1. Introduction
Plaque angiogenesis and plaque hemorrhage are major players in the destabilization and rupture of atherosclerotic lesions [
1]. Plaque microvessels increase in numbers via angiogenesis during vulnerable stages of the disease, and microvessels density has been associated with the onset of rupture and clinical manifestations [
2,
3]. Plaque angiogenesis arises from reduced oxygen availability in the plaque, caused by lesion growth and presence of metabolic active inflammatory cells. Triggered by hypoxia, endothelial cells proliferate and migrate from the vasa vasorum to form microvessels to overcome the oxygen demand in the lesion [
1]. However, these plaque microvessels are characterized by poor pericyte coverage, lack of cell junctions, and are highly susceptible to leakage of erythrocytes, leucocytes, and plasma lipids, together described as intraplaque hemorrhage [
4,
5].
Healthy microvessels are present in most organs and tissues, have a well-organized architecture, act as a protection barrier, and provide nutrients and oxygen by passive diffusion to their surroundings. The type of endothelial lining determines the permeability of the vessel. While in continuous microvessels (abundantly found in the ear skin) the endothelial cell lining is uninterrupted, fenestrated microvessels (found in secreting glands) have more pores to increase molecular diffusion of molecules (up to 66 kDa) without compromising their barrier function [
6,
7]. Contrarily, microvessels in atherosclerotic lesions in their pathological and immature state are thought to have a compromised barrier function and are as such more permeable [
4,
8,
9,
10,
11].
To date, clinical available imaging techniques to study plaque angiogenesis and subsequent intraplaque hemorrhage, such as PET [
12], CT [
13], and MRI [
14], do not have sufficient spatial resolution to visualize cellular events or image the detailed microvessels network in small size animal models. Two-photon intravital microscopy (2P-IVM), due to its high resolution, can overcome these limitations, allowing detailed 3D reconstructions of plaque angiogenesis and real-time evaluation of target dynamic processes, such as in vivo hemorrhage [
7,
15,
16].
We hypothesized that permeability of healthy continuous and fenestrated microvessels in living mice can be compared to plaque microvessel permeability to assess pathological leakiness. Permeability can be evaluated by quantification of extravasation of 40 kDa-size dextrans [
7,
17]. It should leak from all vessels [
7,
17], but is expected to leak more from fenestrated vessels than continuous, and more from pathological vessels than healthy vessels. Moreover, it, is known that capillaries with poor pericytes coverage in context of inflammation can be permeable to large dextrans up to 2000 kDa [
17]. Therefore, we also hypothesized that evaluation of 2000 kDa dextran extravasation in the same experimental setting, might be relevant to further assess the pathological permeability.
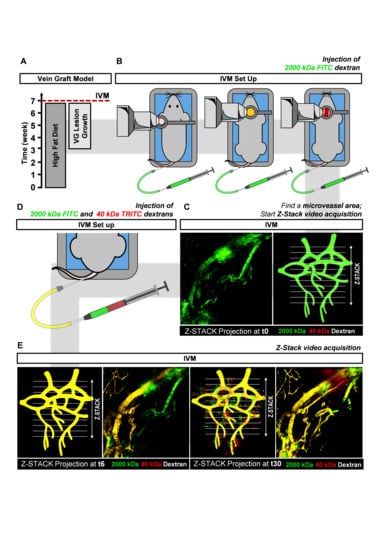
In this study, we set up an advanced 2P-IVM method to visualize atherosclerotic vein graft (VG) lesions in the neck region of living anesthetized mice. (
Figure 1B). With this technique the architecture of adventitial and intimal plaque microvessels in advanced atherosclerotic VG lesions can be visualized (
Figure 1C). This model was chosen since in mice, vasa vasorum angiogenesis only occurs at a very old age and most spontaneous atherosclerotic lesions in mice do not show intraplaque angiogenesis [
11]. We previously showed that hypercholesterolemic ApoE3*Leiden VG lesions, due to their lesion size and state of hypoxia, present with vasa vasorum derived neovascularization, [
8] with the unique characteristic of leaky intimal microvessels and intraplaque hemorrhage [
9]. Furthermore, we report a 2P-IVM method to assess vessel permeability by quantification of 40 kDa and 2000 kDa-labeled dextrans extravasation in healthy microvessels and plaque microvessels in real-time (
Figure 1D,E). We show that plaque microvessels are pathological more permeable in comparison with healthy continuous and fenestrated microvessels, which advocates for their destabilizing role in plaque rupture.
