Prognostic Value of the Overexpression of Fatty Acid Metabolism-Related Enzymes in Squamous Cell Carcinoma of the Head and Neck

 ,
,
Abstract
:1. Introduction
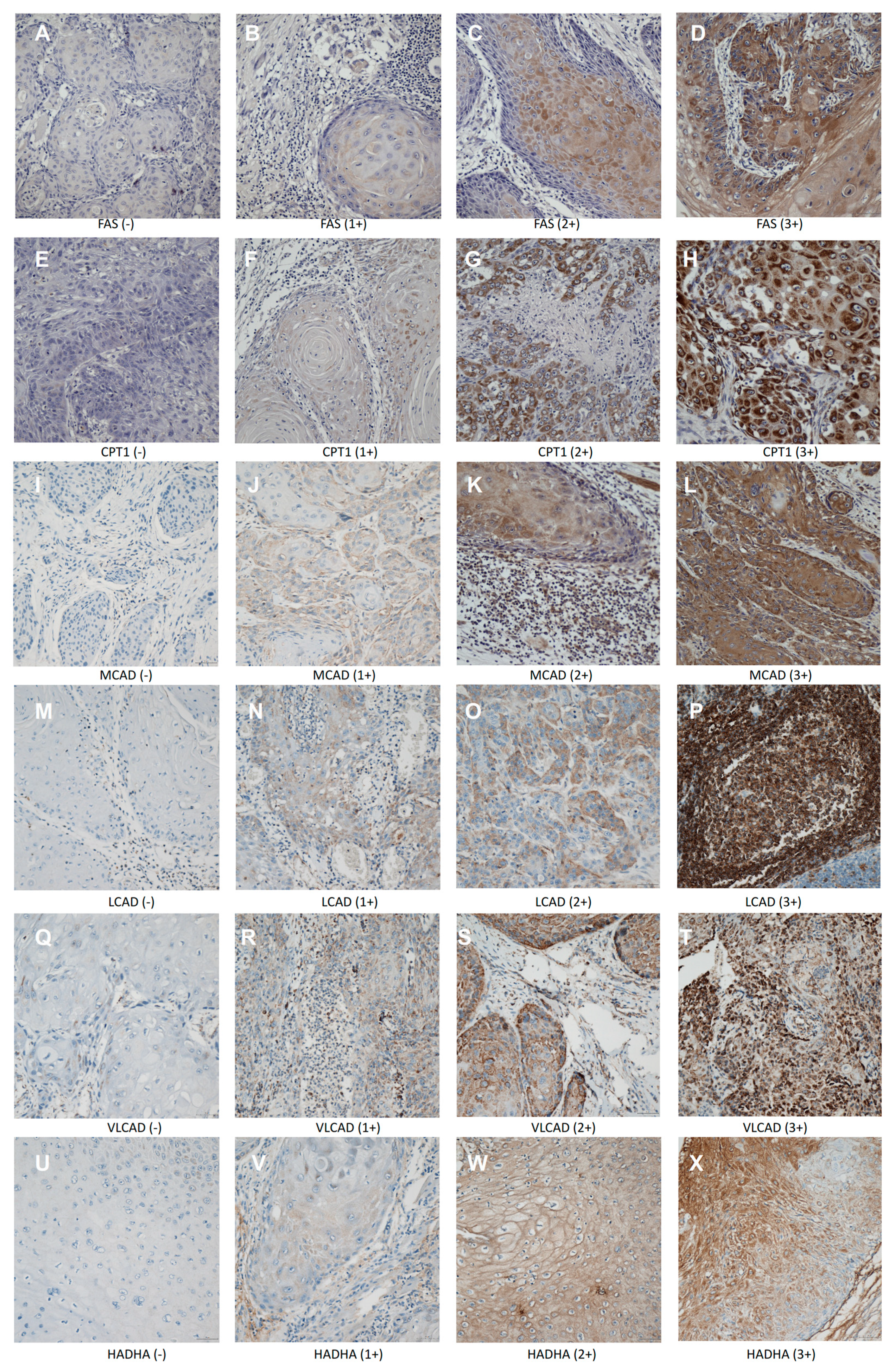
2. Results
2.1. Incidence of Expression of Fao-Related Proteins in Scchn Tumors and Patient Characteristics According to Survival Status, Age, and Clinical Stage
2.2. Univariate and Multivariable Cox Regression Analyses
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Study Subjects and Tissue Microarray Construction
4.3. Immunohistochemistry and Scoring
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACAD | acyl-CoA dehydrogenase |
| ACC | acetyl-CoA carboxylase |
| CI | confidence interval |
| CPT1 | carnitine palmitoyltransferase 1 |
| FAO | fatty acid oxidation |
| FAS | fatty acid synthase |
| HADHA | hydroxyacyl-CoA dehydrogenase/3-ketoacyl-CoA thiolase/enoyl-CoA hydratase |
| HCC | hepatocellular carcinoma |
| HPV | Human Papillomavirus |
| HR | hazard ratio |
| IHC | immunohistochemistry |
| IQR | interquartile range |
| LCAD | long-chain acyl-CoA dehydrogenase |
| MCAD | medium-chain acyl-CoA dehydrogenase |
| OS | overall survival |
| pACC1 | phosphorylated acetyl-CoA carboxylase 1 |
| SCCHN | squamous cell carcinoma of the head and neck |
| TNM | tumor-node-metastasis |
| VLCAD | very-long-chain acyl-CoA dehydrogenase |
| VIF | variance inflation factor |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Gregorio, A.; Martinez-Ramirez, I.; Pedraza-Chaverri, J.; Lizano, M. Reprogramming of energy metabolism in response to radiotherapy in head and neck squamous cell carcinoma. Cancers 2019, 11, 182. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.W.; Lin, Y.H.; Pai, M.H.; Lo, A.C.; Lee, Y.C.; Fang, I.C. Association between phosphorylated AMP-activated protein kinase and acetyl-CoA carboxylase expression and outcome in patients with squamous cell carcinoma of the head and neck. PLoS ONE 2014, 9, e96183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Zhang, C.; Chen, L.; Wang, P.; Fang, Y.; Zhu, J. The role of acetyl-coA carboxylase2 in head and neck squamous cell carcinoma. Peer J. 2019, 7, e7037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronnebaum, S.M.; Joseph, J.W.; Ilkayeva, O.; Burgess, S.C.; Lu, D.; Becker, T.C. Chronic suppression of acetyl-CoA carboxylase 1 in beta-cells impairs insulin secretion via inhibition of glucose rather than lipid metabolism. J. Biol. Chem. 2008, 283, 14248–14256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glatzel, D.K.; Koeberle, A.; Pein, H.; Loser, K.; Stark, A.; Keksel, N. Acetyl-CoA carboxylase 1 regulates endothelial cell migration by shifting the phospholipid composition. J. Lipid