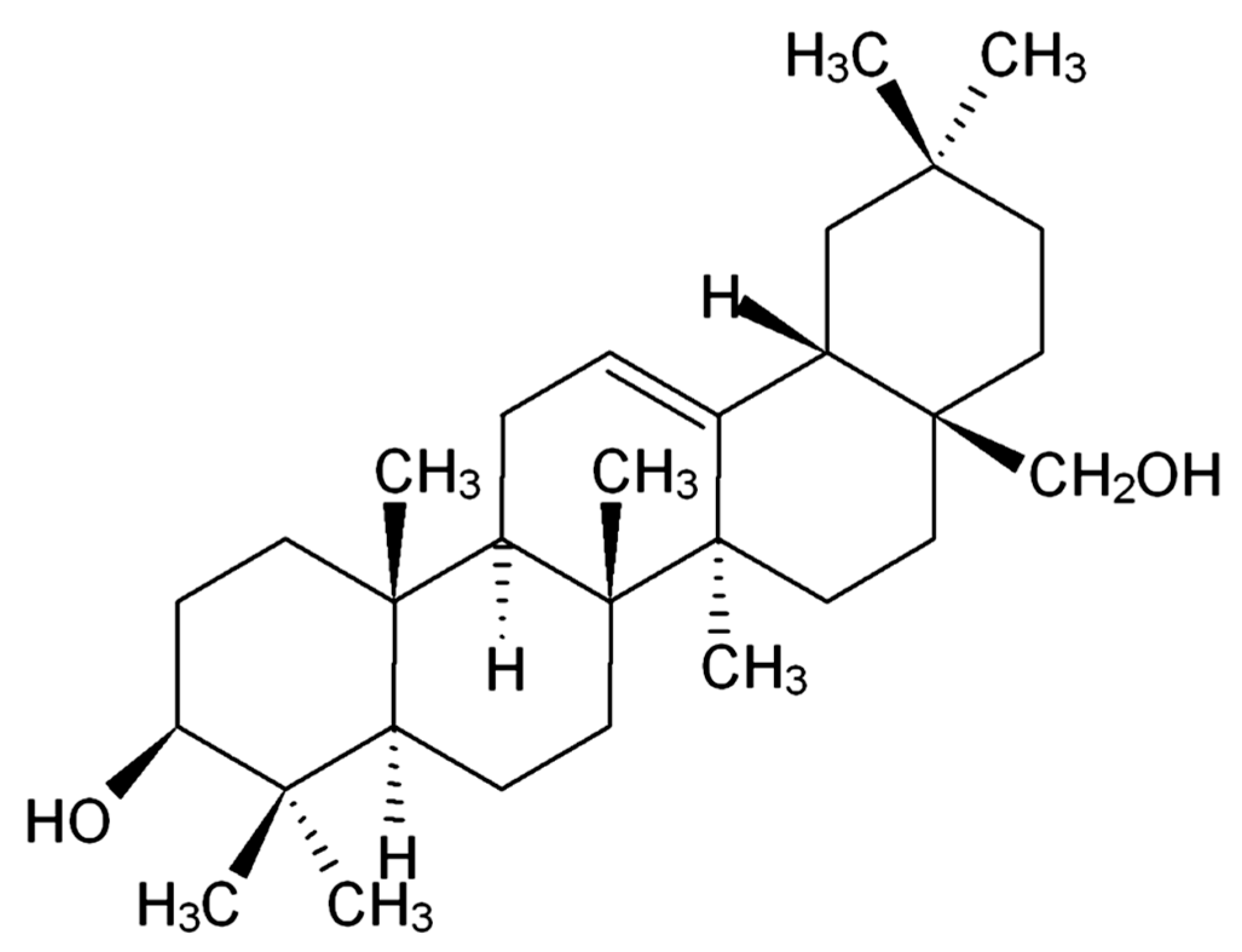
Dietary Erythrodiol Modifies Hepatic Transcriptome in Mice in a Sex and Dose-Dependent Way


 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. Somatometric Parameters
2.2. Histological Analyses
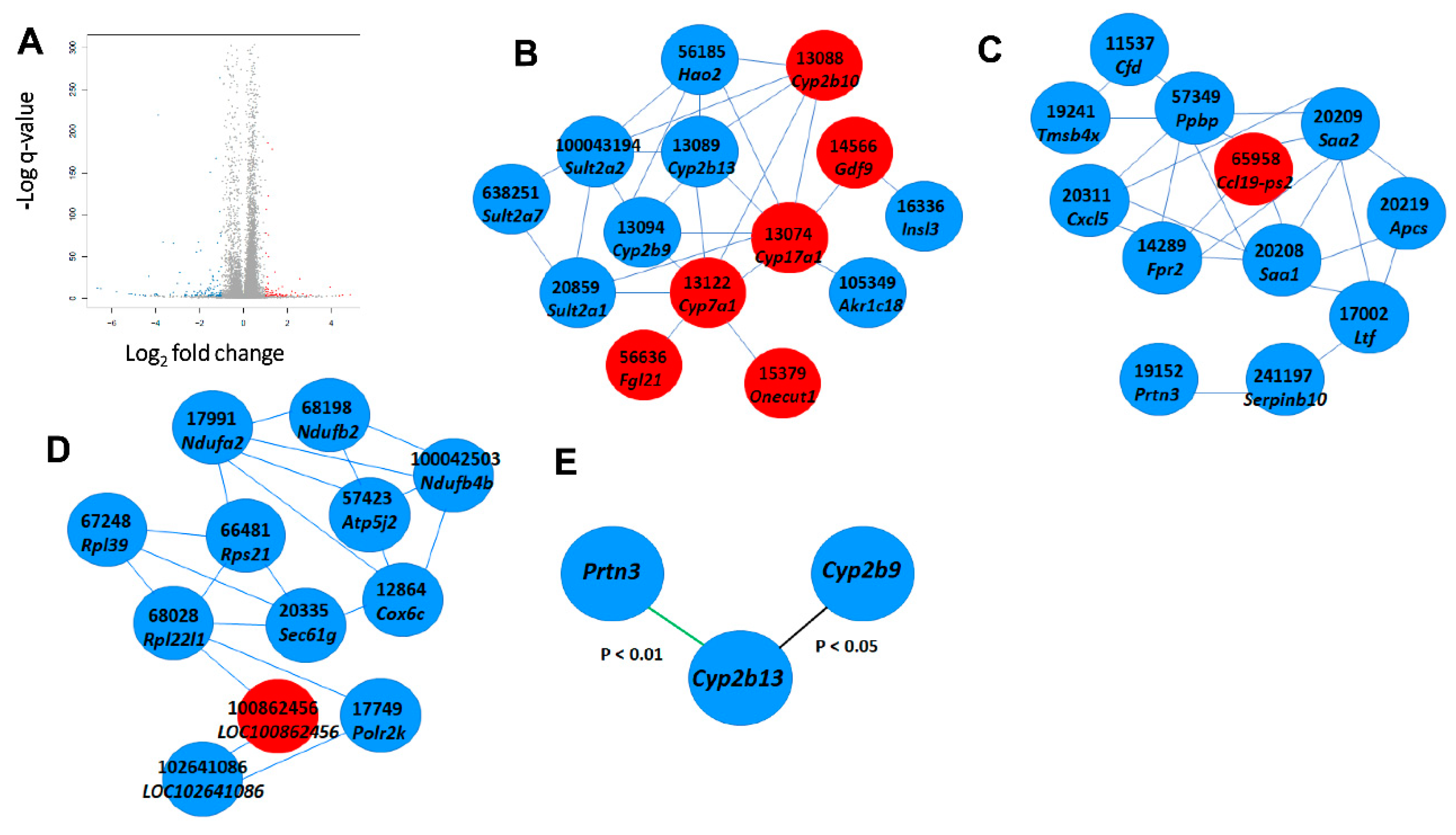
2.3. Hepatic Gene Expression of Apoe-Deficient Male Mice Fed for 12 Weeks on a 10 mg/kg Erythrodiol-Containing Western Diet
2.4. Hepatic Gene Expression in the Livers of Female Apoe-Deficient Mice Fed on a 10 mg/kg Erythrodiol-Containing Western Diet for 12 Weeks
2.5. Influence of Erythrodiol Dose on Selected Hepatic Gene Expressions in Male Apoe-Deficient Mice Fed on Erythrodiol-Containing Western Diets for 12 Weeks
2.6. Influence of Apoa1-Deficiency on Selected Hepatic Gene Expressions on Mice Consuming the 10 mg/kg Erythrodiol-Containing Western Diet for Four Weeks
3. Discussion
4. Materials and Methods
4.1. Animal Models
4.2. Experimental Designs
4.2.1. Effect of Dietary 10 mg/kg Erythrodiol in a Western Diet on Apoe- and Apoa1-Deficient Mice
4.2.2. Effect of Different Doses of Erythrodiol in Western Diets on Male Apoe-Deficient Mice
4.3. Somatometric Analyses
4.4. Liver Histology Analyses
4.5. RNA Isolation
4.6. RNAseq and Data Analyses
4.7. Quantification of mRNA
4.8. Quality Control and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keys, A. Mediterranean diet and public health: Personal reflections. Am. J. Clin. Nutr. 1995, 61, 1321S–1323S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trichopoulou, A.; Costacou, T.; Bamia, C.; Trichopoulos, D. Adherence to a mediterranean diet and survival in a greek population. N. Engl. J. Med. 2003, 348, 2599–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estruch, R.; Ros, E.; Salas-Salvado, J.; Covas, M.I.; Corella, D.; Aros, F.; Gomez-Gracia, E.; Ruiz-Gutierrez, V.; Fiol, M.; Lapetra, J.; et al. Primary prevention of cardiovascular disease with a mediterranean diet supplemented with extra-virgin olive oil or nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, M.A.; Gea, A.; Ruiz-Canela, M. The mediterranean diet and cardiovascular health. Circ. Res. 2019, 124, 779–798. [Google Scholar] [CrossRef] [PubMed]
- Foscolou, A.; Critselis, E.; Panagiotakos, D. Olive oil consumption and human health: A narrative review. Maturitas 2018, 118, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Lou-Bonafonte, J.M.; Arnal, C.; Navarro, M.A.; Osada, J. Efficacy of bioactive compounds from extra virgin olive oil to modulate atherosclerosis development. Mol. Nutr. Food Res. 2012, 56, 1043–1057. [Google Scholar] [CrossRef] [PubMed]
- Habib, L.; Jraij, A.; Khreich, N.; Charcosset, C.; Greige-Gerges, H. Effect of erythrodiol, a natural pentacyclic triterpene from olive oil, on the lipid membrane properties. J. Membr. Biol. 2015, 248, 1079–1087. [Google Scholar] [CrossRef]
- De la Puerta, R.; Martinez-Dominguez, E.; Ruiz-Gutierrez, V. Effect of minor components of virgin olive oil on topical antiinflammatory assays. Zeitschrift für Naturforschung C 2000, 55, 814–819. [Google Scholar] [CrossRef] [Green Version]
- Acín, S.; Navarro, M.A.; Perona, J.S.; Surra, J.C.; Guillen, N.; Arnal, C.; Sarría, A.J.; Arbonés-Mainar, J.M.; Carnicer, R.; Ruiz-Gutiérrez, V.; et al. Microarray analysis of hepatic genes differentially expressed in the presence of the unsaponifiable fraction of olive oil in apolipoprotein e-deficient mice. Br. J. Nutr. 2007, 97, 628–638. [Google Scholar] [CrossRef] [Green Version]
- Juan, M.E.; Wenzel, U.; Daniel, H.; Planas, J.M. Erythrodiol, a natural triterpenoid from olives, has antiproliferative and apoptotic activity in ht-29 human adenocarcinoma cells. Mol. Nutr. Food Res. 2008, 52, 595–599. [Google Scholar] [CrossRef]