3. Discussion
In this study, we used 2P-IVM to visualize adventitial and intimal plaque microvessels in advanced atherosclerotic lesions in ApoE3*Leiden VGs. In this model, a non-diseased caval vein from a donor mouse is used as an interposition in the carotid artery of a hypercholesterolemic ApoE3*Leiden mouse, within 28 day an atherosclerotic lesion forms with adventitial and intimal plaque microvessels with various forms of maturity [
8,
9]. We report a 2P-IVM method to evaluate plaque microvessels leakiness in vivo, by comparing the extravasation of 40 kDa dextran in healthy, continuous, and fenestrated, as well as diseased, plaque microvessels. We demonstrated in real-time, that microvessels in advanced atherosclerotic lesions in VGs are pathologically permeable.
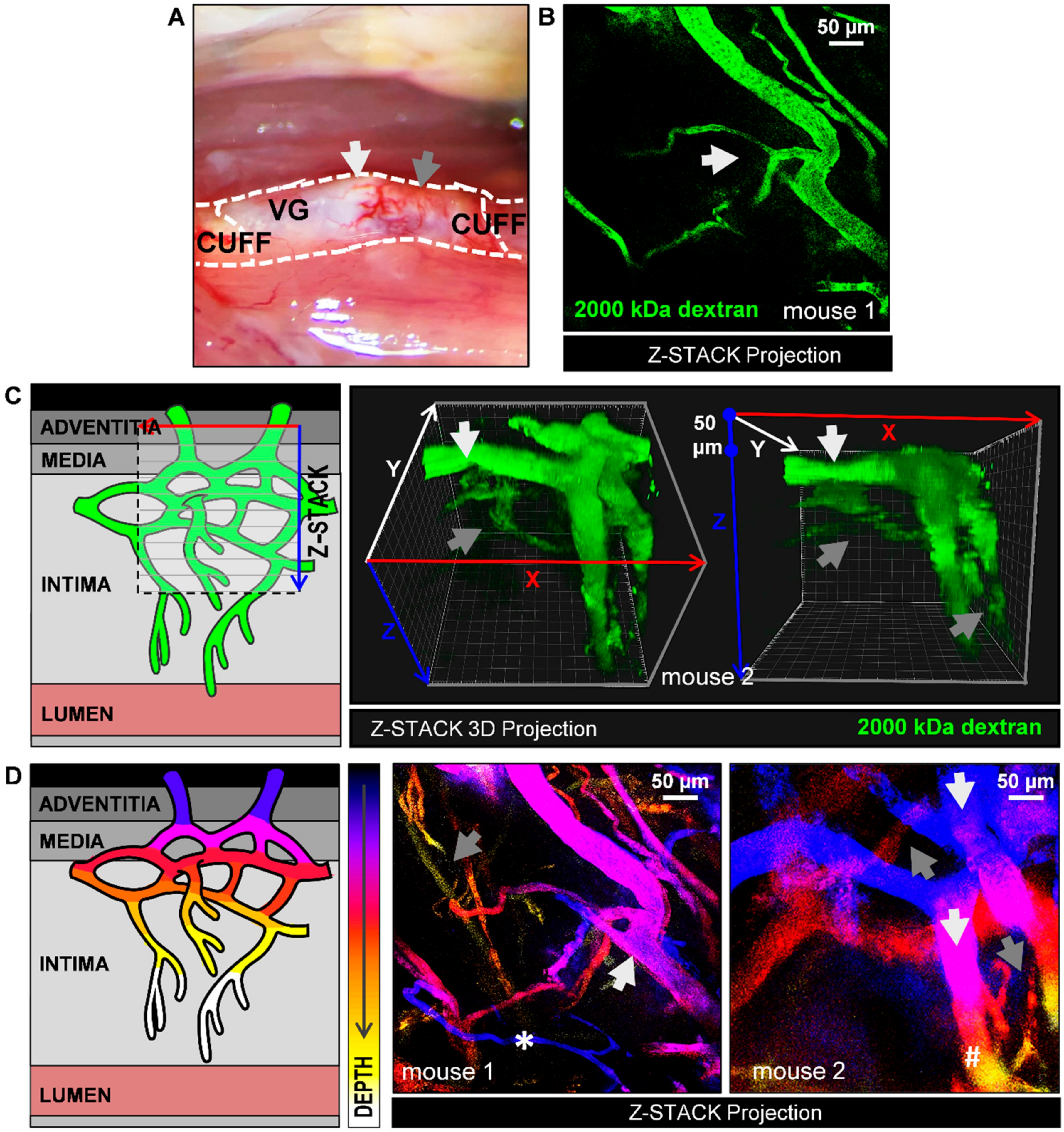
By injecting 2000 kDa-FITC dextrans, we were able to observe in vivo microvessels networks throughout the adventitia, media and extending into the intimal layer of the VG lesion. Larger vessels were detected in the adventitia layer, while more narrow vessel structures were detected further deep in the plaque. We here confirm in vivo the abundant presence of intimal microvessels in advance atherosclerotic lesions in the ApoE3*Leiden VG model that was previously shown with histology [