Res. 2018, 59, 298–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houten, S.M.; Wanders, R.J.A. A general introduction to the biochemistry of mitochondrial fatty acid β-oxidation. J. Inherit. Metab. Dis. 2010, 33, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Patel, S.; Affleck, V.S.; Wilson, I.; Turnbull, D.M.; Joshi, A.R. Fatty acid oxidation is required for the respiration and proliferation of malignant glioma cells. Neuro Oncol. 2017, 19, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.; Kamarajan, P.; Somashekar, B.S.; MacKinnon, N.; Chinnaiyan, A.M.; Kapila, Y.L. Delineating metabolic signatures of head and neck squamous cell carcinoma: Phospholipase A2, a potential therapeutic target. Int. J. Biochem. Cell Biol. 2012, 44, 1852–1861. [Google Scholar] [CrossRef] [Green Version]
- Sandulache, V.C.; Ow, T.J.; Pickering, C.R.; Frederick, M.J.; Zhou, G.; Fokt, I. Glucose, not glutamine, is the dominant energy source required for proliferation and survival of head and neck squamous carcinoma cells. Cancer 2011, 117, 2926–2938. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.T.; Chen, Y.F.; Lin, S.C.; Chang, K.W.; Li, W.C. Targeting cellular metabolism modulates head and neck oncogenesis. Int. J. Mol. Sci. 2019, 20, 3960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Elheiga, L.; Matzuk, M.M.; Abo-Hashema, K.A.; Wakil, S.J. Continuous fatty acid oxidation and reduced fat storage in mice lacking acetyl-CoA carboxylase 2. Science 2001, 291, 2613–2616. [Google Scholar] [CrossRef]
- Kim, T.S.; Leahy, P.; Freake, H.C. Promoter usage determines tissue specific responsiveness of the rat acetyl-CoA carboxylase gene. Biochem. Biophys. Res. Commun. 1996, 225, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Lea, W.; Abbas, A.S.; Sprecher, H.; Vockley, J.; Schulz, H. Long-chain acyl-CoA dehydrogenase is a key enzyme in the mitochondrial beta-oxidation of unsaturated fatty acids. Biochim. Biophys. Acta 2000, 1485, 121–128. [Google Scholar] [CrossRef]
- He, M.; Pei, Z.; Mohsen, A.; Watkins, P.; Murdoch, G.; Veldhoven, P.P.V.; Ensenauer, R.; Vockley, J. Identification and characterization of new long chain acyl-CoA dehydrogenases. Mol. Genet. Metab. 2011, 102, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Chegary, M.; Brinke, H.; Ruiter, J.P.N.; Wijburg, F.A.; Stoll, M.S.K.; Minkler, P.E. Mitochondrial long chain fatty acid beta-oxidation in man and mouse. Biochim. Biophys. Acta 2009, 1791, 806–815. [Google Scholar] [CrossRef] [Green Version]
