Genome-Wide Identification of the Expansin Gene Family and Its Potential Association with Drought Stress in Moso Bamboo

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification of the Expansin Genes in Moso Bamboo
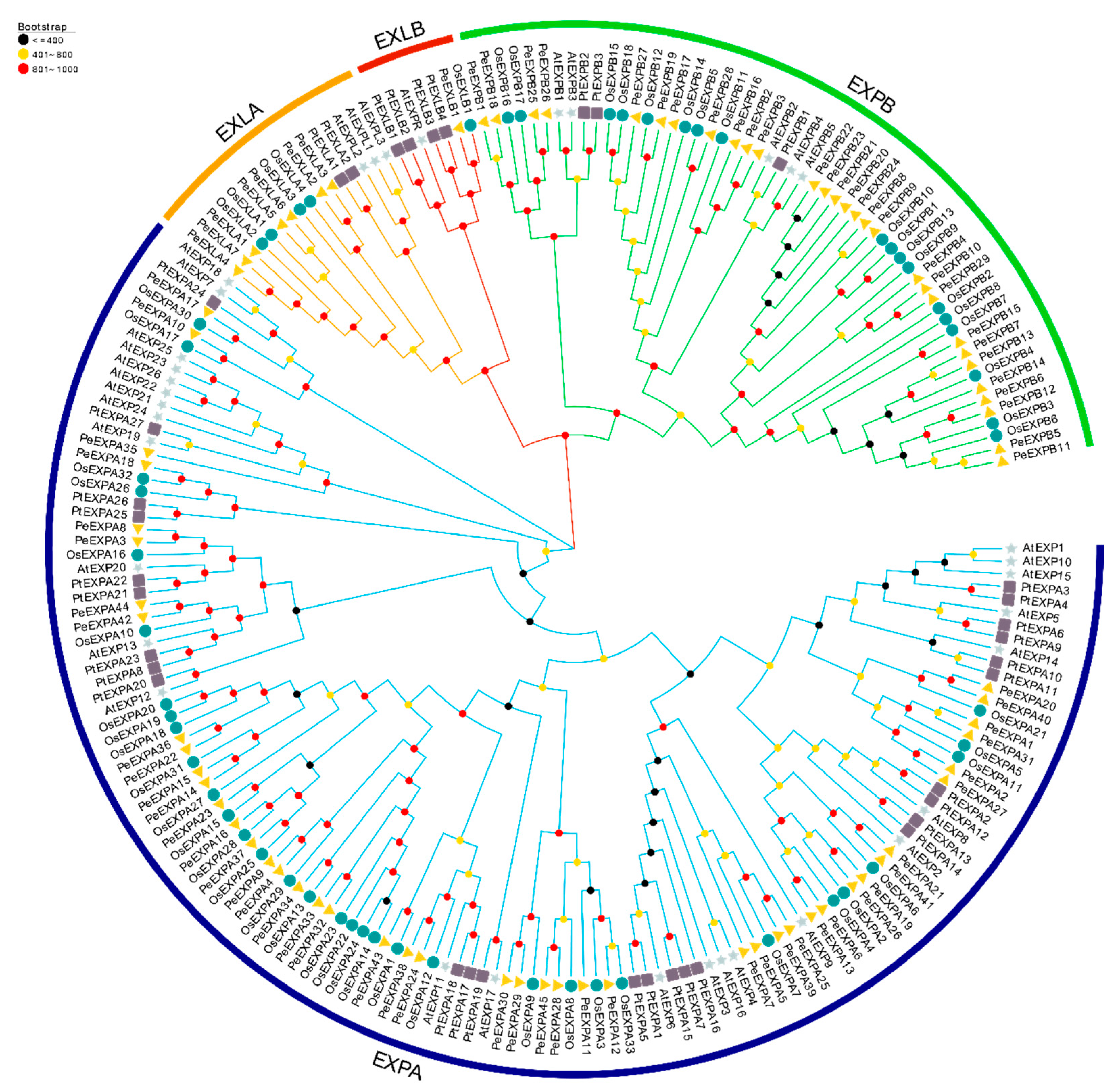
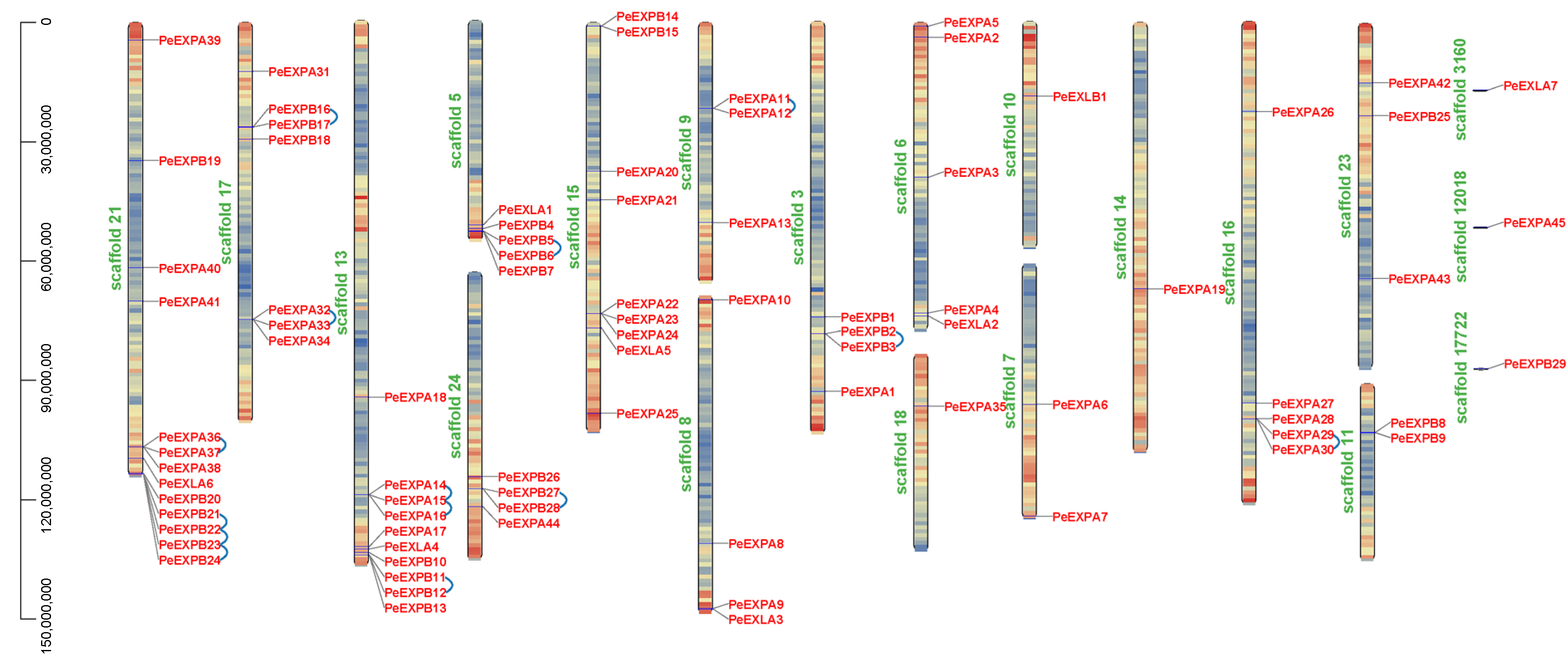
2.2. Phylogenetic Analysis of Expansins in the Four Different Plant Species, Distribution of Expansin Genes on Scaffolds
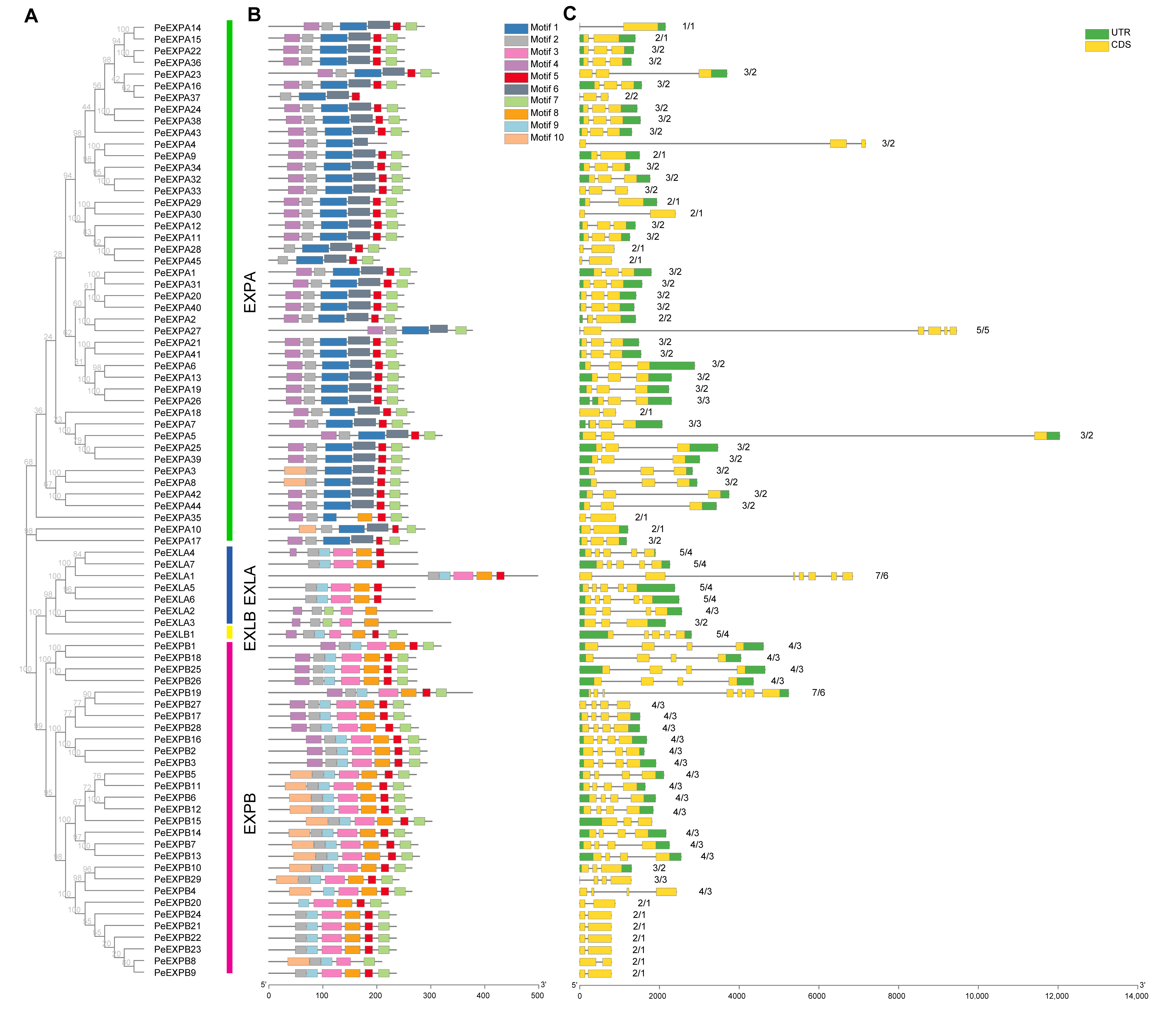
2.3. Gene Structure, Conserved Protein Motifs of PeEXs
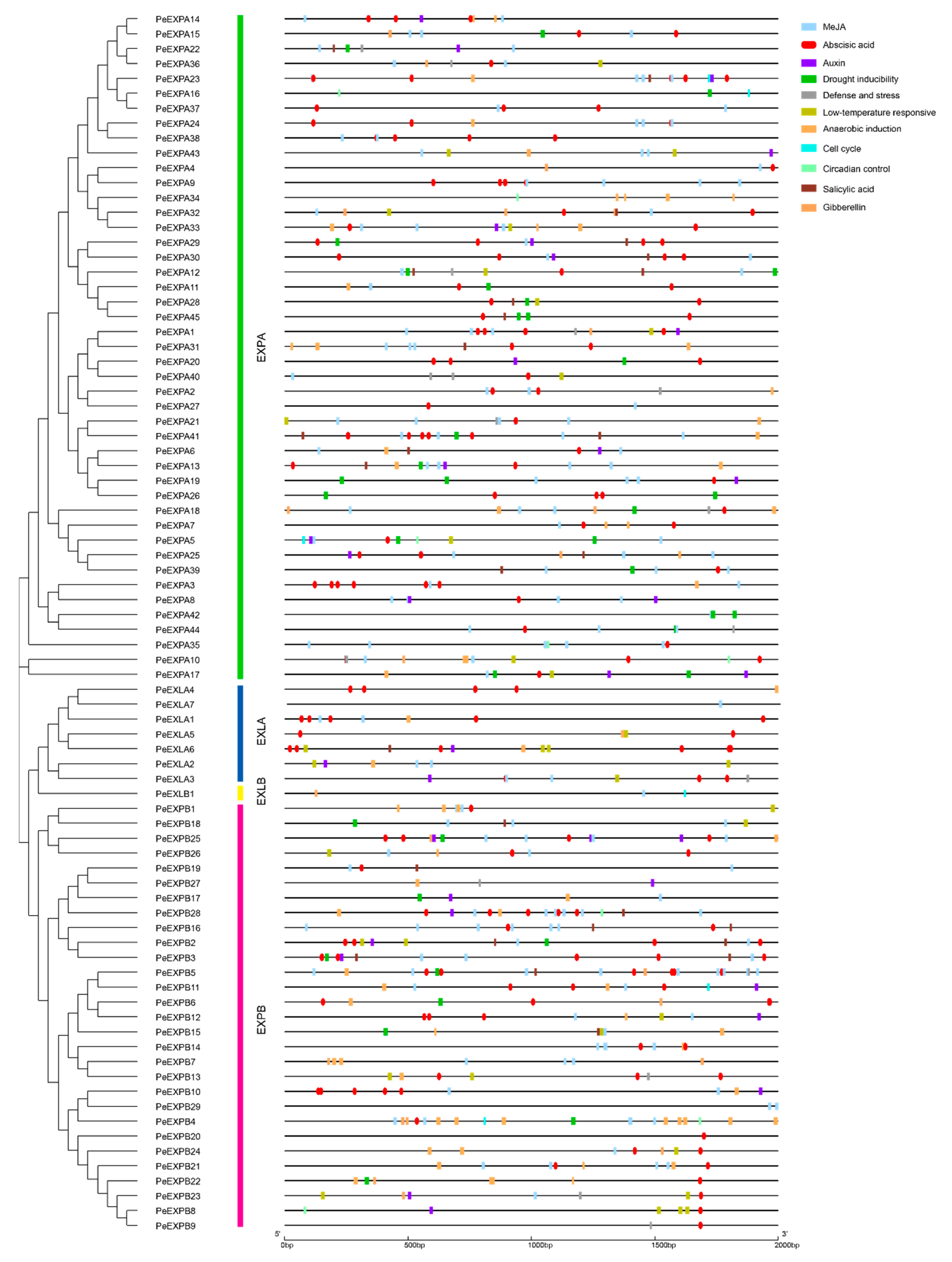
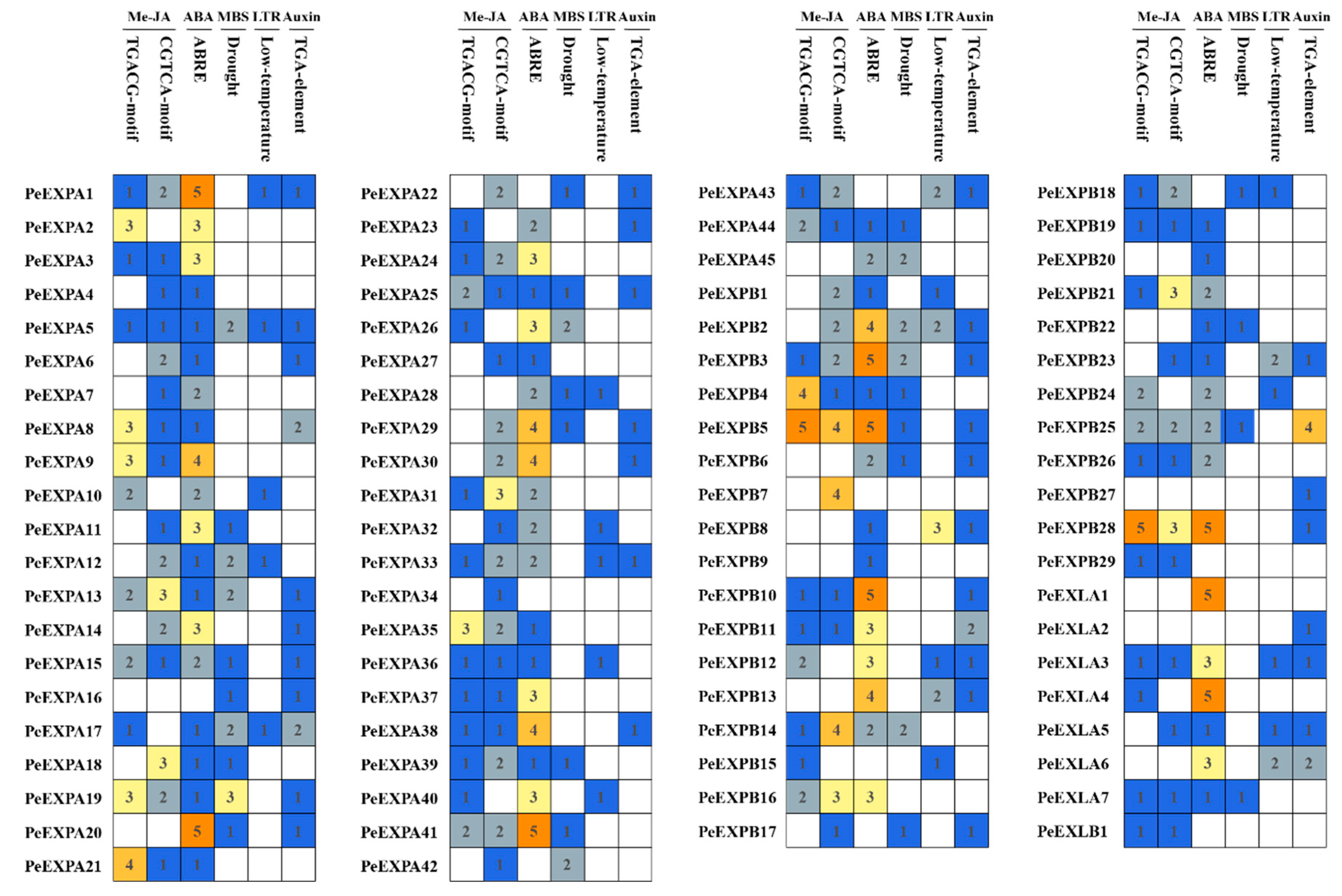
2.4. Detection of Cis-Regulatory Elements in the Promoter Regions of PeEXs
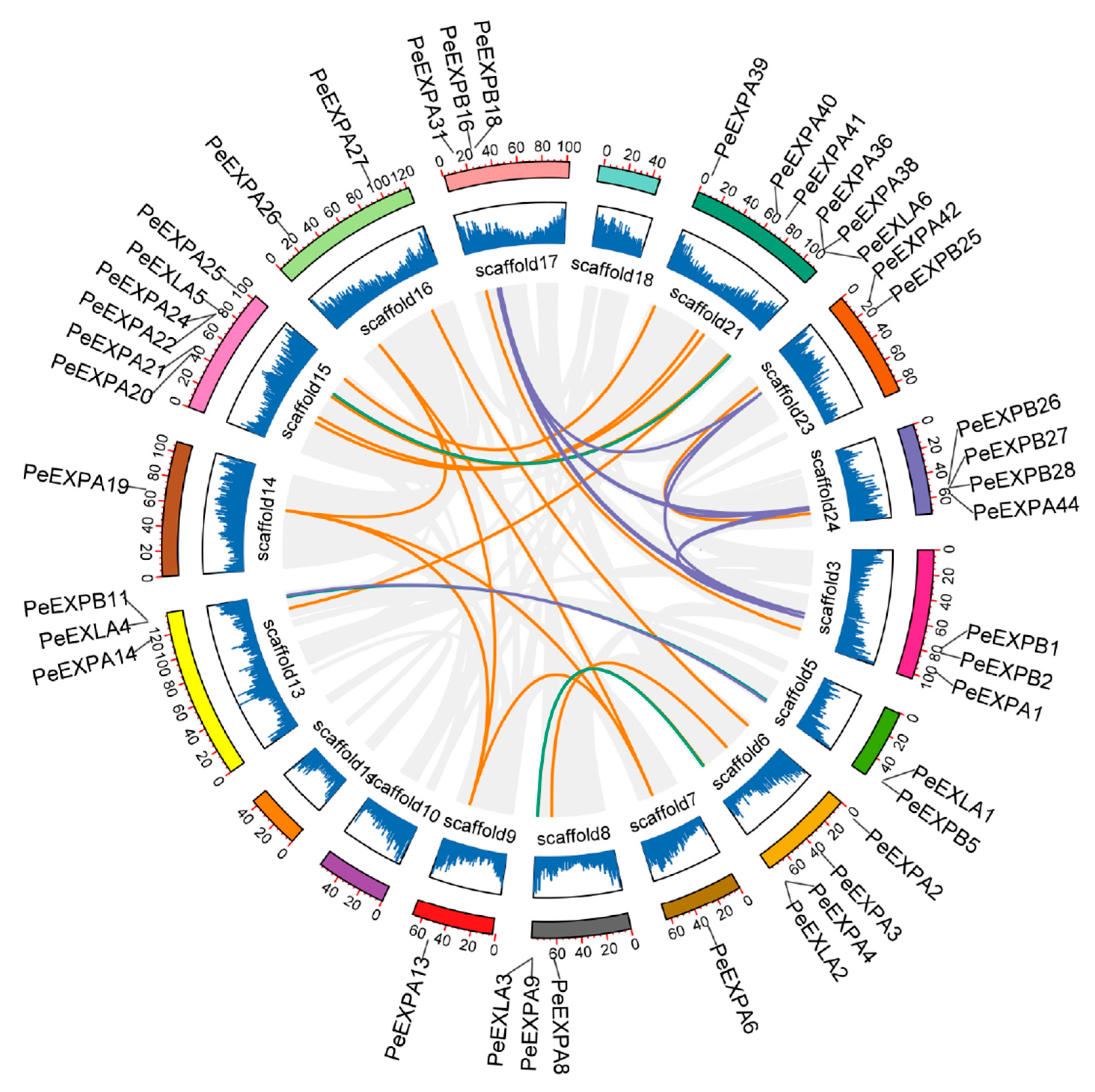
2.5. Synteny Analysis
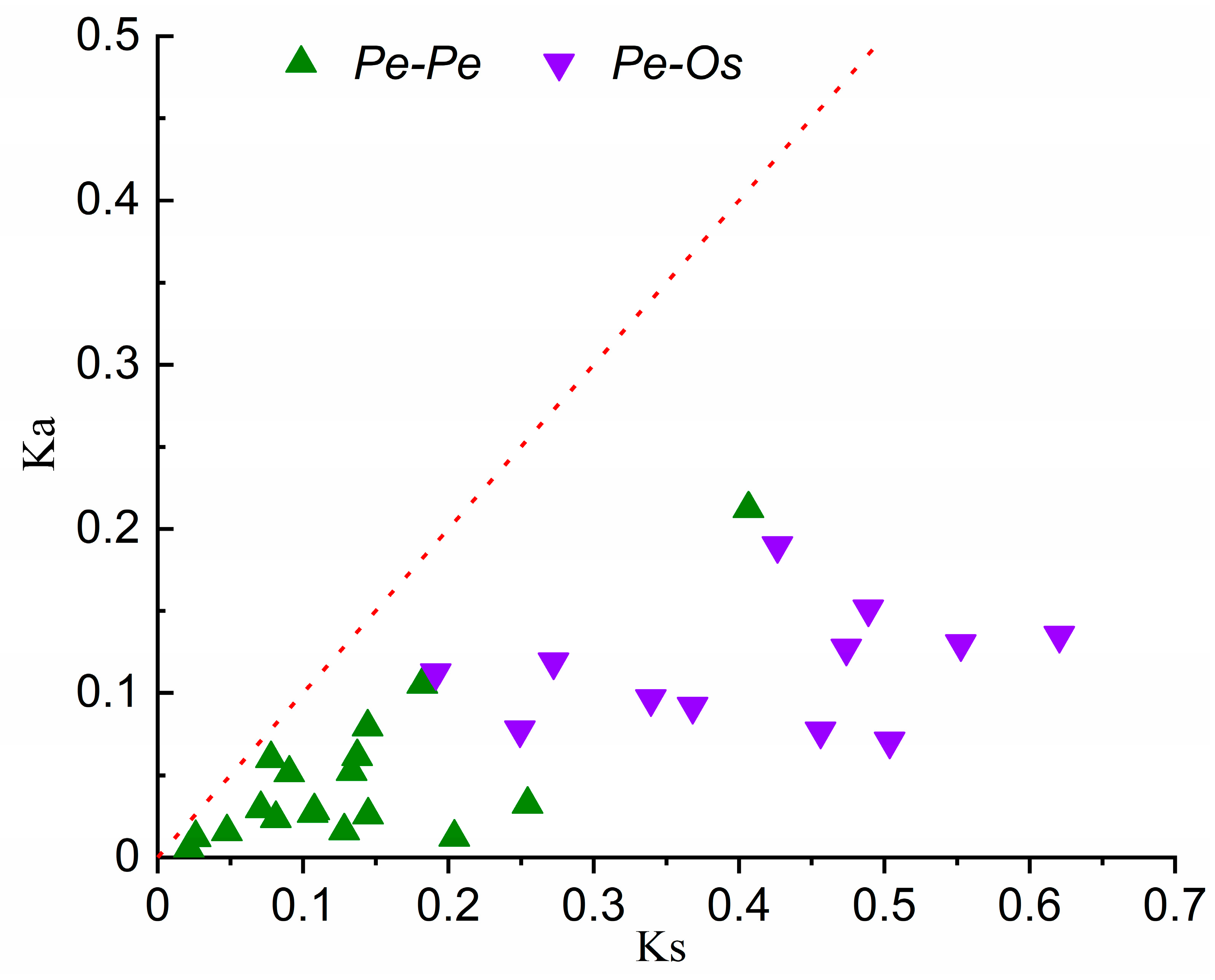
2.6. Evolutionary and Divergence Patterns of the Expansin Genes
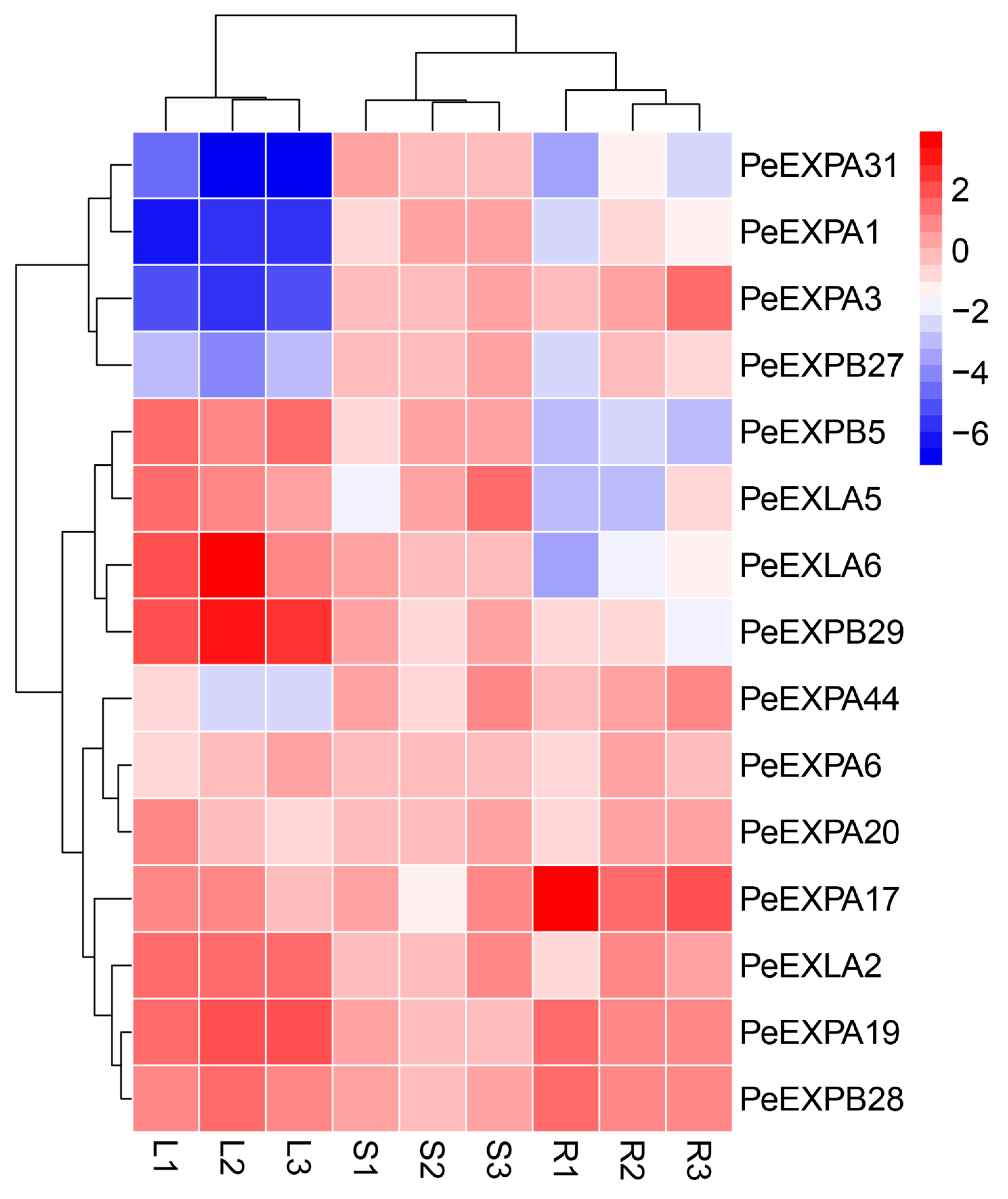
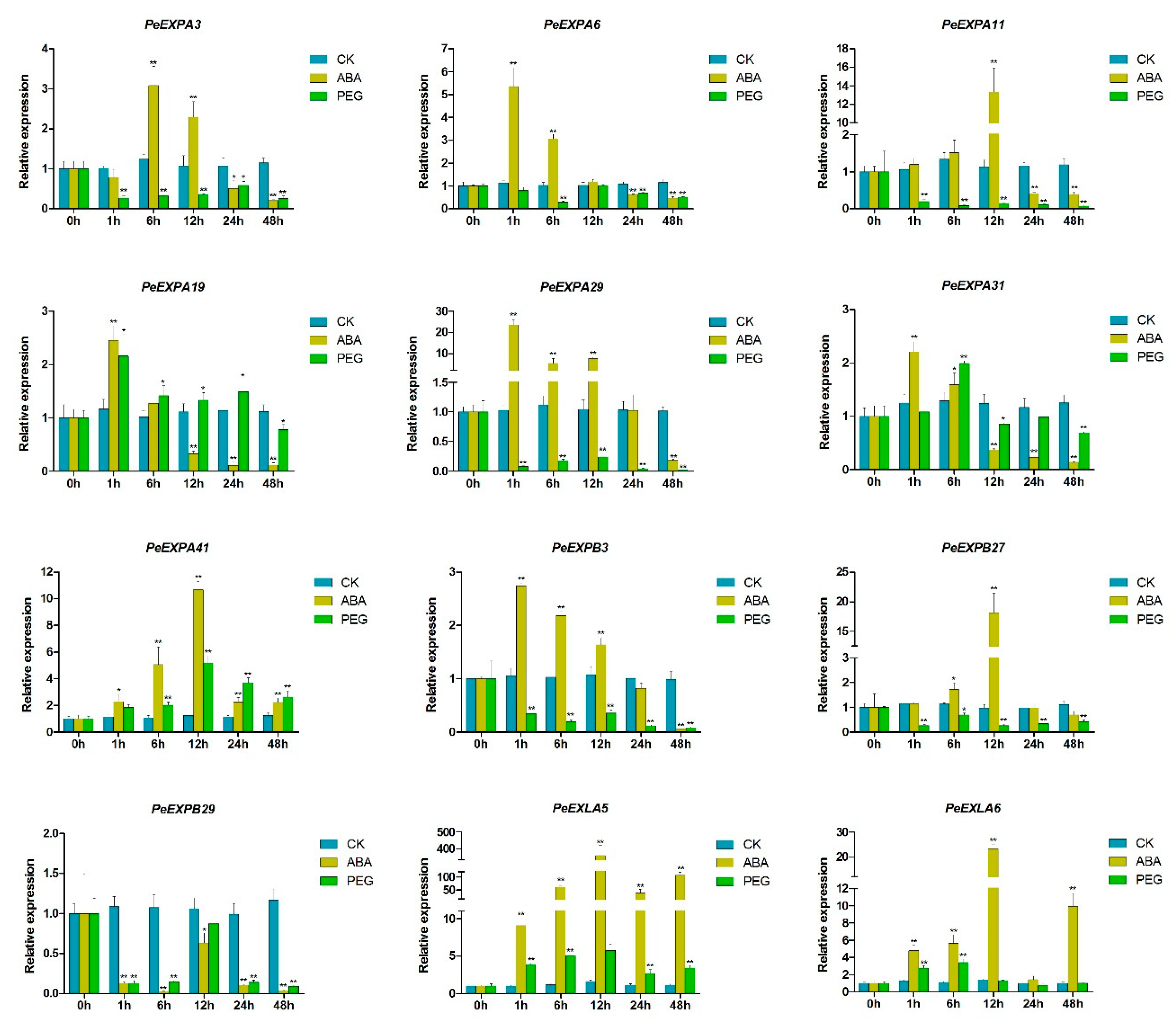
2.7. qRT-PCR Expression Analysis of the Expansin Genes in Moso Bamboo
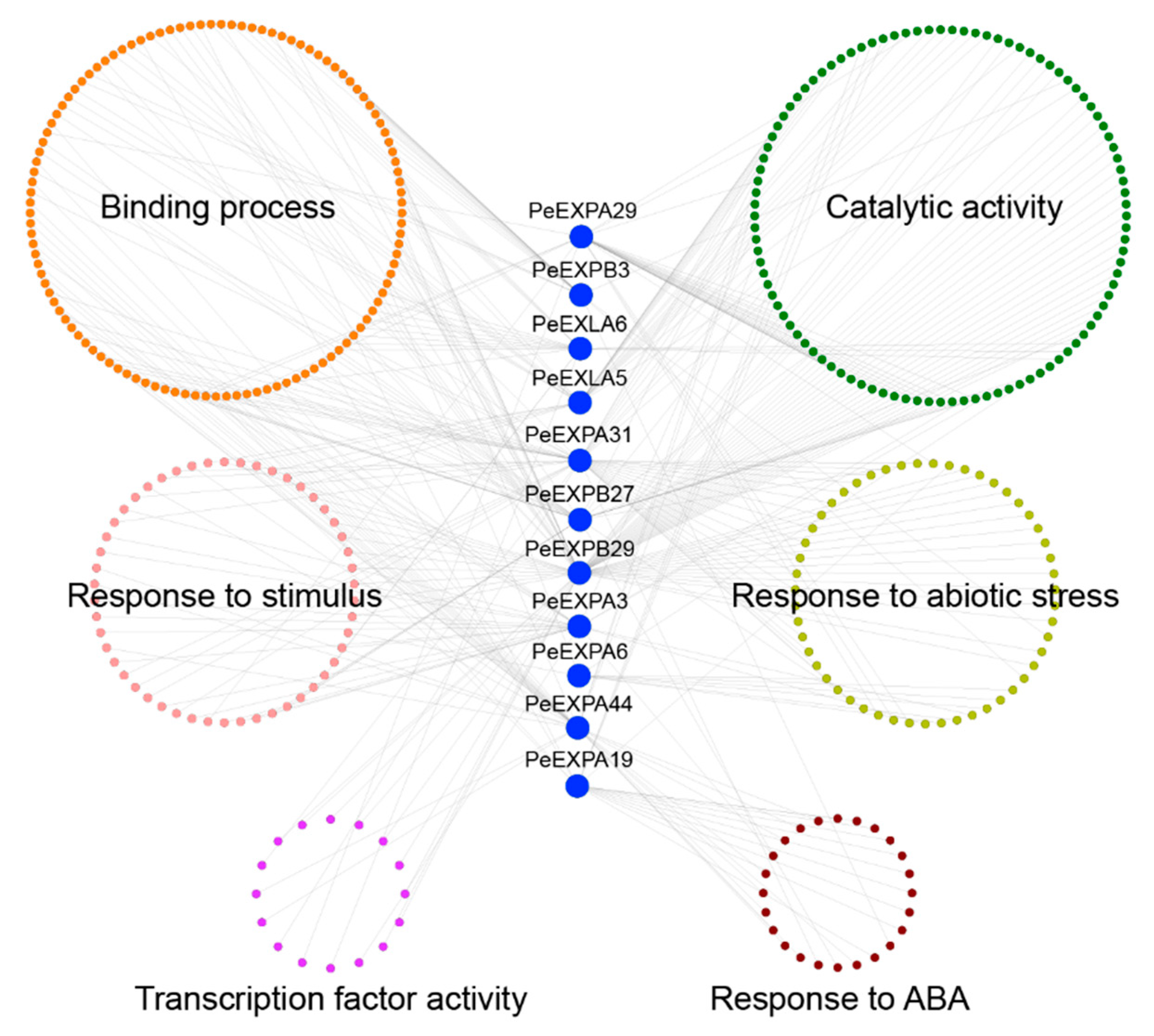
2.8. Co-Expression Analysis of the Expansin Genes in Moso Bamboo
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Genome-Wide Identification of Expansin Genes
5.2. Exon–Intron Structure, Conserved Motif, Genic Physical Location on Scaffolds and Cis-Regulatory Elements Analysis
5.3. Phylogenetic Tree Construction
5.4. Synteny and Gene Duplication Analyzes of the Expansin Genes in Moso Bamboo
5.5. Co-Expression Network Construction
5.6. Plant Materials, Growth Conditions and Stresses Treatment
5.7. RNA Isolation and Quantitative RT-PCR
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PeEXs | Expansin genes of Phyllostachys edulis |
| OsEXs | Expansin genes of Oryza sativa |
| Mya | Million years ago |
| ABRE | Abscisic acid response element |
| G2A | Group-II pollen allergen protein |
| NJ | Neighbor-joining |
| PEG | Polyethylene glycol |
| MeJA | Methyl jasmonate |
| MYB | Myeloblastosis |
| Ka | Nonsynonymous substitution rates |
| Ks | Synonymous substitution rates |
| GO | Gene ontology |
| WT | Wild type |
| TaMPS | Homolog to OsMPS and named according to OsMPS |
| SMART | Simple modular architecture research tool |
| PFAM | Database of protein families |
| MEME | Multiple em for motif elicitation |