8,
9]. This is a rare feature seldomly seen in other atherosclerotic mouse models [
19,
20]. These microvessel networks are characterized by typical vessel features of ongoing angiogenesis, as observed by the irregular microvessel architecture with accentuated turns and branching. Moreover, using 3D and color-depth Z-Stack projections, we demonstrate how vasa vasorum angiogenesis evolves throughout the lesion including the intima.
Fenestrated microvessels (observed in salivary glands) are more permeable than continuous microvessels (located in the ears) due to their increased number of pores, which drives faster molecule diffusion [
6,
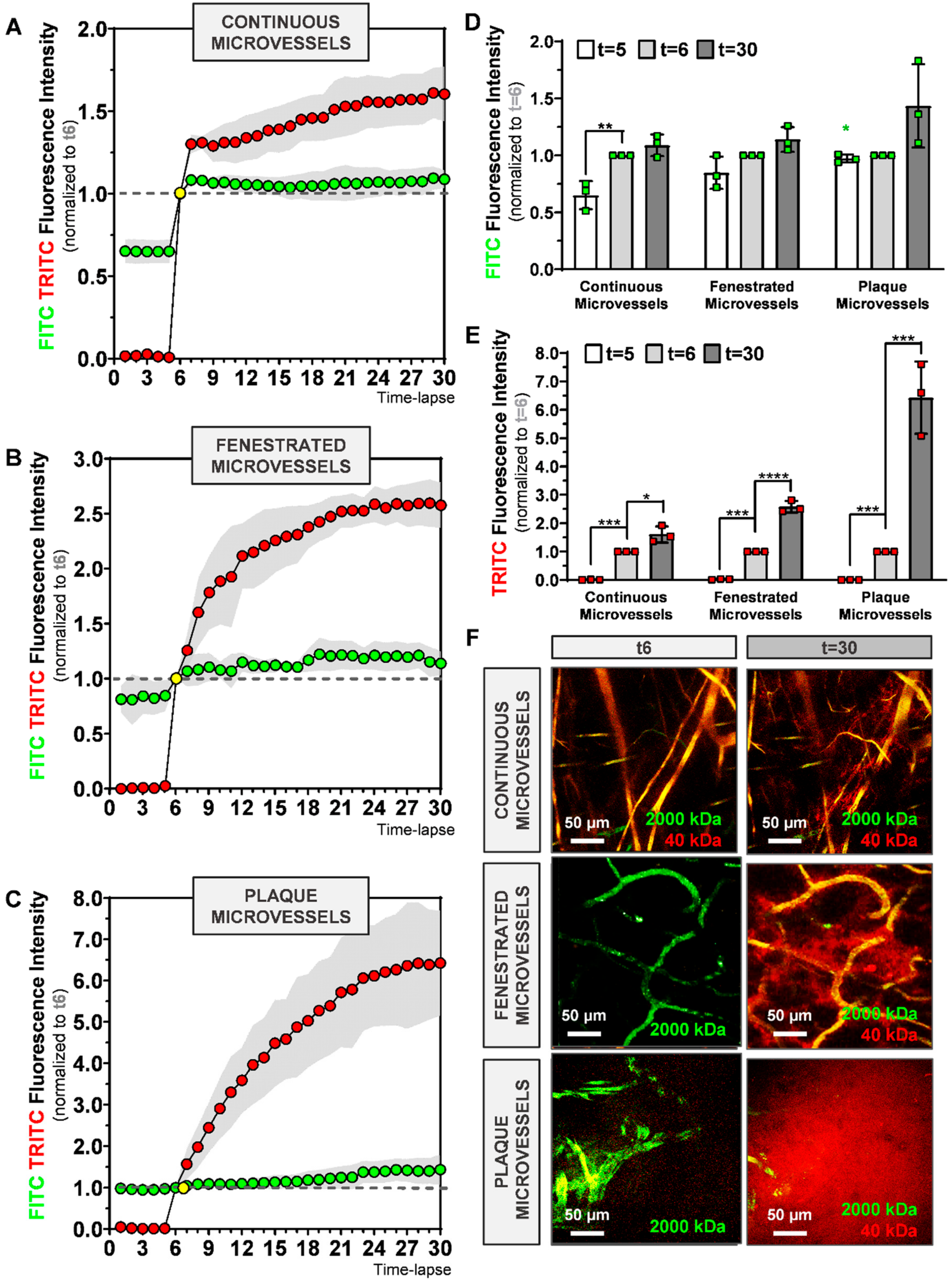
7]. Accordingly, we observed that continuous and fenestrated microvessels follow different 40 kDa dextran extravasation signatures. While in continuous microvessels, extravasation of 40 kDa dextran reached a stable value rapidly in the observation period, extravasation of the 40 kDa dextran in fenestrated microvessels took more time and was more extensive.