- Maher, A.C.; Mohsen, A.W.; Vockley, J.; Tarnopolsky, M.A. Low expression of long-chain acyl-CoA dehydrogenase in human skeletal muscle. Mol. Genet. Metab. 2010, 100, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bharathi, S.S.; Beck, M.E.; Goetzmanet, E.S. The fatty acid oxidation enzyme long-chain acyl-CoA dehydrogenase can be a source of mitochondrial hydrogen peroxide. Redox. Biol. 2019, 26, 101253. [Google Scholar] [CrossRef]
- Huang, D.; Li, T.; Li, X.; Zhang, L.; Sun, L.; He, X. HIF-1-mediated suppression of acyl-CoA dehydrogenases and fatty acid oxidation is critical for cancer progression. Cell Rep. 2014, 8, 1930–1942. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Qin, W.; Jiang, Y.; Yang, Z.; Yuan, B.; Dai, R. ACADL plays a tumor-suppressor role by targeting Hippo/YAP signaling in hepatocellular carcinoma. NPJ Precis Oncol. 2020, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.L.; Li, H.W.; Wang, Y.; Li, C.Q.; You, D.; Jiang, L. Acyl-CoA dehydrogenase long chain expression is associated with esophageal squamous cell carcinoma progression and poor prognosis. Oncol. Targets Ther. 2018, 11, 7643–7653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, V.K.; Ricketts, C.; Bieche, I.; Vacher, S.; Gentle, D.; Lewis, C. Genome-wide DNA methylation profiling of CpG islands in breast cancer identifies novel genes associated with tumorigenicity. Cancer Res. 2011, 71, 2988–2999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 2010, 464, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Liu, R.; Che, J.; Shi, H.; Hou, Y.; Liu, Y. Qualitative and quantitative studies on human B7.1-Fc fusion protein and the application in pharmacokinetic study in rhesus monkeys. J. Pharm. Biomed. Anal. 2011, 54, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.H.; Cheung, C.H.Y.; Liu, Y.L.; Hou, C.L.; Hsu, C.L.; Huang, C.T. Quantitative proteomics of Th-MYCN transgenic mice reveals aurora kinase inhibitor altered metabolic pathways and enhanced ACADM to suppress neuroblastoma progression. J Proteome Res. 2019, 18, 3850–3866. [Google Scholar] [CrossRef]
- Silva, S.D.; Perez, D.E.; Nishimoto, I.N.; Alves, F.A.; Pinto, C.A.L.; Kowalski, L.P. Fatty acid synthase expression in squamous cell carcinoma of the tongue: Clinicopathological findings. Oral. Dis. 2008, 14, 376–382. [Google Scholar] [CrossRef]
- Agostini, M.; Almeida, L.Y.; Bastos, D.C.; Ortega, R.M.; Moreira, F.S.; Seguin, F. The fatty acid synthase inhibitor orlistat reduces the growth and metastasis of orthotopic tongue oral squamous cell carcinomas. Mol. Cancer Ther. 2014, 13, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Aiderus, A.; Black, M.A.; Dunbier, A.K. Fatty acid oxidation is associated with proliferation and prognosis in breast and other cancers. BMC Cancer 2018, 18, 805. [Google Scholar] [CrossRef]