| ROX | ROX reference dye |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
References
- Zhao, H.; Zhao, S.; Bamboo, I.N.; International Network for Bamboo and Rattan; Fei, B.; Liu, H.; Yang, H.; Dai, H.; Wang, D.; Jin, W.; et al. Announcing the Genome Atlas of Bamboo and Rattan (GABR) project: promoting research in evolution and in economically and ecologically beneficial plants. GigaScience 2017, 6, gix046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J. Chinese moso bamboo: Its importance. Bamboo 2001, 22, 5–7. [Google Scholar]
- Peng, Z.; Lu, Y.; Li, L.; Zhao, Q.; Feng, Q.; Gao, Z.; Lu, H.; Hu, T.; Yao, N.; Liu, K. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J. Climate change and forest disturbances: Climate change can affect forests by altering the frequency, intensity, duration, and timing of fire, drought, introduced species, insect and pathogen outbreaks, hurricanes, windstorms, ice storms, or landslides. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, D.J. Loosening of plant cell walls by expansins. Nature 2000, 407, 321–326. [Google Scholar] [CrossRef]
- Choi, D.; Lee, Y.; Cho, H.T.; Kende, H. Regulation of Expansin Gene Expression Affects Growth and Development in Transgenic Rice Plants. Plant Cell 2003, 15, 1386–1398. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Minjie, W.; Limei, Y.; Rui, H.; Imran, A.; Yinbo, G.; John, S. AtEXP2 Is Involved in Seed Germination and Abiotic Stress Response in Arabidopsis. PLoS ONE 2014, 9, e85208. [Google Scholar]
- Cho, H.T.; Cosgrove, D.J. Regulation of Root Hair Initiation and Expansin Gene Expression in Arabidopsis. Plant Cell 2002, 14, 3237–3253. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Choi, H.-S.; Cho, H.-T. Root hair-specific EXPANSIN A7 is required for root hair elongation in Arabidopsis. Mol. Cells 2011, 31, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, P.; Duarte, P.R.; Rance, G.A.; Schubert, M.; Vordermaier, V.; Dai Vu, L.; Murphy, E.; Barro, A.V.; Swarup, K.; Moirangthem, K. EXPANSIN A1-mediated radial swelling of pericycle cells positions anticlinal cell divisions during lateral root initiation. Proc. Natl. Acad. Sci. USA 2019, 116, 8597–8602. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-K.; Ahn, J.H.; Song, S.-K.; Do Choi, Y.; Lee, J.S. Expression of an expansin gene is correlated with root elongation in soybean. Plant Physiol. 