Healthy microvessels have a well-organized architecture that acts as a protection barrier but which does let molecules such as nutrients cross. Plaque microvessels, in contrast, have a disorganized structure with a lack of proper pericyte coverage, diminished VE-cadherins junctions, heterogeneous basement membrane and show an unbalance in angiopoietin 1 and 2 expression [
9]. This unbalanced architecture leads to dysfunctionality as shown by their co-localization with extravasated erythrocytes and inflammatory cells, in part explained by the increased expression of VCAM-1 and ICAM-1, as previously described by histological analysis [
8,
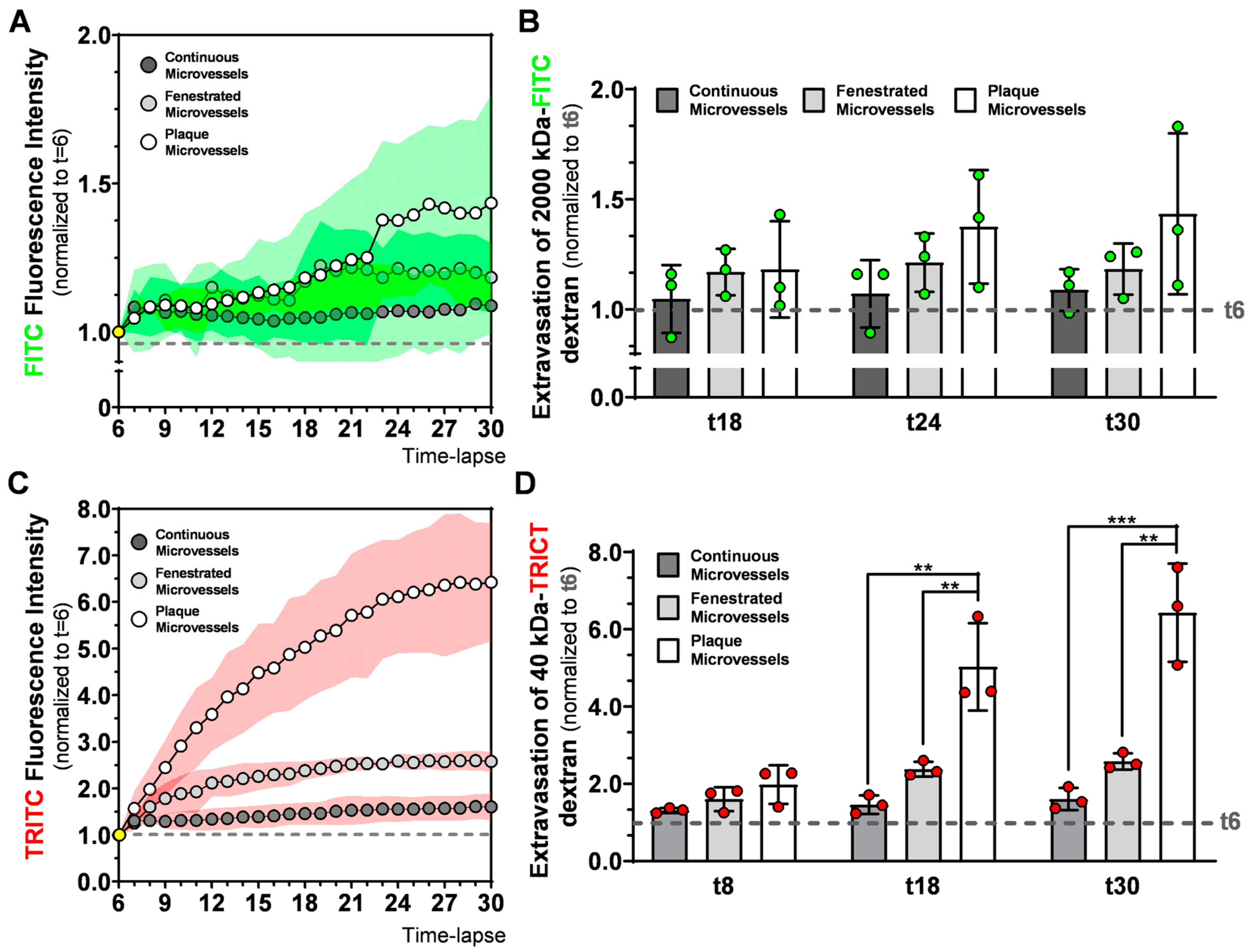
9]. By comparing 40 kDa extravasation patterns in healthy microvessels with the plaque microvessels, we demonstrate in real-time, that microvessels in advanced atherosclerotic lesions are pathological permeable. Their 40 kDa-FITC extravasation curves of the plaque microvessels were clearly different and at the end of the observation period, extravascular 40 kDa dextran was four-fold higher compared to continuous microvessels and 2.5-fold higher compared to fenestrated microvessels. This increased permeability can contribute to the extravasation of erythrocytes and leukocytes which drive plaque instability [
4].
In the 2000 kDa dextran extravasation patterns in continuous, fenestrated, and plaque microvessels FITC extravascular RFI between t6 and t30 showed a trend towards an increase in the plaque microvessels. FITC extravascular RFI at t30 was similar between the groups. The reason why we were not able to detect different FITC extravascular RFI at t30 might be related to the immature nature of the plaque microvessels. Recent studies have shown that microvessels with an immature structure drive macromolecules accumulation by the enhanced permeability and retention (EPR) effect [
21,
22].
In this study, we used two injections of 2000 kDa dextran with the same fluorescent dye (FITC), with the first to visualize blood flow and the second to quantify vessel permeability. After the second injection, FITC intravascular RFI increased significantly between t5 and t6, in all groups. However, FITC extravascular RFI between t5 and t6, only increased significantly in continuous microvessels and did not differ in plaque microvessels. Moreover, when we compared FITC extravascular residual fluorescence (from the first injection) at t5 between groups, it was significantly higher in plaque microvessels compared to continuous microvessels.