- Hardy, S.; El-Assaad, W.; Przybytkowski, E.; Joly, E.; Prentki, M.; Langelier, Y. Saturated fatty acid-induced apoptosis in MDA-MB-231 breast cancer cells. A role for cardiolipin. J. Biol. Chem. 2003, 278, 31861–31870. [Google Scholar] [CrossRef] [Green Version]
- Padanad, M.S.; Konstantinidou, G.; Venkateswaran, N.; Melegari, M.; Rindhe, S.; Mitsche, M. Fatty acid oxidation mediated by acyl-CoA synthetase long chain 3 is required for mutant KRAS lung tumorigenesis. Cell Rep. 2016, 16, 1614–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lydiatt, W.; O’Sullivan, B.; Patel, S. Major changes in head and neck staging for 2018. Am. Soc. Clin. Oncol. Educ. Book 2018, 38, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Chiang, C.Y.; Liao, K.W.; Ho, S.Y.; Chang, Y.F.; Lim, K.H. cIAP-2 expression increases in elderly patients with squamous cell carcinoma of the head and neck. Int. J. Gerontol. 2017, 11, 22–26. [Google Scholar] [CrossRef]


{kind=link}
| Patient Characteristics | Number (%) |
|---|---|
| Age, median (range) | 51.96 (30.3–86.5) |
| <60 years | 74 (72.5%) |
| >60 years | 28 (27.5%) |
| Gender | |
| Female | 7 (6.9%) |
| Male | 95 (93.1%) |
| Adjuvant treatment | |
| None | 17 (16.7%) |
| Yes | 85 (83.3%) |
| Site | |
| Non-oral cavity | 13 (12.7%) |
| Oral cavity | 89 (87.3%) |
| T | |
| 1 | 13 (12.7%) |
| 2 | 31 (30.4%) |
| 3 | 23 (22.5%) |
| 4 | 35 (34.3%) |
| N | |
| 0 | 49 (48.0%) |
| 1 | 19 (18.6%) |
| 2 | 31 (30.4%) |
| 3 | 3 (2.9%) |
| Stage | |
| I | 8 (7.84%) |
| II | 15 (14.71%) |
| III | 25 (24.51%) |
| IV | 54 (52.94%) |
| HPV P16 | |
| Positive | 13 (12.7%) |
| Negative | 89 (87.3%) |
| Factor | Alive (n = 73) | Dead (n = 29) | p-Value a |
|---|---|---|---|
| n (%) | n (%) | ||
| Age | |||
| median (IQR) | 50.64 (45.13, 57.02) | 56.09 (48.61, 67.05) | 0.026 * |
| Pretreatment fasting sugar | |||
| <100 mg/dL | 42 (57.53) | 15 (51.72) | 0.755 |
| ≥100 mg/dL | 31 (42.47) | 14 (48.28) | |
| median (IQR) | 97.00 (91.00, 105.00) | 96.00 (87.00, 118.00) | 0.807 |
| Pretreatment cholesterol | |||
| <200 mg/dL | 49 (67.12) | 20 (68.97) | 1.000 |
| ≥200 mg/dL | 24 (32.88) | 9 (31.03) | |
| median (IQR) | 183.00 (156.00, 208.00) | 172.00 (150.00, 202.00) | 0.404 |
| Pretreatment triglyceride | |||
| <150 mg/dL | 58 (79.45) | 22 (75.86) | 0.896 |
| ≥150 mg/dL | 15 (20.55) | 7 (24.14) | |
| median (IQR) | 109.00 (84.00, 132.00) | 98.00 (84.00, 129.00) | 0.722 |
| Stage | |||
| I/II | 22 (30.14) | 1 (3.45) | 0.004 * |
| III/IV | 51 (69.86) | 28 (96.55) | |
| T | |||
| 1–2 | 35 (47.95) | 9 (31.03) | 0.120 |
| 3–4 | 38 (52.05) | 20 (68.97) | |
| N | |||
| 0 | 40 (54.79) | 9 (31.03) | 0.032 * |
| 1–3 | 33 (45.21) | 20 (68.97) | |
| FAS | |||
| Negative or low expression (0~1+) | 61 (83.56) | 27 (93.10) | 0.207 |