2003, 131, 985–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Luo, Y.; Wang, G.; Feng, C.; Li, H. Genome-wide identification of expansin genes in Brachypodium distachyon and functional characterization of BdEXPA27. Plant Sci. 2020, 296, 110490. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-T.; Cosgrove, D.J. Altered expression of expansin modulates leaf growth and pedicel abscission in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 9783–9788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Jeong, D.H.; Min, C.P.; An, G. Characterization and transcriptional expression of the alpha-expansin gene family in rice. Mol. Cells 2005, 20, 210–218. [Google Scholar]
- Cosgrove, D.J. New genes and new biological roles for expansins. Curr. Opin. Plant Biol. 2000, 3, 73–78. [Google Scholar] [CrossRef]
- Cosgrove, D.J.; Bedinger, P.; Durachko, D.M. Group I allergens of grass pollen as cell wall-loosening agents. Proc. Natl. Acad. Sci. USA 1997, 94, 6559–6564. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, C.R.; Pimentel, P.; Dotto, M.C.; Civello, P.M.; Martínez, G.A.; Herrera, R.; Moya-León, M.A. Expression of five expansin genes during softening of Fragaria chiloensis fruit: Effect of auxin treatment. Postharvest Biol. Technol. 2009, 53, 51–57. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, B.; Li, C.; Lei, C.; Gong, M. A comprehensive expression analysis of the expansin gene family in potato (Solanum tuberosum) discloses stress-responsive expansin-like B genes for drought and heat tolerances. PLoS ONE 2019, 14, e0219837. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, G.; An, J.; Li, Q.; Chen, Y.; Zhao, X.; Wu, J.; Wang, Y.; Hao, Q.; Wang, W. Expansin gene TaEXPA2 positively regulates drought tolerance in transgenic wheat (Triticum aestivum L.). Plant Sci. 2020, 298, 110596. [Google Scholar] [CrossRef]
- Marowa, P.; Ding, A.; Kong, Y. Expansins: Roles in plant growth and potential applications in crop improvement. Plant Cell Rep. 2016, 35, 949–965. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Han, Y.; Zhang, M.; Zhou, S.; Kong, X.; Wang, W. Overexpression of the wheat expansin gene TaEXPA2 improved seed production and drought tolerance in transgenic tobacco plants. PLoS ONE 2016, 11, e0153494. [Google Scholar] [CrossRef] [PubMed]
