Three-Dimensional Cell Cultures as an In Vitro Tool for Prostate Cancer Modeling and Drug Discovery


 ,
, 
 and
and
Abstract
1. Introduction
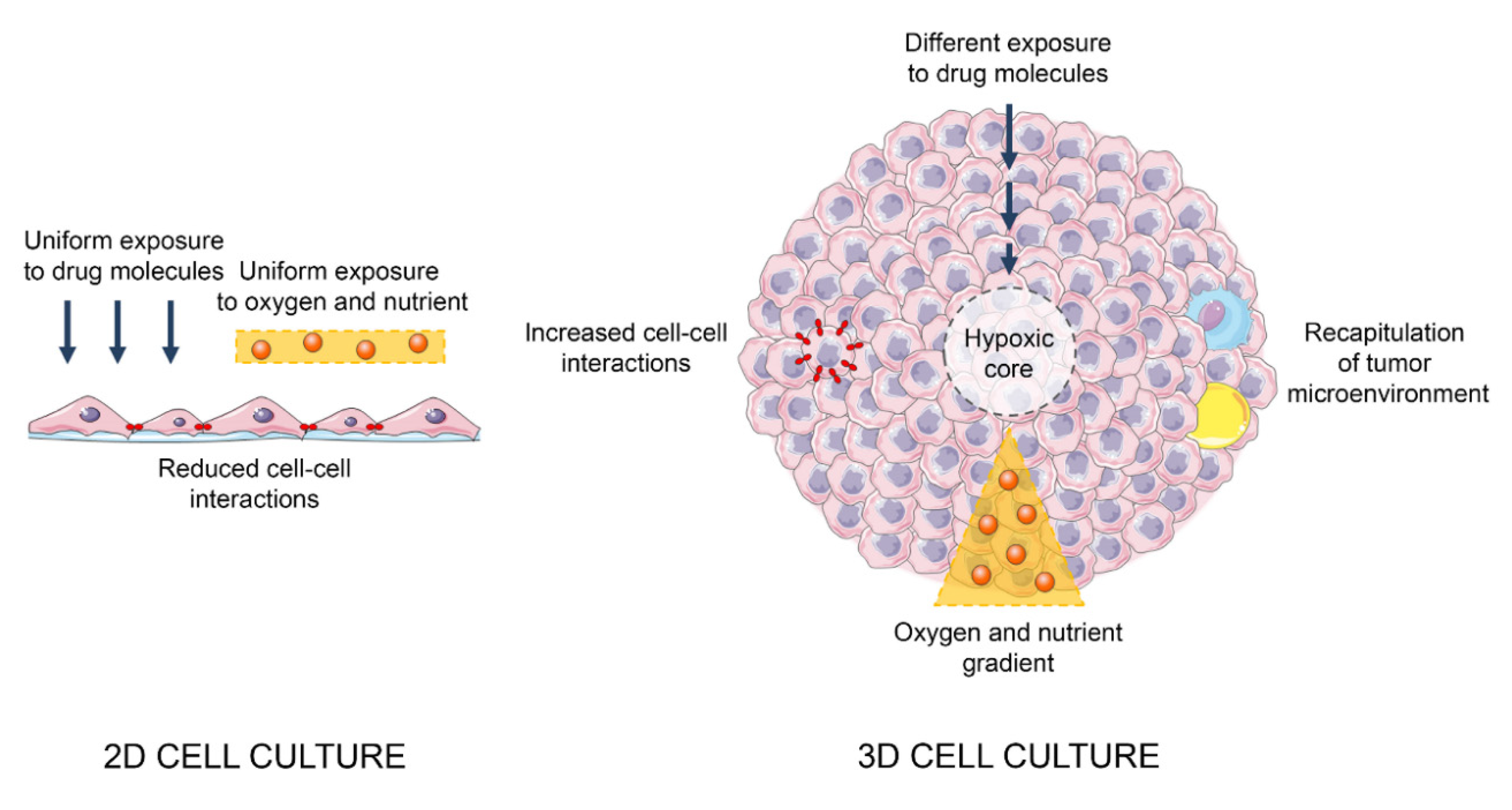
2. 3D Cell Cultures as Preclinical Models of PCa
Methods of 3D PCa Cell Culture
3. Cellular Signaling in 3D PCa Models
3.1. Extracellular Matrix Remodeling and Epithelial-to-Mesenchymal Transition
3.2. Evaluation of Clinical and Diagnostic Features of PCa
3.3. Androgen Signaling
3.4. Assessing Tumor Hypoxia
3.5. Modeling Cancer Stem Cell Plasticity
3.6. Cell Redox Homeostasis and Energetics
3.7. Inflammation
3.8. Angiogenesis
3.9. Metastasis
3.10. PCa and Immunosurveillance
4. 3D Cell Cultures in Drug Discovery and Screening
4.1. Radiotherapy
4.2. Hormone Therapy
4.3. Chemotherapy
4.4. Targeted Therapies
4.5. Novel and Experimental Therapies
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rawla, P. Epidemiology of prostate cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Castelli, G.; Pelosi, E. Cellular and molecular mechanisms underlying prostate cancer development: Therapeutic implications. Medicines 2019, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Teo, M.Y.; Rathkopf, D.E.; Kantoff, P. Treatment of advanced prostate cancer. Annu. Rev. Med. 2019, 70, 479–499. [Google Scholar] [CrossRef] [PubMed]
- Lovitt, C.J.; Shelper, T.B.; Avery, V.M. Advanced cell culture techniques for cancer drug discovery. Biology 2014, 3, 345–367. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.; You, Z. In vitro and in vivo model systems used in prostate cancer research. J. Biol. Methods 2015, 2, 17. [Google Scholar] [CrossRef]
- Jacks, T.; Weinberg, R.A. Taking the study of cancer cell survival to a new dimension. Cell 2002, 111, 923–925. [Google Scholar] [CrossRef]
- Abbott, A. Biology’s new dimension. Nature 2003, 424, 870–872. [Google Scholar] [CrossRef]
- Huh, D.; Matthews, B.D.; Mammoto, A.; Montoya-Zavala, M.; Hsin, H.Y.; Ingber, D.E. Reconstituting organ-level lung functions on a chip. Science 2010, 328, 1662–1668. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L. The principles of humane experimental technique. Med. J. Aust. 1960, 1, 500. [Google Scholar]
- Breslin, S.; O’Driscoll, L. Three-dimensional cell culture: The missing link in drug discovery. Drug Discov. Today 2013, 18, 240–249. [Google Scholar] [CrossRef]
- Lv, D.; Hu, Z.; Lu, L.; Lu, H.; Xu, X. Three-dimensional cell culture: A powerful tool in tumor research and drug discovery. Oncol. Lett. 2017, 14, 6999–7010. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, L.; Lennartsson, L.; Ronquist, G.; Larsson, A.; Nilson, S.; Nilsson, O. Mode of Growth Determines differential expression of prostasomes in cultures of prostate cancer cell lines and opens for studies of prostasome gene expression. Upsala J. Med. Sci. 2006, 111, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Takir, G.G.; Debelec-Butuner, B.; Korkmaz, K.S. 3D cell culture model for prostate cancer cells to mimic inflammatory microenvironment. Proceedings 2018, 2, 1555. [Google Scholar] [CrossRef]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Sommariva, M.; Limonta, P.; Gagliano, N. Epithelial-to-mesenchymal transition markers and cd44 isoforms are differently expressed in 2d and 3d cell cultures of prostate cancer cells. Cells 2019, 8, 143. [Google Scholar] [CrossRef] [PubMed]
- Enmon, R.M.; O’Connor, K.C.; Lacks, D.J.; Schwartz, D.K.; Dotson, R.S. Dynamics of spheroid self-assembly in liquid-overlay culture of DU 145 human prostate cancer cells. Biotechnol. Bioeng. 2001, 72, 579–591. [Google Scholar] [CrossRef]
- Enmon, R.M.; O’Connor, K.C.; Song, H.; Lacks, D.J.; Schwartz, D.K. Aggregation kinetics of well and poorly differentiated human prostate cancer cells. Biotechnol. Bioeng. 2002, 80, 580588. [Google Scholar] [CrossRef]
- Song, H.; O’Connor, K.C.; Lacks, D.J.; Enmon, R.M.; Jain, S.K. Monte Carlo simulation of LNCaP human prostate cancer cell aggregation in liquid-overlay culture. Biotechnol. Prog. 2003, 19, 1742–1749. [Google Scholar] [CrossRef]
- Song, H.; Jain, S.K.; Enmon, R.M.; O’Connor, K.C. Restructuring dynamics of DU 145 and LNCaP prostate cancer spheroids. Vitr. Cell. Dev. Biol. Anim. 2004, 40, 262. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, K.C.; Venczel, M.Z. Predicting aggregation kinetics of DU 145 prostate cancer cells in liquid-overlay culture. Biotechnol. Lett. 2005, 27, 1663–1668. [Google Scholar] [CrossRef]
- Foty, R. A simple hanging drop cell culture protocol for generation of 3D spheroids. J. Vis. Exp. 2011, 2720. [Google Scholar] [CrossRef]