- Perona, J.S.; Arcemis, C.; Ruiz-Gutierrez, V.; Catala, A. Effect of dietary high-oleic-acid oils that are rich in antioxidants on microsomal lipid peroxidation in rats. J. Agric. Food Chem. 2005, 53, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Abbass, H.S.; Ragab, E.A.; El-Salam, A.; Mohammed, I.; El-Hela, A.A. Phytochemical and biological investigation of ficus mysorensis cultivated in egypt. J. Pharm. Chem. Biol. Sci. 2015, 3, 396–407. [Google Scholar]
- Abboud, R.; Charcosset, C.; Greige-Gerges, H. Tetra- and penta-cyclic triterpenes interaction with lipid bilayer membrane: A structural comparative study. J. Membr. Biol. 2016, 249, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Marquez-Martin, A.; De La Puerta, R.; Fernandez-Arche, A.; Ruiz-Gutierrez, V.; Yaqoob, P. Modulation of cytokine secretion by pentacyclic triterpenes from olive pomace oil in human mononuclear cells. Cytokine 2006, 36, 211–217. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Tsoumani, M.E.; Kellici, T.F.; Mavromoustakos, T.; Gerothanassis, I.P.; Tselepis, A.D.; Tzakos, A.G. Deconvoluting the dual antiplatelet activity of a plant extract. J. Agric. Food Chem. 2016, 64, 4511–4521. [Google Scholar] [CrossRef]
- Liu, K.; Qin, Y.H.; Yu, J.Y.; Ma, H.; Song, X.L. 3-beta-epsilonrythrodiol isolated from conyza canadensis inhibits mkn45 human gastric cancer cell proliferation by inducing apoptosis, cell cycle arrest, DNA fragmentation, ros generation and reduces tumor weight and volume in mouse xenograft model. Oncol. Rep. 2016, 35, 2328–2338. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.L.; Lin, K.W.; Huang, A.M.; Tu, H.Y.; Wei, B.L.; Hour, T.C.; Yen, M.H.; Pu, Y.S.; Lin, C.N. Terpenoids induce cell cycle arrest and apoptosis from the stems of celastrus kusanoi associated with reactive oxygen species. J. Agric. Food Chem. 2010, 58, 3808–3812. [Google Scholar] [CrossRef]
- Nkengfack, A.E.; Azebaze, A.G.; Waffo, A.K.; Fomum, Z.T.; Meyer, M.; van Heerden, F.R. Cytotoxic isoflavones from erythrina indica. Phytochemistry 2001, 58, 1113–1120. [Google Scholar] [CrossRef]
- Ebeling, S.; Naumann, K.; Pollok, S.; Wardecki, T.; Vidal, Y.S.S.; Nascimento, J.M.; Boerries, M.; Schmidt, G.; Brandner, J.M.; Merfort, I. From a traditional medicinal plant to a rational drug: Understanding the clinically proven wound healing efficacy of birch bark extract. PLoS ONE 2014, 9, e86147. [Google Scholar] [CrossRef] [Green Version]
- Xiaoli, L.; Naili, W.; Sau, W.M.; Chen, A.S.; Xinsheng, Y. Four new isoflavonoids from the stem bark of erythrina variegata. Chem. Pharm. Bull. 2006, 54, 570–573. [Google Scholar] [CrossRef] [Green Version]
- Allouche, Y.; Warleta, F.; Campos, M.; Sanchez-Quesada, C.; Uceda, M.; Beltran, G.; Gaforio, J.J. Antioxidant, antiproliferative, and pro-apoptotic capacities of pentacyclic triterpenes found in the skin of olives on mcf-7 human breast cancer cells and their effects on DNA damage. J. Agric. Food Chem. 2011, 59, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Ibeas, E.; Carvalho-Tavares, J.; Hernandez, M.; Ruiz-Gutierrez, V.; Nieto, M.L. Natural triterpenic diols promote apoptosis in astrocytoma cells through ros-mediated mitochondrial depolarization and jnk activation. PLoS ONE 2009, 4, e5975. [Google Scholar] [CrossRef] [Green Version]