- Shcherban, T.Y.; Shi, J.; Durachko, D.M.; Guiltinan, M.J.; McQueen-Mason, S.J.; Shieh, M.; Cosgrove, D.J. Molecular cloning and sequence analysis of expansins-a highly conserved, multigene family of proteins that mediate cell wall extension in plants. Proc. Natl. Acad. Sci. USA 1995, 92, 9245–9249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marga, F.; Grandbois, M.; Cosgrove, D.J.; Baskin, T.I. Cell wall extension results in the coordinate separation of parallel microfibrils: Evidence from scanning electron microscopy and atomic force microscopy. Plant J. 2005, 43, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, P.K.; Nath, P. MaExp1, an ethylene-induced expansin from ripening banana fruit. Plant Sci. 2004, 167, 1–1358. [Google Scholar] [CrossRef]
- Cho, H.T.; Kende, H. Tissue localization of expansins in deepwater rice. Plant J. 1998, 15, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Georgelis, N.; Nikolaidis, N.; Cosgrove, D.J. Bacterial expansins and related proteins from the world of microbes. Appl. Microbiol. Biotechnol. 2015, 99, 3807–3823. [Google Scholar] [CrossRef] [Green Version]
- Nikolas, N.; Nicole, D.; Cosgrove, D.J. Plant Expansins in Bacteria and Fungi: Evolution by Horizontal Gene Transfer and Independent Domain Fusion. Mol. Biol. Evol. 2014, 31, 376–386. [Google Scholar] [CrossRef] [Green Version]
- Yennawar, N.H.; Li, L.-C.; Dudzinski, D.M.; Tabuchi, A.; Cosgrove, D.J. Crystal structure and activities of EXPB1 (Zea m 1), a β-expansin and group-1 pollen allergen from maize. Proc. Natl. Acad. Sci. USA 2006, 103, 14664–14671. [Google Scholar] [CrossRef] [Green Version]
- De Marco, A.; Menegatti, E.; Guarneri, M. pH and temperature effects on the molecular conformation of the porcine pancreatic secretory trypsin inhibitor as detected by proton nuclear magnetic resonance. Biochemistry 1982, 21, 222–229. [Google Scholar] [CrossRef]
- Sampedro, J.; Cosgrove, D.J. The expansin superfamily. Genome Biol. 2005, 6, 242. [Google Scholar] [CrossRef] [Green Version]
- Kende, H.; Bradford, K.; Brummell, D.; Cho, H.T.; Voesenek, L.A.C.J. Nomenclature for members of the expansin superfamily of genes and proteins. Plant Mol. Biol. 2004, 55, 311–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Choi, D.; Kende, H. Expansins: Ever-expanding numbers and functions. Curr. Opin. Plant Biol. 2001, 4, 527–532. [Google Scholar] [CrossRef]