Therefore, it is possible that 2000 kDa dextran (from the first injection) accumulates in the extravascular space of plaque microvessels due to the EPR effect, thereby decreasing signal differences during quantification of 2000 kDa extravasation in the second injection. In future approaches that aim to quantify extravasation patterns of macromolecules (such as 2000 kDa dextrans) other methods (e.g., dextrans with differently colored dyes) to detect blood flow should be considered.
Nevertheless, this 2P-IVM imaging methodology allows direct imaging of adventitial and intimal microvessels but also the quantification of the permeability of the microvessels by 40 kDa extravasation in a more realistic test environment compared to post-mortem tissue. Moreover, this technique could be easily adapted to further investigated the dynamics of intraplaque angiogenesis and intraplaque hemorrhage. By injecting fluorescent-labelled cells (such as erythrocytes or inflammatory cells), their extravasation, transmigration, and interaction with the endothelium could easily be monitored and quantified in vivo. Therefore, this 2P-IVM method is a promising approach to validate therapeutic angiogenic interventions targeting advance atherosclerosis in preclinical models in real-time.
4. Materials and Methods
4.1. Animals
All experiments were carried out with approval of the Animal Welfare Committee of the Leiden Medical University Center (28 March 2018; approval number 116002106645-18-096) and in compliance the Directive 2010/63/EU of the European Parliament. Male ApoE3*Leiden mice (n = 12), 10–16 weeks old, were fed with a high-cholesterol inducing diet (2.5% cholesterol and 0.05% cholate w/w, AB diets, Woerden, The Netherlands) during all the experiment. Mice were housed under standard laboratory conditions and received food and water ad libitum.
4.2. Vein Graft Surgery
VG surgery consists of the interposition of the caval vein from a donor mouse in the carotid artery of a recipient mouse, as described before [
8]. In brief, the recipient mouse was fixed in a supine position, and an incision was made in the neck. The parotid glands were put aside exposing the right carotid artery. Next, the carotid artery was ligated and cut in middle, a cuff was placed at both ends of the arterial segments. Subsequently, the ends were everted over the cuffs and ligated. The vena cava was harvested from the donor mouse and positioned between the carotid artery by sleeving it over the cuffs and tightened with 8/0 sutures. Pulsatile flow through the venous conduit confirms a successful procedure. Finally, the parotid gland is put back in position and the skin is sutured. Within 28 days after the surgery, the VG lesions develop from a few cell layers at the start of the engraftment, to a massive thickened vessel wall [
23].
Mice were anesthetized (intraperitoneally) with 5 mg/kg of midazolam (Roche Diagnostics, Basel, Switzerland), 0.5 mg/kg of dexmedetomidine (Orion Corporation, Espoo, Finland) and 0.05 mg/kg of fentanyl (Janssen Pharmaceutical, Beerse, Belgium). After the surgery, the anesthesia was antagonized with 2.5 mg/kg of atipamezole (Orion Corporation,) and 0.5 mg/kg of flumazenil (0.5 mg/kg, Fresenius Kabi, Bad Homburg vor der Höhe, Germany). Then, 0.1 mg/kg of buprenorphine (MSD Animal Health, Boxmeer, The Netherlands) was given for pain relief.
4.3. Two-Photon Intravital Microscopy
Four weeks after the surgery (
Figure 1A), mice (
n = 12) were anesthetized and prepared for intravital imaging on a Zeiss LSM 710 NLO upright multiphoton microscope equipped with a Mai Tai Deep See multiphoton laser (690–1040 nm).
Neck and ear regions were shaved, and a catheter was placed in the tail vein for intravenous injections. To image fenestrated microvessels (in the parotid glands) and plaque microvessels (in the VGs), mice were placed in a supine position on an inset located under the microscope (
Figure 1B). Parotid glands were extracorporated, and VGs were carefully exposed from the surrounding connective tissue. To image continuous microvessels (in the ear skin), mice were placed in prone position on the inset (
Figure 1B). In both positions, temperature was controlled and breathing was monitored. On top of the target tissue, a metal frame was placed. This metal frame was specially designed to allow the use of a water immersion objective (W Plan Apochromat 20×/1.0 DIC M27 75 mm objective) in the different tissues of the mouse body (
Figure 1B).