- Allouche, Y.; Beltran, G.; Gaforio, J.J.; Uceda, M.; Mesa, M.D. Antioxidant and antiatherogenic activities of pentacyclic triterpenic diols and acids. Food Chem. Toxicol. 2010, 48, 2885–2890. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Miana, M.; Jurado-Lopez, R.; Martinez-Martinez, E.; Gomez-Hurtado, N.; Delgado, C.; Bartolome, M.V.; San Roman, J.A.; Cordova, C.; Lahera, V.; et al. Diol triterpenes block profibrotic effects of angiotensin ii and protect from cardiac hypertrophy. PLoS ONE 2012, 7, e41545. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.; Hernandez, M.; Cordova, C.; Nieto, M.L. Natural triterpenes modulate immune-inflammatory markers of experimental autoimmune encephalomyelitis: Therapeutic implications for multiple sclerosis. Br. J. Pharm. 2012, 166, 1708–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, A.P.; Qvist, P.; Lazarus, R.; Lescai, F.; Ju, J.; Nyegaard, M.; Mors, O.; Borglum, A.D.; Li, Q.; Christensen, J.H. Experimental validation of methods for differential gene expression analysis and sample pooling in rna-seq. BMC Genom. 2015, 16, 548. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Wood, C.L.; Blalock, E.M.; Chen, K.C.; Landfield, P.W.; Stromberg, A.J. Statistical implications of pooling rna samples for microarray experiments. BMC Bioinform. 2003, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Gabas-Rivera, C.; Martinez-Beamonte, R.; Rios, J.L.; Navarro, M.A.; Surra, J.C.; Arnal, C.; Rodriguez-Yoldi, M.J.; Osada, J. Dietary oleanolic acid mediates circadian clock gene expression in liver independently of diet and animal model but requires apolipoprotein a1. J. Nutr. Biochem. 2013, 24, 2100–2109. [Google Scholar] [CrossRef]
- Guillen, N.; Acin, S.; Surra, J.C.; Arnal, C.; Godino, J.; Garcia-Granados, A.; Muniesa, P.; Ruiz-Gutierrez, V.; Osada, J. Apolipoprotein e determines the hepatic transcriptional profile of dietary maslinic acid in mice. J. Nutr. Biochem. 2009, 20, 882–893. [Google Scholar] [CrossRef]
- Herrera-Marcos, L.V.; Sancho-Knapik, S.; Gabas-Rivera, C.; Barranquero, C.; Gascon, S.; Romanos, E.; Martinez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Arnal, C.; et al. Pgc1a is responsible for the sex differences in hepatic cidec/fsp27beta mrna expression in hepatic steatosis of mice fed a western diet. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E249–E261. [Google Scholar] [CrossRef]
- Zhao, M.; Zhao, H.; Lin, L.; Wang, Y.; Chen, M.; Wu, B. Nuclear receptor co-repressor rip140 regulates diurnal expression of cytochrome p450 2b10 in mouse liver. Xenobiotica 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Koga, T.; Yao, P.L.; Goudarzi, M.; Murray, I.A.; Balandaram, G.; Gonzalez, F.J.; Perdew, G.H.; Fornace, A.J., Jr.; Peters, J.M. Regulation of cytochrome p450 2b10 (cyp2b10) expression in liver by peroxisome proliferator-activated receptor-beta/delta modulation of sp1 promoter occupancy. J. Biol. Chem. 2016, 291, 25255–25263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabás-Rivera, C.; Jurado-Ruiz, E.; Sánchez-Ortiz, A.; Romanos, E.; Martínez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Arnal, C.; Rodríguez-Yoldi, M.J.; Cristina Andrés-Lacueva, C.; et al. Dietary squalene induces cytochromes cyp2b10 and cyp2c55 independently of sex, dose and diet in several mouse models. Mol. Nutr. Food Res. 2020. [Google Scholar] [CrossRef]
- Heintz, M.M.; Kumar, R.; Rutledge, M.M.; Baldwin, W.S. Cyp2b-null male mice are susceptible to diet-induced obesity and perturbations in lipid homeostasis. J. Nutr. Biochem. 2019, 70, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Mirea, A.M.; Stienstra, R.; Kanneganti, T.D.; Tack, C.J.; Chavakis, T.; Toonen, E.J.M.; Joosten, L.A.B. Mice deficient in the il-1beta activation genes prtn3, elane, and casp1 are protected against the development of obesity-induced nafld. Inflammation 2020, 43, 1054–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Ansari, D.; Zhou, Q.; Sasor, A.; Said Hilmersson, K.; Andersson, R. Low p4ha2 and high prtn3 expression predicts poor survival in patients with pancreatic cancer. Scand. J. Gastroenterol. 