- Wu, Y.; Meeley, R.B.; Cosgrove, D.J. Analysis and Expression of the alpha-Expansin and beta-Expansin Gene Families in Maize. Plant Physiol. 2001, 126, 222–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamurthy, P.; Muthusamy, M.; Kim, J.A.; Jeong, M.-J.; Lee, S.I. Brassica rapa expansin-like B1 gene (BrEXLB1) regulate growth and development in transgenic Arabidopsis and elicits response to abiotic stresses. J. Plant Biochem. Biotechnol. 2019, 28, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yan, H.; Chen, W.; Liu, J.; Jiang, C.; Jiang, H.; Zhu, S.; Cheng, B. Genome-wide identification and characterization of maize expansin genes expressed in endosperm. Mol. Genet. Genom. 2014, 289, 1061–1074. [Google Scholar] [CrossRef]
- Han, Z.; Liu, Y.; Deng, X.; Liu, D.; Liu, Y.; Hu, Y.; Yan, Y. Genome-wide identification and expression analysis of expansin gene family in common wheat (Triticum aestivum L.). BMC Genom. 2019, 20, 1–19. [Google Scholar] [CrossRef]
- Sampedro, J.; Carey, R.E.; Cosgrove, D.J. Genome histories clarify evolution of the expansin superfamily: New insights from the poplar genome and pine ESTs. J. Plant Res. 2006, 119, 11–21. [Google Scholar] [CrossRef]
- Zhao, H.; Gao, Z.; Wang, L.; Wang, J.; Wang, S.; Fei, B.; Chen, C.; Shi, C.; Liu, X.; Zhang, H. Chromosome-level reference genome and alternative splicing atlas of moso bamboo (Phyllostachys edulis). GigaScience 2018, 7, giy115. [Google Scholar] [CrossRef]
- Zhao, H.; Peng, Z.; Fei, B.; Li, L.; Hu, T.; Gao, Z.; Jiang, Z. BambooGDB: A bamboo genome database with functional annotation and an analysis platform. Database 2014, 2014, bau006. [Google Scholar] [CrossRef] [Green Version]
- Chou, K.-C.; Shen, H.-B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xu, R.; Gao, Z.; Chen, C.; Jiang, Z.; Shu, H. A genome-wide analysis of the expansin genes in Malus×Domestica. Mol. Genet. Genom. 2014, 289, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.; Rao, J.; Ren, X.; Li, J. Studying progress of plant cell wall proteins expansins. Chin. Agric. Sci. Bull. 2005, 9, 112–115. [Google Scholar]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, S.; Greiner, S. Growth control by cell wall pectins. Protoplasma 2012, 249, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Bielenberg, D.; Brown, K.M.; Lynch, J.P. Regulation of root hair density by phosphorus availability in Arabidopsis thaliana. Plant, Cell Environ. 