To select areas of interest in the different microvessels types (continuous, fenestrated and plaque), mice were injected with 50 µL of 100 mg/mL FITC-conjugated 2000 kDa dextran (a blood tracer), via the vein catheter (
Figure 1B). These areas were imaged by cycles of time-lapse Z-stacks (40 s each) over 20 min by multiphoton excitation at 488 nm (FITC) and 555 nm (TRITC). Emission was collected by two LSM PMTs at 500–558 nm (FITC) and 578–700 nm (TRITC). At the 5th frame of the time-lapse (t5) a mixture of 100 µL of a 100 mg/mL 40 kDa Dextran-TRITC (42874, Sigma-Aldrich, Zwijndrecht, The Netherlands) and 2000 kDa Dextran-FITC (FD2000S, Sigma-Aldrich, Zwijndrecht, The Netherlands) solution was injected to assess vessel permeability in real-time in all the groups (
Figure 1C).
To study plaque angiogenesis in particular, a separate group of three mice was used to study the architecture of intimal and adventitial microvessels. Then, 50–150µm depth Z-stacks were acquired by multiphoton excitation at 488 nm, after injection of 50 µL 100 mg/mL FITC-conjugated 2000 kDa dextran, via the tail vein catheter. Directly after imaging, all mice were euthanized by exsanguination.
4.4. Data Analysis
4.4.1. Quantification of FITC and TRITC Fluorescence Integrated Density Inside and Outside the Vessel Structures
To quantify FITC and TRITC fluorescence integrated density inside and outside the vessel structures, we converted 2P-IVM time-lapse acquisition files in maximal intensity Z-stack projections (RBG). Based on the literature, 2000 kDa-size dextrans are less prone to extravasate in microvascular structures due to their big size [
17]. Therefore, we used the FITC channel to apply a tight automatic threshold and define vessel structures surface, denominated as Vessel Mask. Vessel Mask comprises all pixels inside the vessel value as 1 (defined by the automatic threshold) and all the pixels outside the vessel value as 0, at all the timelapsess. The Outside Mask was created via inversion. Both Vessel Mask and Outside Mask were then multiplied by the FITC and TRITC channel, generating four additional files: FITC pixels inside the vessel, FITC pixels outside the vessel, TRITC pixels inside the vessel, TRITC pixels outside the vessel.
For the all the six files, relative fluorescence intensity (RFI) was calculated by RawIntegrated density function. Since the area between intravascular and extravascular space are different between mice and organs, FITC and TRITC integrated densities were divided by the number of pixels of theVessel Mask or Outside Mask. Subsequently, FITC and TRITC fluorescence integrated densities reflect all the pixel intensities inside or outside the vessel structures. Since 40 kD and 2000 kDa dextran injection was detected intravitally at the 6th frame of the time-lapse (t6), FITC and TRITC fluorescence integrated intensities, intra and extravascular, were normalized to t6 values, and plotted in XY graphs.
4.4.2. Video Processing of FITC and TRITC Fluorescence Time-Lapse Series Acquired by 2P-IVM
To evaluate 40 kDa and 2000 kDa dextran extravasation patterns, we generated video time-lapse series of maximal FITC and TRITC fluorescence Z-stack projections. 2000 kDa dextran is visualized in the green channel and 40 kDa dextran is visualized in the red channel.
Plaque microvessels in advanced atherosclerotic VG lesions were processed by maximal FITC fluorescence Z-stack projections. Moreover, we generated 3D stack projections using Imaris 3D rendering software (Imaris, Zurich, Switzerland). In both renderings 2000 kDa-FITC dextran is visualized in the green channel.
To distinguish between adventitial and intimal microvessels, we used the temporal color-coding plugin in ImageJ FIJI that used a color LUT based on Z-depth and projects this in a maximum projection image.
4.5. Statistical Analysis
Results are shown as mean ± standard deviation error (SD). One-way ANOVA was used to compare differences between groups. Differences were considered significant when p * ≤ 0.05, p ** ≤ 0.01, p *** ≤ 0.001 or p **** ≤ 0.001.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