2019, 54, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jiang, L.; Rui, L. Identification of mup1 as a regulator for glucose and lipid metabolism in mice. J. Biol. Chem. 2009, 284, 11152–11159. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Fang, X.; Tajima, A.; Geng, X.; Ranganathan, S.; Dong, H.; Trucco, M.; Sperling, M.A. Evolution of hepatic steatosis to fibrosis and adenoma formation in liver-specific growth hormone receptor knockout mice. Front. Endocrinol. 2014, 5, 218. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Liu, J.; Liao, Y.; Jin, C.; Zhang, Z.; Zhao, J.; Liu, K.; Huang, H.; Cao, H.; Cheng, Q. Identification of sec61g as a novel prognostic marker for predicting survival and response to therapies in patients with glioblastoma. Med. Sci. Monit. 2019, 25, 3624–3635. [Google Scholar] [CrossRef]
- Chen, J.; Qian, Z.; Li, F.; Li, J.; Lu, Y. Integrative analysis of microarray data to reveal regulation patterns in the pathogenesis of hepatocellular carcinoma. Gut Liver 2017, 11, 112–120. [Google Scholar] [CrossRef]
- Sultana, N.; Rahman, M.; Myti, S.; Islam, J.; Mustafa, M.G.; Nag, K. A novel knowledge-derived data potentizing method revealed unique liver cancer-associated genetic variants. Hum. Genom. 2019, 13, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollenhauer, J.; Wiemann, S.; Scheurlen, W.; Korn, B.; Hayashi, Y.; Wilgenbus, K.K.; Poustka, A. Dmbt1, a new member of the srcr superfamily, on chromosome 10q25.3–26.1 is deleted in malignant brain tumours. Nat. Genet. 1997, 17, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Kwekel, J.C.; Desai, V.G.; Moland, C.L.; Branham, W.S.; Fuscoe, J.C. Age and sex dependent changes in liver gene expression during the life cycle of the rat. BMC Genom. 2010, 11, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Miao, L.; Yao, J.; Feng, C.; Li, C.; Gao, M.; Liu, M.; Gong, L.; Wang, Y.; Qi, X.; et al. Role of multiple micrornas in the sexually dimorphic expression of cyp2b9 in mouse liver. Drug Metab. Dispos. 2013, 41, 1732–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Mota, L.C.; Litoff, E.J.; Rooney, J.P.; Boswell, W.T.; Courter, E.; Henderson, C.M.; Hernandez, J.P.; Corton, J.C.; Moore, D.D.; et al. Compensatory changes in cyp expression in three different toxicology mouse models: Car-null, cyp3a-null, and cyp2b9/10/13-null mice. PLoS ONE 2017, 12, e0174355. [Google Scholar] [CrossRef] [PubMed]
- Jarukamjorn, K.; Sakuma, T.; Nemoto, N. Discriminating activation of cyp2b9 expression in male c57bl/6 mouse liver by beta-estradiol. Biochem. Biophys. Res. Commun. 2000, 279, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kaneko, Y.; Cho, T.; Goto, K.; Otsuka, T.; Yamamoto, S.; Goto, S.; Maruyama, H.; Narita, I. Prolactin upregulates female-predominant p450 gene expressions and downregulates male-predominant gene expressions in mouse liver. Drug Metab. Dispos. 2017, 45, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Quesada, C.; Lopez-Biedma, A.; Warleta, F.; Campos, M.; Beltran, G.; Gaforio, J.J. Bioactive properties of the main triterpenes found in olives, virgin olive oil, and leaves of olea europaea. J. Agric. Food Chem. 2013, 61, 12173–12182. [Google Scholar] [CrossRef]
- Arbonés-Mainar, J.M.; Navarro, M.A.; Acín, S.; Guzmán, M.A.; Arnal, C.; Surra, J.C.; Carnicer, R.; Roche, H.M.; Osada, J. Trans-10, cis-12- and cis-9, trans-11-conjugated linoleic acid isomers selectively modify hdl-apolipoprotein composition in apolipoprotein e knockout mice. J. Nutr. 2006, 136, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Gimenez, E.; Juan, M.E.; Calvo-Melia, S.; Planas, J.M. A sensitive liquid chromatography-mass spectrometry method for the simultaneous determination in plasma of pentacyclic triterpenes of Olea europaea L. Food Chem. 2017, 229, 534–541. [Google Scholar] [CrossRef]