2001, 24, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.; Wenwen, Y.; Zang, G.; Kang, Z.; Zhang, Z.; Huang, J.; Wang, G. OsEXPB2, a β-expansin gene, is involved in rice root system architecture. Mol. Breed. 2015, 35, 41. [Google Scholar] [CrossRef]
- Guo, Y.M.; Qin, Z.L.; Zhang, G.P.; Jin, L.; Wang, G.X.; Huang, J.L. The Effect of OsEXPA8 Gene on Suspension Cell Cycle and Size in Rice(Oryza sativa). J. Agric. Biotechnol. 2016, 24, 18–25. [Google Scholar]
- Wei, P.C.; Zhang, X.Q.; Zhao, P.; Wang, X.C. Regulation of stomatal opening by the guard cell expansin AtEXPA1. Plant Signal. Behav. 2011, 6, 740–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, A.L.; Kalamaki, M.S.; Kurien, P.A.; Gurrieri, S.; Bennett, A.B. Simultaneous transgenic suppression of LePG and LeExp1 influences fruit texture and juice viscosity in a fresh market tomato variety. J. Agric. Food Chem. 2003, 51, 7450–7455. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.P.; McQueen-Mason, S.J.; Manning, K. Expression of six expansin genes in relation to extension activity in developing strawberry fruit. J. Exp. Bot. 2001, 52, 1437–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.-D.; Gao, Z.; Cantini, C.; Loescher, W.H.; Van Nocker, S. Fruit ripening in sour cherry: Changes in expression of genes encoding expansins and other cell-wall-modifying enzymes. J. Am. Soc. Hortic. Sci. 2003, 128, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Obenland, D.M.; Crisosto, C.H.; Rose, J.K.C. Expansin protein levels decline with the development of mealiness in peaches. Postharvest Biol. Technol. 2003, 29, 11–18. [Google Scholar] [CrossRef]
- Wei, Q.; Jiao, C.; Ding, Y.; Gao, S.; Guo, L.; Chen, M.; Hu, P.; Xia, S.; Ren, G.; Fei, Z. Cellular and molecular characterizations of a slow-growth variant provide insights into the fast growth of bamboo. Tree Physiol. 2018, 38, 641–654. [Google Scholar] [CrossRef]