- Eder, T.; Eder, I.E. 3D Hanging Drop Culture to Establish Prostate Cancer Organoids. Methods Mol. Biol. 2017, 1612, 167–175. [Google Scholar] [PubMed]
- Bhatia, S.N.; Ingber, D.E. Microfluidic organs-on-chips. Nat. Biotechnol. 2014, 32, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, A.Y.; Torisawa, Y.-S.; Tung, Y.-C.; Sud, S.; Taichman, R.S.; Pienta, K.J.; Takayama, S. Microfluidic system for formation of PC-3 prostate cancer co-culture spheroids. Biomaterials 2009, 30, 3020–3027. [Google Scholar] [CrossRef] [PubMed]
- Härmä, V.; Virtanen, J.; Mäkelä, R.; Happonen, A.; Mpindi, J.P.; Knuuttila, M.; Kohonen, P.; Lötjönen, J.; Kallioniemi, O.; Nees, M. A comprehensive panel of three-dimensional models for studies of prostate cancer growth, invasion and drug responses. PLoS ONE 2010, 5, e10431. [Google Scholar] [CrossRef]
- Dolega, M.E.; Abeille, F.; Picollet-D’hahan, N.; Gidrol, X. Controlled 3D culture in Matrigel microbeads to analyze clonal acinar development. Biomaterials 2015, 52, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, J.; Nie, Q.; Zhang, Q.; Liu, S.; Ge, D.; You, Z. Organoid culture of human prostate cancer cell lines LNCaP and C4-2B. Am. J. Clin. Exp. Urol. 2017, 5, 25–33. [Google Scholar]
- Haq, S.; Samuel, V.; Haxho, F.; Akasov, R.; Leko, M.; Burov, S.; Markvicheva, E.; Szewczuk, M.R. Sialylation facilitates self-assembly of 3D multicellular prostaspheres by using cyclo-RGDfK(TPP) peptide. Onco Targets Ther. 2017, 10, 2427–2447. [Google Scholar] [CrossRef]
- Meinert, C.; Theodoropoulos, C.; Klein, T.J.; Hutmacher, D.W.; Loessner, D. A method for prostate and breast cancer cell spheroid cultures using gelatin methacryloyl-based hydrogels. Methods Mol. Biol. 2018, 1786, 175–194. [Google Scholar]
- Hainline, K.M.; Gu, F.; Handley, J.F.; Tian, Y.F.; Wu, Y.; de Wet, L.; Vander Griend, D.J.; Collier, J.H. Self-assembling peptide gels for 3d prostate cancer spheroid culture. Macromol. Biosci. 2019, 19, 1800249. [Google Scholar] [CrossRef]
- Katti, K.S.; Molla, M.S.; Karandish, F.; Haldar, M.K.; Mallik, S.; Katti, D.R. Sequential culture on biomimetic nanoclay scaffolds forms three-dimensional tumoroids. J. Biomed. Mater. Res. A 2016, 104, 1591–1602. [Google Scholar] [CrossRef]
- Cecilia, A.; Baecker, A.; Hamann, E.; Rack, A.; van de Kamp, T.; Gruhl, F.J.; Hofmann, R.; Moosmann, J.; Hahn, S.; Kashef, J.; et al. Optimizing structural and mechanical properties of cryogel scaffolds for use in prostate cancer cell culturing. Mater. Sci. Eng. C 2017, 71, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Bäcker, A.; Erhardt, O.; Wietbrock, L.; Schel, N.; Göppert, B.; Dirschka, M.; Abaffy, P.; Sollich, T.; Cecilia, A.; Gruhl, F.J. Silk scaffolds connected with different naturally occurring biomaterials for prostate cancer cell cultivation in 3D. Biopolymers 2017, 107, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Centenera, M.M.; Gillis, J.L.; Hanson, A.R.; Jindal, S.; Taylor, R.A.; Risbridger, G.P.; Sutherland, P.D.; Scher, H.I.; Raj, G.V.; Knudsen, K.E.; et al. Evidence for efficacy of new Hsp90 inhibitors revealed by ex vivo culture of human prostate tumors. Clin. Cancer Res. 2012, 18, 3562–3570. [Google Scholar] [CrossRef] [PubMed]
- Centenera, M.M.; Raj, G.V.; Knudsen, K.E.; Tilley, V.D.; Butler, L.M. Ex vivo culture of human prostate tissue and drug development. Nat. Rev. Urol. 2013, 10, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, E.V.; Centenera, M.M.; Moldovan, M.; Das, R.; Irani, S.; Vincent, A.D.; Chan, H.; Horvath, L.G.; Lynn, D.J.; Daly, R.J.; et al. Identification of novel response and predictive biomarkers to Hsp90 inhibitors through proteomic profiling of patient-derived prostate tumor explants. Mol. Cell. Proteom. 2018, 17, 1470–1486. [Google Scholar] [CrossRef] [PubMed]
- Centenera, M.M.; Hickey, T.H.; Jindal, S.; Ryan, N.K.; Ravindranathan, P.; Mohammed, H.; Robinson, J.L.; Schiewer, M.J.; Ma, S.; Kapur, P.; et al. A patient-derived explant (PDE) model of hormone-dependent cancer. Mol. Oncol. 2018, 12, 1608–1622. [Google Scholar] [CrossRef]
- Shafi, A.A.; Schiewer, M.J.; de Leeuw, R.; Dylgjeri, E.; McCue, P.A.; Shah, N.; GomellA, L.G.; Lallas, C.D.; Trabulsi, E.J.; Centenera, M.M.; et al. Patient-derived models reveal impact of the tumor microenvironment on therapeutic response. Eur. Urol. Oncol. 2018, 1, 325–337. [Google Scholar] [CrossRef]
- Mutuku, S.M.; Trim, P.J.; Prabhala, B.K.; Irani, S.; Bremert, K.L.; Logan, J.M.; Brooks, D.A.; Stahl, J.; Centenera, M.M.; Snel, M.F.; et al. Evaluation of small molecule drug uptake in patient-derived prostate cancer explants by mass spectrometry. Sci. Rep. 2019, 9, 15008. [Google Scholar] [CrossRef]