- Acin, S.; Navarro, M.A.; Perona, J.S.; Arbones-Mainar, J.M.; Surra, J.C.; Guzman, M.A.; Carnicer, R.; Arnal, C.; Orman, I.; Segovia, J.C.; et al. Olive oil preparation determines the atherosclerotic protection in apolipoprotein e knockout mice. J. Nutr. Biochem. 2007, 18, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Surra, J.C.; Guillen, N.; Arbones-Mainar, J.M.; Barranquero, C.; Navarro, M.A.; Arnal, C.; Orman, I.; Segovia, J.C.; Osada, J. Sex as a profound modifier of atherosclerotic lesion development in apolipoprotein e-deficient mice with different genetic backgrounds. J. Atheroscler. Thromb. 2010, 17, 712–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Reddick, R.L.; Maeda, N. Lack of apoa-i is not associated with increased susceptibility to atherosclerosis in mice. Arter. Thromb. 1993, 13, 1814–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabas-Rivera, C.; Barranquero, C.; Martinez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Osada, J. Dietary squalene increases high density lipoprotein-cholesterol and paraoxonase 1 and decreases oxidative stress in mice. PLoS ONE 2014, 9, e104224. [Google Scholar] [CrossRef] [PubMed]
- Carnicer, R.; Navarro, M.A.; Arbones-Mainar, J.M.; Arnal, C.; Surra, J.C.; Acin, S.; Sarria, A.; Blanco-Vaca, F.; Maeda, N.; Osada, J. Genetically based hypertension generated through interaction of mild hypoalphalipoproteinemia and mild hyperhomocysteinemia. J. Hypertens. 2007, 25, 1597–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillen, N.; Acin, S.; Navarro, M.A.; Perona, J.S.; Arbones-Mainar, J.M.; Arnal, C.; Sarria, A.J.; Surra, J.C.; Carnicer, R.; Orman, I.; et al. Squalene in a sex-dependent manner modulates atherosclerotic lesion which correlates with hepatic fat content in apoe-knockout male mice. Atherosclerosis 2008, 197, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. Stringtie enables improved reconstruction of a transcriptome from rna-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of rna-seq experiments with tophat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Zhang, Y.; Ye, Z.-Q.; Liu, X.-Q.; Zhao, S.-Q.; Wei, L.; Gao, G. Cpc: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- Shen, S.; Park, J.W.; Lu, Z.-X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. Rmats: Robust and flexible detection of differential alternative splicing from replicate rna-seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. Rsem: Accurate transcript quantification from rna-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Process | GenBank | Name | Gene Symbol | SL2R | p-Value |
|---|---|---|---|---|---|
| Nucleosome assembly | NM_001195421.1 | H4 clustered histone 17 | H4c17/Hist1h4m | 4.8 | 0.0001 |
| Transcription factor | XM_006537451.3 | DNA-directed RNA polymerases I, II, and III subunit RPABC4 | LOC100862456 | 4.5 | 0.001 |
| Immune response | XM_006536385.3 | Chemokine (C-C motif) ligand 19, pseudogene | Ccl19-ps2 | 4.4 | 0.001 |
| Ion transport | NM_011644.3 | Xndc1-transient receptor potential cation channel, subfamily C, member 2 | Xntrpc | 4.0 | 0.000 |
| Cell differentiation, signaling activity | NM_183282.2 | Actin-like 9 | Actl9 | 3.1 | 0.0003 |
| Proteolysis | NM_025350.4 | Pancreatic carboxypeptidase A1 | Cpa1 | 2.9 | 0.001 |
| NA | XM_017321851.2 | Extensin-like isoform X2 | Gm40365 | 2.7 | 0.000 |
| Chromosomal stability | NM_178212.3 | H2A clustered histone 19 | H2ac19/Hist2h2aa2 | 2.6 | 0.000 |
| Aminopeptidase activity | NM_029008.1 | Laeverin | Lvrn | 2.5 | 0.001 |