- Tuteja, N. Abscisic acid and abiotic stress signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Wang, M.; Shi, Z.; Miao, X. OsEXPA10 mediates the balance between growth and resistance to biotic stress in rice. Plant Cell Rep. 2018, 37, 993–1002. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, H.; Yang, R.; Xu, X.; Liu, X.; Xu, J. Over-expression of PttEXPA8 gene showed various resistances to diverse stresses. Int. J. Biol. Macromol. 2019, 130, 50–57. [Google Scholar] [CrossRef]
- Li, Y.; Wu, B.; Yu, Y.; Yang, G.; Wu, C.; Zheng, C. Genome-wide analysis of the RING finger gene family in apple. Mol. Genet. Genom. 2011, 286, 81–94. [Google Scholar] [CrossRef]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.; Hotz, H.R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L. The Pfam protein families database. Nucleic Acids Res. 2008, 32, D138. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Lu, Z.; Jin, K.; Qiu, W.; Qiao, G.; Han, X.; Zhuo, R. SPDE: A Multi-functional Software for Sequence Processing and Data Extraction. bioRxiv 2020. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Ying, C.; Jing, J.; Jin, X.; Jiang, H.; Yan, H.; Cheng, B.; Marc, R.R. Genome Duplication and Gene Loss Affect the Evolution of Heat Shock Transcription Factor Genes in Legumes. PLoS ONE 2014, 9, e102825. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.; Conery, J.S. The Evolutionary Fate and Consequences of Duplicate Genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhao, H.; Xu, W.; You, Q.; Yan, H.; Gao, Z.; Su, Z. Co-expression gene network analysis and functional module identification in bamboo growth and development. Front. Genet. 2018, 9, 574. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yang, Y.; Lou, S.; Wei, W.; Zhao, Z.; Ren, Y.; Lin, C.; Ma, L. Genome-Wide Characterization and Gene Expression Analyses of GATA Transcription Factors in Moso Bamboo (Phyllostachys edulis). Int. J. Mol. Sci. 2020, 21, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Ma, J.; Guo, Q.; Li, X.; Wang, H.; Lu, M.; Blazquez, M.A. Selection of Reference Genes for Quantitative Real-Time PCR in Bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e56573. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | EXPA | EXPB | EXLA | EXLB | Total | References |
|---|---|---|---|---|---|---|
| Arabidopsis | 26 | 6 | 3 | 1 | 36 | [30] |
| Rice | 33 | 18 | 4 | 1 | 56 | [30] |
| Poplar | 27 | 3 | 2 | 4 | 36 | [37] |
