analysis of the decay of immunoreactivity in stored prostate needle biopsy sections. Am. J. Clin. Pathol. 2000, 113, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohseni, M.; Mihandoost, E.; Shirazi, A.; Sepehrizadeh, Z.; Bazzaz, J.T.; Ghazi-Khansari, M. Melatonin may play a role in modulation of bax and bcl-2 expression levels to protect rat peripheral blood lymphocytes from gamma irradiation-induced apoptosis. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2012, 738, 19–27. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Al-Khalifa, M.S.; Al-Quraishy, S.; Zrieq, R.; Moneim, A.E.A. Indigofera oblongifolia mitigates lead-acetateinduced kidney damage and apoptosis in a rat model. Drug Des. Dev. Ther. 2016, 10, 1847–1856. [Google Scholar] [CrossRef]
- Lebda, M.A.; El-Far, A.H.; Noreldin, A.E.; Elewa, Y.H.A.; Al Jaouni, S.K.; Mousa, S.A. Protective Effects of Miswak ( Salvadora persica ) against Experimentally Induced Gastric Ulcers in Rats. Oxidative Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Hafez, M.M.; Hamed, S.S.; El-Khadragy, M.F.; Hassan, Z.K.; Al Rejaie, S.S.; Sayed-Ahmed, M.M.; Al-Harbi, N.O.; Al-Hosaini, K.A.; Al-Harbi, M.M.; Alhoshani, A.R.; et al. Effect of ginseng extract on the TGF-β1 signaling pathway in CCl4-induced liver fibrosis in rats. BMC Complement. Altern. Med. 2017, 17, 45. [Google Scholar] [CrossRef] [Green Version]
- Zaragozá, R.; García, C.; Rus, A.D.; Pallardó, F.V.; Barber, T.; Torres, L.; Miralles, V.J.; Viña, J.R. Inhibition of liver trans-sulphuration pathway by propargylglycine mimics gene expression changes found in the mammary gland of weaned lactating rats: Role of glutathione. Biochem. J. 2003, 373, 825–834. [Google Scholar] [CrossRef]
- Wu, N.; Sarna, L.K.; Siow, Y.L.; Karmin, O. Regulation of hepatic cholesterol biosynthesis by berberine during hyperhomocysteinemia. Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R635–R643. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Far, A.H.; Lebda, M.A.; Noreldin, A.E.; Atta, M.S.; Elewa, Y.H.A.; Elfeky, M.; Mousa, S.A. Quercetin Attenuates Pancreatic and Renal D-Galactose-Induced Aging-Related Oxidative Alterations in Rats. Int. J. Mol. Sci. 2020, 21, 4348. https://doi.org/10.3390/ijms21124348
El-Far AH, Lebda MA, Noreldin AE, Atta MS, Elewa YHA, Elfeky M, Mousa SA. Quercetin Attenuates Pancreatic and Renal D-Galactose-Induced Aging-Related Oxidative Alterations in Rats. International Journal of Molecular Sciences. 2020; 21(12):4348. https://doi.org/10.3390/ijms21124348
Chicago/Turabian StyleEl-Far, Ali H., Mohamed A. Lebda, Ahmed E. Noreldin, Mustafa S. Atta, Yaser H. A. Elewa, Mohamed Elfeky, and Shaker A. Mousa. 2020. "Quercetin Attenuates Pancreatic and Renal D-Galactose-Induced Aging-Related Oxidative Alterations in Rats" International Journal of Molecular Sciences 21, no. 12: 4348. https://doi.org/10.3390/ijms21124348