- Chu, J.H.; Yu, S.; Hayward, S.W.; Chan, F.L. Development of a three-dimensional culture model of prostatic epithelial cells and its use for the study of epithelial-mesenchymal transition and inhibition of PI3K pathway in prostate cancer. Prostate 2009, 69, 428–442. [Google Scholar] [CrossRef]
- Cao, J.; Chiarelli, C.; Richman, O.; Zarrabi, K.; Kozarekar, P.; Zucker, S. Membrane type 1 matrix metalloproteinase induces epithelial-to-mesenchymal transition in prostate cancer. J. Biol. Chem. 2008, 283, 6232–6240. [Google Scholar] [CrossRef]
- Härmä, V.; Knuuttila, M.; Virtanen, J.; Mirtti, T.; Kohonen, P.; Kovanen, P.; Happonen, A.; Kaewphan, S.; Ahonen, I.; Kallioniemi, O.; et al. Lysophosphatidic acid and sphingosine-1-phosphate promote morphogenesis and block invasion of prostate cancer cells in three-dimensional organotypic models. Oncogene 2012, 31, 2075–2089. [Google Scholar] [CrossRef] [PubMed]
- Ilic, D.; Djulbegovic, M.; Jung, J.H.; Hwang, E.C.; Zhou, Q.; Cleves, A.; Agoritsas, T.; Dahm, P. Prostate cancer screening with prostate-specific antigen (PSA) test: A systematic review and meta-analysis. BMJ 2018, 362, k3519. [Google Scholar] [CrossRef] [PubMed]
- Ballangrud, Å.M.; Yang, W.H.; Dnistrian, A.; Lampen, N.M.; Sgouros, G. Growth and characterization of LNCaP prostate cancer cell spheroids. Clin. Cancer Res. 1999, 5, 3171s–3176s. [Google Scholar] [PubMed]
- Lang, S.H.; Sharrard, R.M.; Stark, M.; Villette, J.M.; Maitland, N.J. Prostate epithelial cell lines form spheroids with evidence of glandular differentiation in three-dimensional Matrigel cultures. Br. J. Cancer 2001, 85, 590–599. [Google Scholar] [CrossRef]
- Song, H.; O’Connor, K.C.; David, O.; Giordano, C.L.; Pappas-LeBeau, H.; Clejan, S. Immunohistochemical analysis of differentiation in static and mixed prostate cancer spheroids. J. Cell. Mol. Med. 2003, 7, 180–186. [Google Scholar] [CrossRef]
- Murphy, R.A.; Watson, A.Y.; Rhodes, J.A. Biological sources of nerve growth factor. Appl. Neurophysiol. 1984, 47, 33–42. [Google Scholar] [CrossRef]
- Cunha, G.R.; Ricke, W.; Thomson, A.; Marker, P.C.; Risbridger, G.; Hayward, S.W.; Wang, Y.Z.; Donjacour, A.A.; Kurita, T. Hormonal, cellular, and molecular regulation of normal and neoplastic prostatic development. J. Steroid Biochem. Mol. Biol. 2004, 92, 221–236. [Google Scholar] [CrossRef]
- Di Donato, M.; Cernera, G.; Auricchio, F.; Migliaccio, A.; Castoria, G. Cross-talk between androgen receptor and nerve growth factor receptor in prostate cancer cells: Implications for a new therapeutic approach. Cell Death Discov. 2018, 4, 5. [Google Scholar] [CrossRef]
- Di Donato, M.; Cernera, G.; Migliaccio, A.; Castoria, G. Nerve growth factor induces proliferation and aggressiveness in prostate cancer cells. Cancers 2019, 11, 784. [Google Scholar] [CrossRef]
- Sutherland, R.M. Cell and environment interactions in tumor microregions: The multicell spheroid model. Science 1988, 240, 177–184. [Google Scholar] [CrossRef]
- Takagi, A.; Watanabe, M.; Ishii, Y.; Morita, J.; Hirokawa, Y.; Matsuzaki, T.; Shiraishi, T. Three-dimensional cellular spheroid formation provides human prostate tumor cells with tissue-like features. Anticancer Res. 2007, 27, 45–53. [Google Scholar] [PubMed]
- Kuşoğlu, A.; Biray Avcı, Ç. Cancer stem cells: A brief review of the current status. Gene 2019, 681, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, T.; Ohata, H.; Sato, A.; Yamawaki, K.; Enomoto, T.; Okamoto, K. Tumor-derived spheroids: Relevance to cancer stem cells and clinical applications. Cancer Sci. 2017, 108, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Goksel, G.; Bilir, A.; Uslu, R.; Akbulut, H.; Guven, U.; Oktem, G. WNT1 gene expression alters in heterogeneous population of prostate cancer cells; decreased expression pattern observed in CD133+/CD44+ prostate cancer stem cell spheroids. J. BUON 2014, 19, 207–214. [Google Scholar]
- Oktem, G.; Sercan, O.; Guven, U.; Uslu, R.; Uysal, A.; Goksel, G.; Ayla, S.; Bilir, A. Cancer stem cell differentiation: TGFβ1 and versican may trigger molecules for the organization of tumor spheroids. Oncol. Rep. 2014, 32, 641–649. [Google Scholar] [CrossRef]
- Florczyk, S.J.; Kievit, F.M.; Wang, K.; Erickson, A.E.; Ellenbogen, R.G.; Zhang, M. 3D porous chitosan–alginate scaffolds promote proliferation and enrichment of cancer stem-like cells. J. Mater. Chem. B 2016, 4, 6326–6334. [Google Scholar] [CrossRef]