| Apple | 34 | 1 | 2 | 4 | 41 | [41] |
| Moso bamboo | 45 | 29 | 7 | 1 | 82 |
| Motif | Length | Protein Sequence | PFAM Domain |
|---|---|---|---|
| 1 | 50 | VTVTATNFCPPNYALPSDBGGWCNPPRQHFDMAZPAFEKIAIYRAGIVPV | DPBB_1 |
| 2 | 21 | ALSTALFNDGAGCGACYZIRC | |
| 3 | 37 | HFDLSGTAFGAMAKPGKADQLRHAGIIDIQFRRVPCK | |
| 4 | 29 | HATFYGGSDASGTMGGACGYGNLYSQGYG | |
| 5 | 15 | WTPMSRNWGANWQSB | |
| 6 | 41 | RRVPCVRKGGIRFTINGHSYFELVLVTNVAGAGDVASVSVK | Pollen_allerg_1 |
| 7 | 21 | FRVTSSDGRTLVANBVIPAGW | |
| 8 | 29 | FHVEKGSNPNYLAVLVEYEDGDGDIVAVD | Pollen_allerg_1 |
| 9 | 21 | TKPEACSGEPVTVVITDMNYE | |
| 10 | 41 | GKWLAAKATWYGAPTGAGPDDNGGACGFKBVNQPPFSSMTS |
| Pe–Os | Pe–Pe |
|---|---|
| PeEXPB15/OsEXPB7 | PeEXPA20/PeEXPA40 |
| PeEXPB28/OsEXPB5 | PeEXPA2/PeEXPA27 |
| PeEXPB19/OsEXPB12 | PeEXPA21/PeEXPA41 |
| PeEXLB1/OsEXLB1 | PeEXPA19/PeEXPA26 |
| PeEXPA17/OsEXPA30 | PeEXPA25/PeEXPA39 |
| PeEXPA10/OsEXPA17 | PeEXPA28/PeEXPA45 |
| PeEXPA18/OsEXPA32 | PeEXPA29/PeEXPA30 |
| PeEXPA23/OsEXPA15 | PeEXPA24/PeEXPA38 |
| PeEXPA16/OsEXPA28 | PeEXPA32/PeEXPA33 |
| PeEXPA37/OsEXPA25 | PeEXPA4/PeEXPA9 |
| PeEXPA34/OsEXPA13 | PeEXPA14/PeEXPA15 |
| PeEXPA43/OsEXPA1 | PeEXPA22/PeEXPA36 |
| PeEXPA42/PeEXPA44 | |
| PeEXPA3/PeEXPA8 | |
| PeEXLA2/PeEXLA3 | |
| PeEXPB25/PeEXPB26 | |
| PeEXPB2/PeEXPB3 | |
| PeEXPB10/PeEXPB29 | |
| PeEXPB6/PeEXPB12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, K.-M.; Zhuo, R.-Y.; Xu, D.; Wang, Y.-J.; Fan, H.-J.; Huang, B.-Y.; Qiao, G.-R. Genome-Wide Identification of the Expansin Gene Family and Its Potential Association with Drought Stress in Moso Bamboo. Int. J. Mol. Sci. 2020, 21, 9491. https://doi.org/10.3390/ijms21249491
Jin K-M, Zhuo R-Y, Xu D, Wang Y-J, Fan H-J, Huang B-Y, Qiao G-R. Genome-Wide Identification of the Expansin Gene Family and Its Potential Association with Drought Stress in Moso Bamboo. International Journal of Molecular Sciences. 2020; 21(24):9491. https://doi.org/10.3390/ijms21249491
Chicago/Turabian StyleJin, Kang-Ming, Ren-Ying Zhuo, Dong Xu, Yu-Jun Wang, Hui-Jin Fan, Bi-Yun Huang, and Gui-Rong Qiao. 2020. "Genome-Wide Identification of the Expansin Gene Family and Its Potential Association with Drought Stress in Moso Bamboo" International Journal of Molecular Sciences 21, no. 24: 9491. https://doi.org/10.3390/ijms21249491