| G protein-coupled receptor signaling | NM_146337.1 | Olfactory receptor 1396 | Olfr1396 | 2.5 | 0.001 |
| Proteolysis | NM_025583.2 | Chymotrypsinogen B1 | Ctrb1 | 2.4 | 0.000 |
| NA | XM_017313070.1 | Predicted gene/ coiled-coil domain containing 168 | Gm8251/Ccdc168 | 2.2 | 0.000 |
| Ion transmembrane transport | NM_001099298.3 | Sodium channel, voltage-gated, type II, alpha | Scn2a | 2.2 | 0.000 |
| Cell adhesion | NM_001033364.3 | Cadherin-related family member 2 | Cdhr2 | 2.0 | 0.000 |
| Short-term neuronal synaptic plasticity | NM_172737.4 | Shisa family member7 | Shisa7 | 2.0 | 0.0003 |
| Regulation of immune response | NM_178786.4 | Selection and upkeep of intraepithelial T cells 4 | Skint4 | 1.9 | 0.001 |
| Chromatin organization | NM_139218.1 | Developmental pluripotency-associated 3 | Dppa3 | 2.0 | 0.001 |
| Transcription factor | NM_001029933.3 | Zinc finger protein 114 | Zfp114 | 1.9 | 0.001 |
| Ubiquitin-protein transferase activity | NM_027708.1 | F-box protein 24 | Fbxo24 | 1.8 | 0.0002 |
| Protein glycosylation and carbohydrate metabolism | NM_008051.6 | Fucosyltransferase 1 | Fut1 | 1.8 | 0.000 |
| G protein-coupled receptor | NM_001011852.2 | Olfactory receptor 1029 | Olfr1029 | 1.7 | 0.001 |
| NA | NA | Predicted gene, 40600 | Gm40600 | 1.7 | 0.0001 |
| Cell adhesion | NM_178685.5 | Protocadherin 20 | Pcdh20 | 1.7 | 0.000 |
| Metal ion binding | NM_001220499.3 | Ring finger 223 | Rnf223 | 1.7 | 0.001 |
| NA | NM_029608.1 | Family with sequence similarity 209 | Fam209 | 1.6 | 0.0003 |
| P450 pathways | NM_009999.4 | Cytochrome P450, family 2, subfamily b, polypeptide 10 | Cyp2b10 | 1.6 | 0.000 |
| NA | NM_027511.1 | Histidine rich carboxyl terminus 1 | Hrct1 | 1.6 | 0.001 |
| Cell adhesion | NM_033585.2 | Protocadherin gamma subfamily A, 2 | Pcdhga2 | 1.5 | 0.000 |
| Retinoic acid binding | NM_029958.1 | Lipocalin 12 | Lcn12 | 1.5 | 0.0001 |
| Biological Process | GenBank | Name | Gene Symbol | SL2R | p-Value |
|---|---|---|---|---|---|
| Transcription factor | XM_017319408.2 | Zinc finger protein 969 | Zfp969 | −6.7 | 0.0000 |
| Transcription factor | NM_001242944.1 | Zinc finger protein 965 | Zfp965 | −6.5 | 0.0000 |
| Muscle structure | NM_011652.3 | Titin | Ttn | −5.8 | 0.0000 |
| Transcription factor | NM_001290127.1 | RNA binding motif protein 14(Rbma4) and RNA binding motif protein 4 (Rbm4) | Rbm14-rbm4 | −5.2 | 0.0000 |
| Protein transmembrane transporter activity | NM_011343.3 | Translocase Sec61 gamma subunit | Sec61g | −5.1 | 0.0000 |
| Cell differentiation | NM_001081425 | RNA binding motif protein 24 | Rbm24 | −4.9 | 0.0001 |
| NA | NM_029025.3 | Transmembrane protein 81 | Tmem81 | −4.7 | 0.0002 |
| NA | NA | Nuclear body protein SP140-like | LOC105247075 | −4.5 | 0.001 |
| Metal ion binding, nucleic acid binding | NM_053113.2 | Ribonuclease, RNase A family, 2A (liver, eosinophil-derived neurotoxin) | Rnase2a | −4.5 | 0.001 |
| Sulfotransferase activity | NM_009286.2 | Sulfotransferase family 2A, member 2 | Sult2a2 | −4.4 | 0.001 |
| Response to oxidative stress | XM_001478443.6 | Predicted NADH:ubiquinone oxidoreductase subunit B4B | Ndufb4b | −4.3 | 0.0000 |
| Sulfation of steroids and bile acids | NM_001111296.2 | Sulfotransferase family 2A, member 1 | Sult2a1 | −4.0 | 0.0000 |
| Hydrogen peroxide catabolic process and oxygen transport | NM_001278161.1 | Hemoglobin, beta adult major chain | Hbb-b1 | −4.0 | 0.0000 |
| NA | NA | Small nuclear ribonucleoprotein F | Gm13092 | −4.0 | 0.0000 |
| Ion transport | XM_006509537.4 | Predicted solute carrier family 5 (sodium iodide symporter) | Slc5a5 | −3.9 | 0.0002 |
| Cell differentiation and protein transport | NM_001347632.2 | Deleted in malignant brain tumors 1 | Dmbt1 | −3.9 | 0.0000 |