| Moderate or strong expression (2~3+) | 12 (16.44) | 2 (6.90) | |
| CPT1 | |||
| Negative to moderate expression (0~2+) | 63 (86.30) | 25 (86.21) | 0.990 |
| Strong expression (3+) | 10 (13.70) | 4 (13.79) | |
| MCAD | |||
| Negative to moderate expression (0~2+) | 51 (69.86) | 16 (55.17) | 0.159 |
| Strong expression (3+) | 22 (30.14) | 13 (44.83) | |
| LCAD | |||
| Negative or low expression (0~1+) | 59 (80.82) | 27 (93.10) | 0.124 |
| Moderate or strong expression (2~3+) | 14 (19.18) | 2 (6.90) | |
| VLCAD | |||
| Negative or low expression (0~1+) | 65 (89.04) | 23 (79.31) | 0.198 |
| Moderate or strong expression (2~3+) | 8 (10.96) | 6 (20.69) | |
| HADHA | |||
| Negative to moderate expression (0~2+) | 66 (90.41) | 24 (82.76) | 0.279 |
| Strong expression (3+) | 7 (9.59) | 5 (17.24) |
| Factor | Stage I/II (n = 23) | Stage III/IV (n = 79) | p-Value a |
|---|---|---|---|
| Age median (IQR) | 52.64 (45.14, 63.30) | 51.78 (45.13~61.03) | 0.87 |
| Pretreatment fasting sugar median (IQR) | 92.00 (86.00, 102.00) | 99.00 (88.00, 113.00) | 0.01 * |
| Pretreatment cholesterol median (IQR) | 180.00 (157.00, 201.00) | 183.00 (148.00, 208.00) | 0.95 |
| Pretreatment triglyceride median (IQR) | 109.00 (93.00, 152.00) | 106.00 (83.00, 152.00) | 0.50 |
| FAS (2~3+) n (%) | 2 (8.70%) | 12 (15.19%) | 0.73 |
| CPT1(3+) n (%) | 3 (13.0%) | 11 (13.92%) | 1.00 |
| MCAD (3+) n (%) | 2 (8.70%) | 33 (41.77%) | 0.01 * |
| LCAD (2~3+) n (%) | 1 (4.35%) | 15 (18.99%) | 0.11 |
| VLCAD (2~3+) n (%) | 2 (8.70%) | 12 (15.19%) | 0.73 |
| HADHA (3+) n (%) | 2 (8.70%) | 10 (12.66%) | 1.00 |
| Factor | T1/T2 (n = 44) | T3/T4 (n = 58) | p-Value a |
|---|---|---|---|
| Age median (IQR) | 52.67 (47.87, 63.55) | 51.59 (42.61, 58.43) | 0.15 |
| Pretreatment fasting sugar median (IQR) | 94.00 (87.00, 104.50) | 99.50 (91.00, 116.00) | 0.08 |
| Pretreatment cholesterol median (IQR) | 188.50 (157.00, 206.00) | 174.00 (147.00, 204.00) | 0.30 |
| Pretreatment triglyceride median (IQR) | 110.00 (93.00, 151.50) | 100.50 (81.00, 128.00) | 0.13 |
| FAS (2~3+) n (%) | 2 (4.55%) | 12 (20.69%) | 0.04 * |
| CPT1(3+) n (%) | 3 (6.82%) | 11 (18.97%) | 0.14 |
| MCAD (3+) n (%) | 8 (18.18%) | 27 (46.55%) | 0.01 * |
| LCAD (2~3+) n (%) | 7 (15.91%) | 9 (15.52%) | 1.00 |
| VLCAD (2~3+) n (%) | 4 (9.09%) | 10 (17.24%) | 0.37 |
| HADHA (3+) n (%) | 3 (6.82%) | 9 (15.52%) | 0.30 |
| Factor | Crude HR (95% CI) | p-Value | Adjusted HR (95% CI) | p-Value |
|---|---|---|---|---|
| Age, per 10 years | 1.49 (1.11–1.98) | 0.007 * | 1.75 (1.22–2.49) | 0.002 * |
| Pretreatment fasting sugar | ||||
| ≥100 vs. <100 (mg/dL) | 1.12 (0.54–2.31) | 0.770 | 0.63 (0.29–1.37) | 0.245 |
| Stage: III/IV vs. I/II | 9.59 (1.30–70.51) | 0.026* | 14.33 (1.89–108.60) | 0.010 * |