- Binkhathlan, Z.; Lavasanifar, A. P-glycoprotein inhibition as a therapeutic approach for overcoming multidrug resistance in cancer: Current status and future perspectives. Curr. Cancer Drug Targets 2013, 13, 326–346. [Google Scholar] [CrossRef]
- Wartenberg, M.; Fischer, K.; Hescheler, J.; Sauer, H. Redox regulation of P-glycoprotein-mediated multidrug resistance in multicellular prostate tumor spheroids. Int. J. Cancer 2000, 85, 267–274. [Google Scholar] [CrossRef]
- Wartenberg, M.; Ling, F.C.; Schallenberg, M.; Bäumer, A.T.; Petrat, K.; Hescheler, J.; Sauer, H. Down-regulation of intrinsic P-glycoprotein expression in multicellular prostate tumor spheroids by reactive oxygen species. J. Biol. Chem. 2001, 276, 17420–17428. [Google Scholar] [CrossRef]
- Wartenberg, M.; Fischer, K.; Hescheler, J.; Sauer, H. Modulation of intrinsic P-glycoprotein expression in multicellular prostate tumor spheroids by cell cycle inhibitors. Biochim. Biophys. Acta Mol. Cell Res. 2002, 1589, 49–62. [Google Scholar] [CrossRef]
- Wartenberg, M.; Gronczynska, S.; Bekhite, M.M.; Saric, T.; Niedermeier, W.; Hescheler, J.; Sauer, H. Regulation of the multidrug resistance transporter P-glycoprotein in multicellular prostate tumor spheroids by hyperthermia and reactive oxygen species. Int. J. Cancer 2005, 113, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Wartenberg, M.; Richter, M.; Datchev, A.; Günther, S.; Milosevic, N.; Bekhite, M.M.; Figulla, H.R.; Aran, J.M.; Pétriz, J.; Sauer, H. Glycolytic pyruvate regulates P-glycoprotein expression in multicellular tumor spheroids via modulation of the intracellular redox state. J. Cell. Biochem. 2010, 109, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Wartenberg, M.; Hoffmann, E.; Schwindt, H.; Grünheck, F.; Petros, J.; Arnold, J.R.S.; Hescheler, J.; Sauer, H. Reactive oxygen species-linked regulation of the multidrug resistance transporter P-glycoprotein in Nox-1 overexpressing prostate tumor spheroids. FEBS Lett. 2005, 579, 4541–4549. [Google Scholar] [CrossRef] [PubMed]
- Sauer, H.; Wartenberg, M.; Hescheler, J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell. Physiol. Biochem. 2001, 11, 173–186. [Google Scholar] [CrossRef]
- Wartenberg, M.; Schallenberg, M.; Hescheler, J.; Sauer, H. Reactive oxygen species-mediated regulation of eNOS and iNOS expression in multicellular prostate tumor spheroids. Int. J. Cancer 2003, 104, 274–282. [Google Scholar] [CrossRef]
- Taruno, A. ATP release channels. Int. J. Mol. Sci. 2018, 19, 808. [Google Scholar] [CrossRef]
- Sauer, H.; Diedershagen, H.; Hescheler, J.; Wartenberg, M. Calcium-dependence of hydrogen peroxide-induced c-fos expression and growth stimulation of multicellular prostate tumor spheroids. FEBS Lett. 1997, 419, 201–205. [Google Scholar] [CrossRef]
- Sauer, H.; Hescheler, J.; Reis, D.; Diedershagen, H.; Niedermeier, W.; Wartenberg, M. DC electrical field-induced c-fos expression and growth stimulation in multicellular prostate cancer spheroids. Br. J. Cancer 1997, 75, 1481–1488. [Google Scholar] [CrossRef][Green Version]
- Sauer, H.; Ritgen, J.; Hescheler, J.; Wartenberg, M. Hypotonic Ca2+ signaling and volume regulation in proliferating and quiescent cells from multicellular spheroids. J. Cell. Physiol. 1998, 175, 129–140. [Google Scholar] [CrossRef]
- Wartenberg, M.; Diedershagen, H.; Hescheler, J.; Sauer, H. Growth stimulation versus induction of cell quiescence by hydrogen peroxide in prostate tumor spheroids is encoded by the duration of the Ca2+ response. J. Biol. Chem. 1999, 274, 27759–27767. [Google Scholar] [CrossRef]
- Sauer, H.; Klimm, B.; Hescheler, J.; Wartenberg, M. Activation of p90RSK and growth stimulation of multicellular tumor spheroids are dependent on reactive oxygen species generated after purinergic receptor stimulation by ATP. FASEB J. 2001, 15, 2539–2541. [Google Scholar] [CrossRef] [PubMed]
- Sauer, H.; Stanelle, R.; Hescheler, J.; Wartenberg, M. The DC electrical-field-induced Ca2+ response and growth stimulation of multicellular tumor spheroids are mediated by ATP release and purinergic receptor stimulation. J. Cell Sci. 2002, 115, 3265–3273. [Google Scholar] [PubMed]
- Wang, D.; Dubois, R.N. Eicosanoids and cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.S.Y. Role of Nonsteroidal anti-inflammatory drugs (NSAIDs) in cancer prevention and cancer promotion. Adv. Pharmacol. Sci. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Sha, W.; Olesch, C.; Hanaka, H.; Rådmark, O.; Weigert, A.; Brüne, B. Necrosis in DU145 prostate cancer spheroids induces COX-2/mPGES-1-derived PGE 2 to promote tumor growth and to inhibit T cell activation. Int. J. Cancer 2013, 133, 1578–1588. [Google Scholar] [CrossRef]