| P450 pathway | NM_007813.2 | Cytochrome P450, family 2, subfamily b, polypeptide 13 | Cyp2b13 | −3.7 | 0.0000 |
| G protein-coupled receptor signaling pathway | NM_010999.3 | Olfactory receptor 56p | Olfr56 | −3.3 | 0.0000 |
| Cation transport | NM_172583.3 | Transmembrane protein 63c | Tmem63c | −3.2 | 0.0000 |
| Regulation of GTPase activity | NM_011178.2 | Proteinase 3 | Prtn3 | −3.2 | 0.0000 |
| Ions and reactive oxygen species responses | NM_134066.3 | Aldo-keto reductase family 1, member C18 | Akr1c18 | −3.0 | 0.0000 |
| NA | NA | Circumsporozoite protein-like | LOC108167857 | −2.7 | 0.0007 |
| Cell adhesion and blood coagulation | NM_001001999.1 | Glycoprotein Ib, beta polypeptide | Gp1bb | −2.7 | 0.0001 |
| NA | NM_001013773.3 | Neurexophilin and PC-esterase domain family, member 5 | Nxpe5 | −2.6 | 0.0005 |
| Signaling pathway | NM_001101656.2 | CD300 molecule like family member D4 | Cd300ld4 | −2.6 | 0.0000 |
| Regulation of transcription | NG_065348.1 | Coiled-coil-helix-coiled-coil-helix domain containing 2, pseudogene on chromosome 4 | Chchd2-ps | −2.6 | 0.0000 |
| Carbohydrate catabolism | NM_001042711.2 | Amylase 2a5 | Amy2a5 | −2.5 | 0.0000 |
| Cell-matrix adhesion | NM_080457.3 | Mucin 4 | Muc4 | −2.3 | 0.0006 |
| Transcription | NM_001346707 | Predicted gene 3055 | Gm3055 | −2.3 | 0.001 |
| G-protein coupled receptor signaling pathway | NM_013564.7 | Insulin-like 3 | Insl3 | −2.2 | 0.0000 |
| Immune response | NM_011280.2 | Tripartite motif-containing 10 | Trim10 | −2.2 | 0.0001 |
| Signaling pathway | NM_010014.3 | Disabled 1 | Dab1 | −2.1 | 0.0000 |
| Cell growth and differentiation | NM_010052.5 | Delta like non-canonical Notch ligand 1 | Dlk1 | −2.1 | 0.0002 |
| Regulation of cell adhesion | NM_001351947.1 | Olfactomedin 4 | Olfm4 | −2.1 | 0.0000 |
| Ion transport | NM_172469.3 | Chloride intracellular channel 6 | Clic6 | −2.1 | 0.0000 |
| Oxidoreductase activity | NM_021509.5 | Monooxygenase, DBH-like 1 | Moxd1 | −2.0 | 0.0000 |
| P450 pathway | NM_010000.2 | Cytochrome P450, family 2, subfamily b, polypeptide 9 | Cyp2b9 | −2.0 | 0.0000 |
| Ion binding | NM_009789.2 | S100 calcium binding protein G | S100g | −2.0 | 0.0000 |
| Immune response | NM_001013832.2 | G protein-coupled receptor 31, D17Leh66b region | Gpr31b | −2.0 | 0.0003 |
| Proteolysis | NM_010810.5 | Matrix metallopeptidase 7 | Mmp7 | −2.0 | 0.0000 |
| Oxidation process | NM_019545.4 | Hydroxyacid oxidase 2 | Hao2 | −1.9 | 0.0000 |
| Visual perception. | NM_021352.3 | Crystallin, beta B3 | Crybb3 | −1.9 | 0.0000 |
| Iron-binding | NM_008522.3 | Lactotransferrin | Ltf | −1.9 | 0.0000 |
| Glycolytic process | NM_001025388.2 | Enolase 1B | Eno1b | −1.9 | 0.0000 |
| Chromatin organization | NM_178187.4 | H2A clustered histone 8 | Hist1h2ae | −1.7 | 0.0000 |
| Signaling pathway and hydrogen peroxide catabolism | NM_001362755.1 | Dual oxidase 2 | Duox2 | −1.7 | 0.0000 |
| mRNA splicing | NM_183024.1 | Ribonucleoprotein, PTB-binding 2 | Raver2 | −1.7 | 0.0000 |
| Cytoplasmic translation | NM_026517.3 | Ribosomal protein L22 like 1 | Rpl22l1 | −1.7 | 0.00000 |
| Keratinization | NM_009264.2 | Small proline-rich protein 1A | Sprr1a | −1.6 | 0.0000 |
| Carbohydrate and MHC class I protein binding | NM_133203.5 | Killer cell lectin-like receptor, subfamily A, member 17 | Klra17 | −1.6 | 0.001 |
| Modulation of synaptic transmission | NM_023716.2 | Tubulin, beta 2B class IIB | Tubb2b | −1.6 | 0.0000 |
| Ion transmembrane transport | NM_146017.3 | Gamma-aminobutyric acid (GABA) A receptor, pi | Gabrp | −1.6 | 0.0002 |
| G protein-coupled receptor activity | NM_001104614.1 | Vomeronasal 2, receptor 3 | Vmn2r3 | −1.6 | 0.0000 |