| FAS: 2~3+ vs. 0~1+ | 0.41 (0.10–1.75) | 0.232 | 0.33 (0.08–1.42) | 0.135 |
| MCAD: 3+ vs. 0~2+ | 1.75 (0.84–3.65) | 0.133 | 1.30 (0.62–2.73) | 0.495 |
| LCAD: 2~3+ vs. 0~1+ | 0.36 (0.09–1.51) | 0.161 | 0.20 (0.05–0.87) | 0.032 * |
| VLCAD: 2~3+ vs. 0~1+ | 1.57 (0.64–3.85) | 0.328 | 1.26 (0.49–3.21) | 0.635 |
| HADHA: 3+ vs. 0~2+ | 1.54 (0.58–4.04) | 0.385 | 0.74 (0.27–2.01) | 0.549 |
| Factor | Crude HR (95% CI) | p-Value | Adjusted HR (95% CI) | p-Value |
|---|---|---|---|---|
| Age, per 10 years | 1.53 (1.10–2.12) | 0.012 * | 1.59 (1.10–2.29) | 0.014 * |
| Pretreatment fasting sugar: ≥100 vs. <100 mg/dL | 0.95 (0.45–1.99) | 0.885 | 0.72 (0.33–1.56) | 0.401 |
| FAS: 2~3+ vs. 0~1+ | 0.17 (0.02–1.21) | 0.077 | 0.15 (0.02–1.13) | 0.066 |
| MCAD: 3+ vs. 0~2+ | 1.34 (0.64–2.82) | 0.439 | 1.36 (0.64–2.90) | 0.431 |
| LCAD: 2~3+ vs. 0~1+ | 0.28 (0.07–1.18) | 0.084 | 0.21 (0.05–0.91) | 0.037 * |
| VLCAD: 2~3+ vs. 0~1+ | 1.46 (0.59–3.60) | 0.415 | 1.41 (0.54–3.64) | 0.483 |
| HADHA: 3+ vs. 0~2+ | 1.44 (0.55–3.80) | 0.460 | 0.80 (0.29–2.20) | 0.668 |
| Factor | Crude HR (95% CI) | p-Value | Adjusted HR (95% CI) | p -Value |
|---|---|---|---|---|
| Age, per 10 years | 1.46 (1.01–2.12) | 0.047 * | 1.92 (1.16–3.18) | 0.012 * |
| Pretreatment fasting sugar: ≥100 vs. <100 mg/dL | 0.67 (0.28–1.61) | 0.369 | 0.41 (0.16–1.07) | 0.069 |
| FAS: 2~3+ vs. 0~1+ | 0.16 (0.02–1.19) | 0.073 | 0.13 (0.02–1.00) | 0.050 |
| MCAD: 3+ vs. 0~2+ | 1.00 (0.42–2.42) | 0.997 | 1.16 (0.46–2.92) | 0.757 |
| LCAD: 2~3+ vs. 0~1+ | 0.24 (0.03–1.78) | 0.162 | 0.09 (0.01–0.85) | 0.035 * |
| VLCAD: 2~3+ vs. 0~1+ | 1.51 (0.55–4.16) | 0.425 | 0.99 (0.32–3.11) | 0.989 |
| HADHA: 3+ vs. 0~2+ | 1.22 (0.41–3.66) | 0.723 | 0.44 (0.13–1.50) | 0.187 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.-W.; Wu, P.-S.; Lin, S.-H.; Huang, W.-Y.; Kuo, Y.-S.; Lin, H.-P. Prognostic Value of the Overexpression of Fatty Acid Metabolism-Related Enzymes in Squamous Cell Carcinoma of the Head and Neck. Int. J. Mol. Sci. 2020, 21, 6851. https://doi.org/10.3390/ijms21186851
Su Y-W, Wu P-S, Lin S-H, Huang W-Y, Kuo Y-S, Lin H-P. Prognostic Value of the Overexpression of Fatty Acid Metabolism-Related Enzymes in Squamous Cell Carcinoma of the Head and Neck. International Journal of Molecular Sciences. 2020; 21(18):6851. https://doi.org/10.3390/ijms21186851
Chicago/Turabian StyleSu, Ying-Wen, Pao-Shu Wu, Sheng-Hsiang Lin, Wen-Yu Huang, Yu-Shao Kuo, and Hung-Pin Lin. 2020. "Prognostic Value of the Overexpression of Fatty Acid Metabolism-Related Enzymes in Squamous Cell Carcinoma of the Head and Neck" International Journal of Molecular Sciences 21, no. 18: 6851. https://doi.org/10.3390/ijms21186851