- Xu, X.; Gurski, L.A.; Zhang, C.; Harrington, D.A.; Farach-Carson, M.C.; Jia, X. Recreating the tumor microenvironment in a bilayer, hyaluronic acid hydrogel construct for the growth of prostate cancer spheroids. Biomaterials 2012, 33, 9049–9060. [Google Scholar] [CrossRef]
- Taubenberger, A.V.; Bray, L.J.; Haller, B.; Shaposhnykov, A.; Binner, M.; Freudenberg, U.; Guck, J.; Werner, C. 3D extracellular matrix interactions modulate tumour cell growth, invasion and angiogenesis in engineered tumour microenvironments. Acta Biomater. 2016, 36, 73–85. [Google Scholar] [CrossRef]
- Cathomas, R.; Bajory, Z.; Bouzid, M.; El Ghoneimy, A.; Gillessen, S.; Goncalves, F.; Kacso, G.; Kramer, G.; Milecki, P.; Pacik, D.; et al. Management of bone metastases in patients with castration-resistant prostate cancer. Urol. Int. 2014, 92, 377–386. [Google Scholar] [CrossRef]
- Salamanna, F.; Contartese, D.; Maglio, M.; Fini, M. A systematic review on in vitro 3D bone metastases models. A new horizon to recapitulate the native clinical scenario? Oncotarget 2016, 7, 44803–44820. [Google Scholar] [CrossRef]
- Salamanna, F.; Borsari, V.; Brogini, S.; Giavaresi, G.; Parrilli, A.; Cepollaro, S.; Cadossi, M.; Martini, L.; Mazzotti, A.; Fini, M. An in vitro 3D bone metastasis model by using a human bone tissue culture and human sex-related cancer cells. Oncotarget 2016, 7, 76966–76983. [Google Scholar] [CrossRef]
- Sieh, S.; Lubik, A.A.; Clements, J.A.; Nelson, C.C.; Hutmacher, D.W. Interactions between human osteoblasts and prostate cancer cells in a novel 3D in vitro model. Organogenesis 2010, 6, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Herroon, M.K.; Diedrich, J.D.; Podgorski, I. New 3D-culture approaches to study interactions of bone marrow adipocytes with metastatic prostate cancer cells. Front. Endocrinol. 2016, 7, 84. [Google Scholar] [CrossRef]
- Sfanos, K.S.; De Marzo, A.M. Prostate cancer and inflammation: The evidence. Histopathology 2012, 60, 199–215. [Google Scholar] [CrossRef]
- Dang, T.; Liou, G.Y. Macrophage cytokines enhance cell proliferation of normal prostate epithelial cells through activation of ERK and Akt. Sci. Rep. 2018, 8, 7718. [Google Scholar] [CrossRef]
- Sheehan, C.; D’Souza-Schorey, C. Tumor-derived extracellular vesicles: Molecular parcels that enable regulation of the immune response in cancer. J. Cell Sci. 2019, 132, jcs235085. [Google Scholar] [CrossRef]
- Xie, F.; Zhou, X.; Fang, M.; Li, H.; Su, P.; Tu, Y.; Zhang, L.; Zhou, F. Extracellular vesicles in cancer immune microenvironment and cancer immunotherapy. Adv. Sci. 2019, 6, 1901779. [Google Scholar] [CrossRef]
- Sadovska, L.; Zandberga, E.; Sagini, K.; Jēkabsons, K.; Riekstiņa, U.; Kalniņa, Z.; Llorente, A.; Linē, A. A novel 3D heterotypic spheroid model for studying extracellular vesicle-mediated tumour and immune cell communication. Biochem. Biophys. Res. Commun. 2018, 495, 1930–1935. [Google Scholar] [CrossRef]
- Palacios, D.A.; Miyake, M.; Rosser, C.J. Radiosensitization in prostate cancer: Mechanisms and targets. BMC Urol. 2013, 13, 4. [Google Scholar] [CrossRef]
- Camus, V.L.; Stewart, G.; Nailon, W.H.; McLaren, D.B.; Campbell, C.J. Measuring the effects of fractionated radiation therapy in a 3D prostate cancer model system using SERS nanosensors. Analyst 2016, 141, 5056–5061. [Google Scholar] [CrossRef][Green Version]
- Emfietzoglou, D.; Kostarelos, K.; Papakostas, A.; Yang, W.H.; Ballangrud, Å.; Song, H.; Sgouros, G. Liposome-mediated radiotherapeutics within avascular tumor spheroids: Comparative dosimetry study for various radionuclides, liposome systems, and a targeting antibody. J. Nucl. Med. 2005, 46, 89–97. [Google Scholar]
- Rae, C.; Haberkorn, U.; Babich, J.W.; Mairs, R.J. Inhibition of fatty acid synthase sensitizes prostate cancer cells to radiotherapy. Radiat. Res. 2015, 184, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Enmon, R.; Yang, W.H.; Ballangrud, Å.M.; Solit, D.B.; Heller, G.; Rosen, N.; Scher, H.I.; Sgouros, G. Combination treatment with 17-n-allylamino-17-demethoxy geldanamycin and acute irradiation produces supra-additive growth suppression in human prostate carcinoma spheroids. Cancer Res. 2003, 63, 8393–8399. [Google Scholar] [PubMed]