| Sulfotransferase activity | NM_001184981.2 | Sulfotransferase family 2A, member 7 | Sult2a7 | −1.6 | 0.0000 |
| Interferon response | NM_011579.3 | T cell specific GTPase 1 | Tgtp1 | −1.5 | 0.0000 |
| Transcription factor | NM_001033123.3 | Predicted gene 14288 | Gm14288 | −1.5 | 0.0000 |
| Binding monosaccharides | NM_001134644.1 | Major urinary protein 13 | Mup13 | −1.5 | 0.0000 |
| Regulation of gene expression, protein kinase B signaling | NM_001163011.1 | Major urinary protein 1 | Mup1 | −1.5 | 0.0000 |
| Gene Symbol | Control (n = 14) | Erythrodiol (n = 15) | Fold Change | SL2R |
|---|---|---|---|---|
| H4c17 | 1.1 ± 0.5 | 1.0 ± 0.3 | 0.85 | −0.23 |
| LOC100862456 | 1.0 ± 0.3 | 1.1 ± 0.9 | 1.1 | 0.13 |
| Ccl19-ps2 | 1.0 ± 0.2 | 0.7 ± 0.3 * | 0.67 | −0.58 |
| Ctrb1 | 2.5 ± 3.4 | 100 ± 341 | 40 | 5.32 |
| Cyp2b10 | 1.5 ± 1.4 | 2.3 ± 5.4 * | 1.46 | 0.55 |
| Zfp969 | 1.9 ± 2.0 | 2.5 ± 1.3 | 1.34 | 0.42 |
| Zfp965 | 1.2 ± 0.6 | 1.5 ± 0.7 | 1.55 | 0.63 |
| Ttn | 1.3 ± 0.8 | 1.1 ± 0.7 | 0.90 | −0.16 |
| Rbm14-rbm4 | 1.0 ± 0.2 | 0.8 ± 0.3 * | 0.77 | −0.37 |
| Sec61g | 4.6 ± 9.4 | 0.1 ± 0.01 * | 0.01 | −6.54 |
| Rbm24 | 1.9 ± 3.4 | 0.8 ± 0.7 | 0.44 | −1.19 |
| Tmem81 | 1.1 ± 0.5 | 0.6 ± 0.2 * | 0.49 | −1.02 |
| Rnase2a | 1.8 ± 2.2 | 1.4 ± 1.5 | 0.79 | −0.34 |
| Sult2a2 | 4.6 ± 13 | 2.1 ± 3.9 | 0.45 | −1.15 |
| Ndufb4b | 1.1 ± 0.6 | 1.1 ± 0.5 | 0.97 | −0.04 |
| Dmbt1 | 1.4 ± 2.0 | 0.01 ± 0.01 | 0.01 | −6.81 |
| Cyp2b13 | 46 ± 105 | 6.2 ± 19 | 0.14 | −2.89 |
| Prtn3 | 7.4 ± 17 | 0.3 ± 0.2 * | 0.04 | −4.79 |
| Amy2a5 | 1.1 ± 0.4 | 0.4 ± 0.1 * | 0.41 | −1.28 |
| Cyp2b9 | 21 ± 31 | 1.0 ± 1.9 * | 0.05 | −4.36 |
| Mup1 | 1.4 ± 0.8 | 0.6 ± 0.5 * | 0.44 | −1.19 |
| Gene Symbol | Control (n = 12) | Erythrodiol (n = 13) |
|---|---|---|
| Cyp2b10 | 1.6 ± 1.4 | 1.4 ± 1.5 |
| Dmbt1 | 1.9 ± 2.8 | 6.5 ± 9.6 |
| Cyp2b13 | 19 ± 18 | 17 ± 17 |
| Prtn3 | 1.4 ± 1.4 | 1.5 ± 1.0 |
| Cyp2b9 | 5.7 ± 4.1 | 5.2 ± 4.3 |
| Gene Symbol | Control (n = 17) | 0.5 mg/kg Erythrodiol (n = 16) | 1 mg/kg Erythrodiol (n = 17) | 5 mg/kg Erythrodiol (n = 17) |
|---|---|---|---|---|
| Cyp2b10 | 1.2 ± 0.7 | 11.1 ± 41.3 | 1.6 ± 1.1 | 1.7 ± 1.8 |
| Dmbt1 | 4.5 ± 15 | 2.5 ± 4.3 | 8.5 ± 27.0 | 1.7 ± 2.5 |
| Amy2a5 | 1.0 ± 0.2 | 1.3 ± 0.2 | 1.1 ± 0.2 | 1.1 ± 0.4 |
| Prtn3 | 1.9 ± 2.9 | 1.1 ± 1.2 | 3.2 ± 7.8 * | 1.2 ± 1.0 |
| Cyp2b9 | 1.6 ± 1.3 | 3.0 ± 2.6 * | 2.2 ± 1.9 | 2.1 ± 1.7 |
| Gene Symbol | Males | Females | ||
|---|---|---|---|---|
| Control (n = 14) | Erythrodiol (n = 15) | Control (n = 9) | Erythrodiol (n = 9) | |
| Cyp2b10 | 30 ± 110 | 1.2 ± 1.8 | 7.2 ± 19 | 0.9 ± 0.9 |
| Dmbt1 | 5.0 ± 10.8 | 7.3 ± 19 | 9.8 ± 21 | 87 ± 156 |
| Cyp2b13 | 4.2 ± 6.3 | 4.5 ± 9.8 | 1.3 ± 0.9 | 1.9 ± 1.1 |
| Prtn3 | 1.2 ± 0.8 | 1.1 ± 0.8 | 1.2 ± 0.9 | 5.3 ± 8.1 |
| Cyp2b9 | 2.4 ± 3.8 | 1.6 ± 1.9 | 1.1 ± 0.5 | 1.2 ± 0.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abuobeid, R.; Herrera-Marcos, L.; Navarro, M.A.; Arnal, C.; Martínez-Beamonte, R.; Surra, J.; Osada, J. Dietary Erythrodiol Modifies Hepatic Transcriptome in Mice in a Sex and Dose-Dependent Way. Int. J. Mol. Sci. 2020, 21, 7331. https://doi.org/10.3390/ijms21197331
Abuobeid R, Herrera-Marcos L, Navarro MA, Arnal C, Martínez-Beamonte R, Surra J, Osada J. Dietary Erythrodiol Modifies Hepatic Transcriptome in Mice in a Sex and Dose-Dependent Way. International Journal of Molecular Sciences. 2020; 21(19):7331. https://doi.org/10.3390/ijms21197331
Chicago/Turabian StyleAbuobeid, Roubi, Luis Herrera-Marcos, María A. Navarro, Carmen Arnal, Roberto Martínez-Beamonte, Joaquín Surra, and Jesús Osada. 2020. "Dietary Erythrodiol Modifies Hepatic Transcriptome in Mice in a Sex and Dose-Dependent Way" International Journal of Molecular Sciences 21, no. 19: 7331. https://doi.org/10.3390/ijms21197331