- Crawford, E.D.; Heidenreich, A.; Lawrentschuk, N.; Tombal, B.; Pompeo, A.C.L.; Mendoza-Valdes, A.; Miller, K.; Debruyne, F.M.J.; Klotz, L. Androgen-targeted therapy in men with prostate cancer: Evolving practice and future considerations. Prostate Cancer Prostatic Dis. 2019, 22, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Maughan, B.L.; Antonarakis, E.S. Androgen pathway resistance in prostate cancer and therapeutic implications. Expert Opin. Pharmacother. 2015, 16, 1521–1537. [Google Scholar] [CrossRef] [PubMed]
- Eder, T.; Weber, A.; Neuwirt, H.; Grünbacher, G.; Ploner, C.; Klocker, H.; Sampson, N.; Eder, I. Cancer-associated fibroblasts modify the response of prostate cancer cells to androgen and anti-androgens in three-dimensional spheroid culture. Int. J. Mol. Sci. 2016, 17, 1458. [Google Scholar] [CrossRef]
- Thomas, T.S.; Pachynski, R.K. Treatment of advanced prostate cancer. Mo. Med. 2018, 115, 156–161. [Google Scholar]
- Karandish, F.; Haldar, M.K.; You, S.; Brooks, A.E.; Brooks, B.D.; Guo, B.; Choi, Y.; Mallik, S. Prostate-specific membrane antigen targeted polymersomes for delivering mocetinostat and docetaxel to prostate cancer cell spheroids. ACS Omega 2016, 1, 952–962. [Google Scholar] [CrossRef]
- Du, A.W.; Lu, H.; Stenzel, M.H. Core-cross-linking accelerates antitumor activities of paclitaxel–conjugate micelles to prostate multicellular tumor spheroids: A Comparison of 2D and 3D Models. Biomacromolecules 2015, 16, 1470–1479. [Google Scholar] [CrossRef]
- Petrioli, R.; Fiaschi, A.I.; Francini, E.; Pascucci, A.; Francini, G. The role of doxorubicin and epirubicin in the treatment of patients with metastatic hormone-refractory prostate cancer. Cancer Treat. Rev. 2008, 34, 710–718. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Yasui, W.; Pettaway, C.A.; Yano, S.; Oue, N.; Tahara, E.; Fidler, I.J. Interferon-alpha prevents selection of doxorubicin-resistant undifferentiated-androgen-insensitive metastatic human prostate cancer cells. Prostate 2001, 49, 19–29. [Google Scholar] [CrossRef]
- Wartenberg, M.; Hescheler, J.; Acker, H.; Diedershagen, H.; Sauer, H. Doxorubicin distribution in multicellular prostate cancer spheroids evaluated by confocal laser scanning microscopy and the optical probe technique. Cytometry 1998, 31, 137–145. [Google Scholar] [CrossRef]
- Peng, Z.-H.; Kopeček, J. Enhancing accumulation and penetration of HPMA copolymer–doxorubicin conjugates in 2D and 3D prostate cancer cells via iRGD conjugation with an MMP-2 cleavable spacer. J. Am. Chem. Soc. 2015, 137, 6726–6729. [Google Scholar] [CrossRef]
- Toren, P.; Zoubeidi, A. Targeting the PI3K/Akt pathway in prostate cancer: Challenges and opportunities (Review). Int. J. Oncol. 2014, 45, 1793–1801. [Google Scholar] [CrossRef]
- Jones, D.T.; Valli, A.; Haider, S.; Zhang, Q.; Smethurst, E.A.; Schug, Z.T.; Peck, B.; Aboagye, E.O.; Critchlow, S.E.; Schulze, A.; et al. 3D growth of cancer cells elicits sensitivity to kinase inhibitors but not lipid metabolism modifiers. Mol. Cancer Ther. 2019, 18, 376–388. [Google Scholar] [CrossRef]
- Tee, S.S.; Suster, I.; Truong, S.; Jeong, S.; Eskandari, R.; DiGialleonardo, V.; Alvarez, J.A.; Aldeborgh, H.N.; Keshari, K.R. Targeted AKT inhibition in prostate cancer cells and spheroids reduces aerobic glycolysis and generation of hyperpolarized [1-13C] lactate. Mol. Cancer Res. 2018, 16, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Zhao, Z.Z.; Bo, H.B.; Hao, X.J.; Wang, J.Q. Applications of ruthenium complex in tumor diagnosis and therapy. Front. Pharmacol. 2018, 9, 1323. [Google Scholar] [CrossRef]
- De Grandis, R.A.; dos Santos, P.W.D.S.; de Oliveira, K.M.; Machado, A.R.T.; Aissa, A.F.; Batista, A.A.; Antunes, L.M.G.; Pavan, F.R. Novel lawsone-containing ruthenium(II) complexes: Synthesis, characterization and anticancer activity on 2D and 3D spheroid models of prostate cancer cells. Bioorg. Chem. 2019, 85, 455–468. [Google Scholar] [CrossRef]
- Manasanch, E.E.; Orlowski, R.Z. Proteasome inhibitors in cancer therapy. Nat. Rev. Clin. Oncol. 2017, 5, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.; Man, S.; Elliott, P.; Adams, J.; Kerbel, R.S. Lack of multicellular drug resistance observed in human ovarian and prostate carcinoma treated with the proteasome inhibitor PS-341. Clin. Cancer Res. 2000, 6, 3719–3728. [Google Scholar] [PubMed]
- Kostapanos, M.; Elisaf, M.; Mikhailidis, D. Pioglitazone and cancer: Angel or demon? Curr. Pharm. Des. 2013, 19, 4913–4929. [Google Scholar] [CrossRef]
- Gottfried, E.; Rogenhofer, S.; Waibel, H.; Kunz-Schughart, L.A.; Reichle, A.; Wehrstein, M.; Peuker, A.; Peter, K.; Hartmannsgruber, G.; Andreesen, R.; et al. Pioglitazone modulates tumor cell metabolism and proliferation in multicellular tumor spheroids. Cancer Chemother. Pharmacol. 2011, 67, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Amer, M.H. Gene therapy for cancer: Present status and future perspective. Mol. Cell. Ther. 2014, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Rao, D.D.; Senzer, N.; Nemunaitis, J. RNA interference and cancer therapy. Pharm. Res. 2011, 28, 2983–2995. [Google Scholar] [PubMed]
- Evans, J.C.; Malhotra, M.; Sweeney, K.; Darcy, R.; Nelson, C.C.; Hollier, B.G.; O’Driscoll, C.M. Folate-targeted amphiphilic cyclodextrin nanoparticles incorporating a fusogenic peptide deliver therapeutic siRNA and inhibit the invasive capacity of 3D prostate cancer tumours. Int. J. Pharm. 2017, 532, 511–518. [Google Scholar] [CrossRef]
- Wang, K.; Kievit, F.M.; Florczyk, S.J.; Stephen, Z.R.; Zhang, M. 3D porous chitosan–alginate scaffolds as an in vitro model for evaluating nanoparticle-mediated tumor targeting and gene delivery to prostate cancer. Biomacromolecules 2015, 16, 3362–3372. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Guo, J.; Tierney, E.G.; Curtin, C.M.; Malhotra, M.; Darcy, R.; O’Brien, F.J.; O’Driscoll, C.M. The use of collagen-based scaffolds to simulate prostate cancer bone metastases with potential for evaluating delivery of nanoparticulate gene therapeutics. Biomaterials 2015, 66, 53–66. [Google Scholar] [CrossRef]
- Gerber, H.-P.; Baldwin, M.E.; Shojaei, F. Vascular endothelial growth factor antibodies for anti-angiogenic therapy. In Tumor Angiogenesis; Springer: Berlin/Heidelberg, Germany, 2008; pp. 377–393. [Google Scholar]
- Borgström, P.; Bourdon, M.A.; Hillan, K.J.; Sriramarao, P.; Ferrara, N. Neutralizing anti-vascular endothelial growth factor antibody completely inhibits angiogenesis and growth of human prostate carcinoma micro tumors in vivo. Prostate 1998, 35, 1–10. [Google Scholar] [CrossRef]
- Montagnani Marelli, M.; Marzagalli, M.; Fontana, F.; Raimondi, M.; Moretti, R.M.; Limonta, P. Anticancer properties of tocotrienols: A review of cellular mechanisms and molecular targets. J. Cell. Physiol. 2019, 234, 1147–1164. [Google Scholar]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Moretti, R.M.; Marelli, M.M.; Limonta, P. Tocotrienols and cancer: From the state of the art to promising novel patents. Recent Pat. Anticancer Drug Discov. 2019, 14, 5–18. [Google Scholar]
- Fontana, F.; Moretti, R.M.; Raimondi, M.; Marzagalli, M.; Beretta, G.; Procacci, P.; Sartori, P.; Montagnani Marelli, M.; Limonta, P. δ-Tocotrienol induces apoptosis, involving endoplasmic reticulum stress and autophagy, and paraptosis in prostate cancer cells. Cell Prolif. 2019, 52, e12576. [Google Scholar] [CrossRef]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Di Domizio, A.; Limonta, P. Natural compounds in prostate cancer prevention and treatment: Mechanisms of action and molecular targets. Cells 2020, 9, 460. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Audano, M.; Beretta, G.; Procacci, P.; Sartori, P.; Mitro, N.; Limonta, P. Mitochondrial functional and structural impairment is involved in the antitumor activity of δ-tocotrienol in prostate cancer cells. Free Radic. Biol. Med. 2020, 160, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.; Burgoyne, T.; Lee, A.; Stehno-Bittel, L.; Zaid, G. Arum Palaestinum with isovanillin, linolenic acid and β-sitosterol inhibits prostate cancer spheroids and reduces the growth rate of prostate tumors in mice. BMC Complement. Altern. Med. 2015, 15, 264. [Google Scholar] [CrossRef] [PubMed]
- Chambers, K.F.; Mosaad, E.M.O.; Russell, P.J.; Clements, J.A.; Doran, M.R. 3D cultures of prostate cancer cells cultured in a novel high-throughput culture platform are more resistant to chemotherapeutics compared to cells cultured in monolayer. PLoS ONE 2014, 9, e111029. [Google Scholar] [CrossRef] [PubMed]
- Mittler, F.; Obeïd, P.; Rulina, A.V.; Haguet, V.; Gidrol, X.; Balakirev, M.Y. High-content monitoring of drug effects in a 3d spheroid model. Front. Oncol. 2017, 7, 293. [Google Scholar] [CrossRef]
- Mosaad, E.O.; Chambers, K.F.; Futrega, K.; Clements, J.A.; Doran, M.R. The Microwell-mesh: A high-throughput 3D prostate cancer spheroid and drug-testing platform. Sci. Rep. 2018, 8, 253. [Google Scholar] [CrossRef]
- Mosaad, E.; Chambers, K.; Futrega, K.; Clements, J.; Doran, M.R. Using high throughput microtissue culture to study the difference in prostate cancer cell behavior and drug response in 2D and 3D co-cultures. BMC Cancer 2018, 18, 592. [Google Scholar] [CrossRef]


{kind=link}
| Method | Advantages | Concerns | Ref. |
|---|---|---|---|
| Suspension cell cultures | Simple, low-cost, consistent yield, suitable for multicellular spheroids | Difficult control of spheroid size, lack of extracellular matrix surrogates, not suitable for migration/invasion or cell viability assays | [12,13,14,15,16,17,18,19] |
| Hanging drop | Low-cost, uniform spheroids | Labor-intensive, difficult medium exchange, lack of extracellular matrix surrogates, not suitable for migration/invasion or cell viability assays | [21] |
| Microfluidic devices | Uniform spheroids, control of spheroid size, fast spheroid formation, constant perfusion, uniform distribution of oxygen and nutrients | Specialized equipment and expertise, expensive, labor-intensive | [23] |
| Gel-embedding | Extracellular matrix-mimics, suitable for migration/invasion assays, wide variety of polymers | Undefined composition of natural gels, structural modification over time, labor-intensive | [24,25,26,27,28,29] |
| Scaffolds | High tissue mimics, wide variety of materials with wide variety of properties | Expensive, labor-intensive, possible variability between scaffolds | [30,31,32] |
| Patient-derived explants | High tissue mimics, direct assessment of patients’ therapeutic responses | Reliance on fresh tissue, specialized equipment and expertise, labor-intensive | [33,34,35,36,37,38] |
| Treatment | Model | Outcomes | Ref. |
|---|---|---|---|
| Radiotherapy | PC3 cell spheroids (hanging drop) | Surface-Enhanced Raman Spectroscopy (SERS) can be used to assess treatment response | [89] |
| LNCaP-LN3 cell spheroids (suspension) | Liposomes can deliver radiation to the central region of micrometastatic tumors | [90] | |
| LNCaP, PC3, and CWR22Rv1 cell spheroids (suspension) | C75 and 17-N-allylamino-17-demethoxy geldanamycin (17AAG) can sensitize cancer cells to ionizing radiation | [91,92] | |
| Hormone therapy | LNCaP cell/fibroblast co-culture (hanging drop) | Fibroblasts can confer bicalutamide and enzalutamide resistance to cancer cells | [95] |
| Chemotherapy | LNCaP and PC3 cell spheroids (suspension) | Folic acid-coated polymersomes and micelles containing taxanes can increase drug cytotoxicity by reaching the core of the tumor | [97,98] |
| LNCaP-LN3 cell spheroids (suspension) | Doxorubicin can suppress tumor growth | [100] | |
| DU145 cell spheroids (suspension) | 3D spatial arrangement confers doxorubicin resistance to cancer cells via overexpression of P-glycoprotein | [101] | |
| DU145 cell spheroids (suspension) | Polymer-doxorubicin conjugates can enhance drug penetration in tumor tissues | [102] | |
| Targeted therapies | PC3 cell spheroids (suspension) | PTEN/AKT/mTOR inhibitors can suppress tumor growth | [104,105] |
| LNCaP cell spheroids (Matrigel) | Hyperpolarized magnetic resonance spectroscopy (HP-MRS) can be used to assess treatment response | [105] | |
| Novel and experimental therapies | LNCaP, DU145 and PC3 cell spheroids (suspension); 22Rv1 cell spheroids (microfluidic device) PC3 cell spheroids (Matrigel); LNCaP, PC3, and TRAMP-C2 cell spheroids (scaffold) | Ruthenium complexes, proteasome inhibitors, pioglitazone, anti-VEGF antibodies, and nutraceuticals can suppress tumor growth Cationic and cyclodextrin particles are suitable carriers for gene or RNA interference (RNAi) therapies | [107,109,111,118,124] [114,115,116] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontana, F.; Raimondi, M.; Marzagalli, M.; Sommariva, M.; Gagliano, N.; Limonta, P. Three-Dimensional Cell Cultures as an In Vitro Tool for Prostate Cancer Modeling and Drug Discovery. Int. J. Mol. Sci. 2020, 21, 6806. https://doi.org/10.3390/ijms21186806
Fontana F, Raimondi M, Marzagalli M, Sommariva M, Gagliano N, Limonta P. Three-Dimensional Cell Cultures as an In Vitro Tool for Prostate Cancer Modeling and Drug Discovery. International Journal of Molecular Sciences. 2020; 21(18):6806. https://doi.org/10.3390/ijms21186806
Chicago/Turabian StyleFontana, Fabrizio, Michela Raimondi, Monica Marzagalli, Michele Sommariva, Nicoletta Gagliano, and Patrizia Limonta. 2020. "Three-Dimensional Cell Cultures as an In Vitro Tool for Prostate Cancer Modeling and Drug Discovery" International Journal of Molecular Sciences 21, no. 18: 6806. https://doi.org/10.3390/ijms21186806
APA StyleFontana, F., Raimondi, M., Marzagalli, M., Sommariva, M., Gagliano, N., & Limonta, P. (2020). Three-Dimensional Cell Cultures as an In Vitro Tool for Prostate Cancer Modeling and Drug Discovery. International Journal of Molecular Sciences, 21(18), 6806. https://doi.org/10.3390/ijms21186806