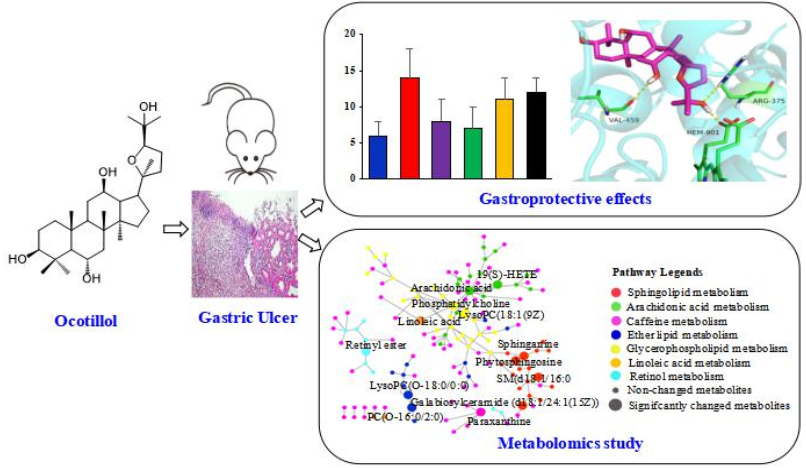
Protective Effect of Ocotillol, the Derivate of Ocotillol-Type Saponins in Panax Genus, against Acetic Acid-Induced Gastric Ulcer in Rats Based on Untargeted Metabolomics

 , , ,
, , ,
Abstract
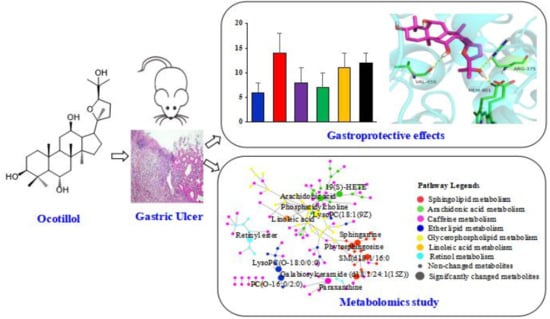
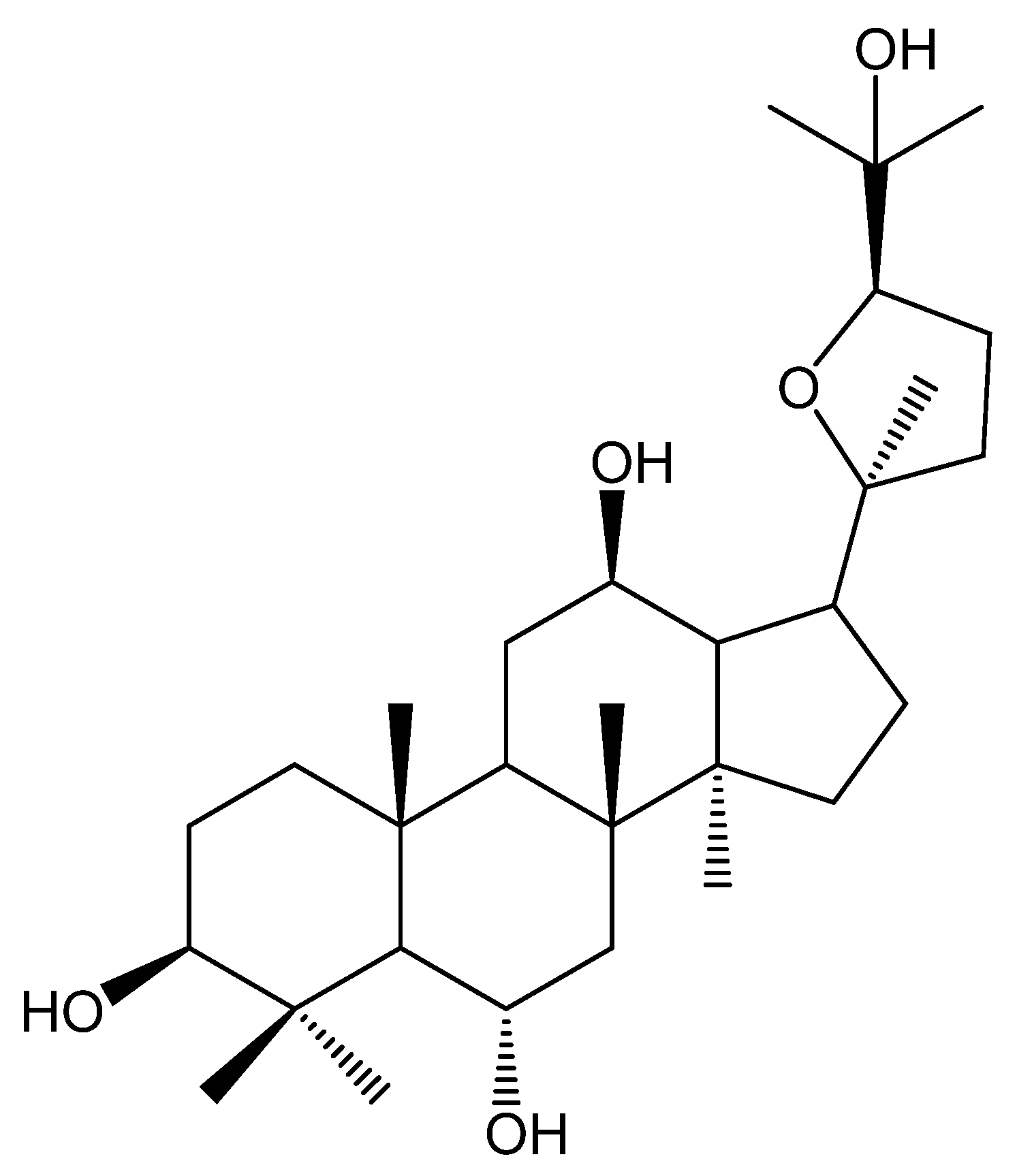
:
1. Introduction
2. Results
2.1. Gastroprotective Effect
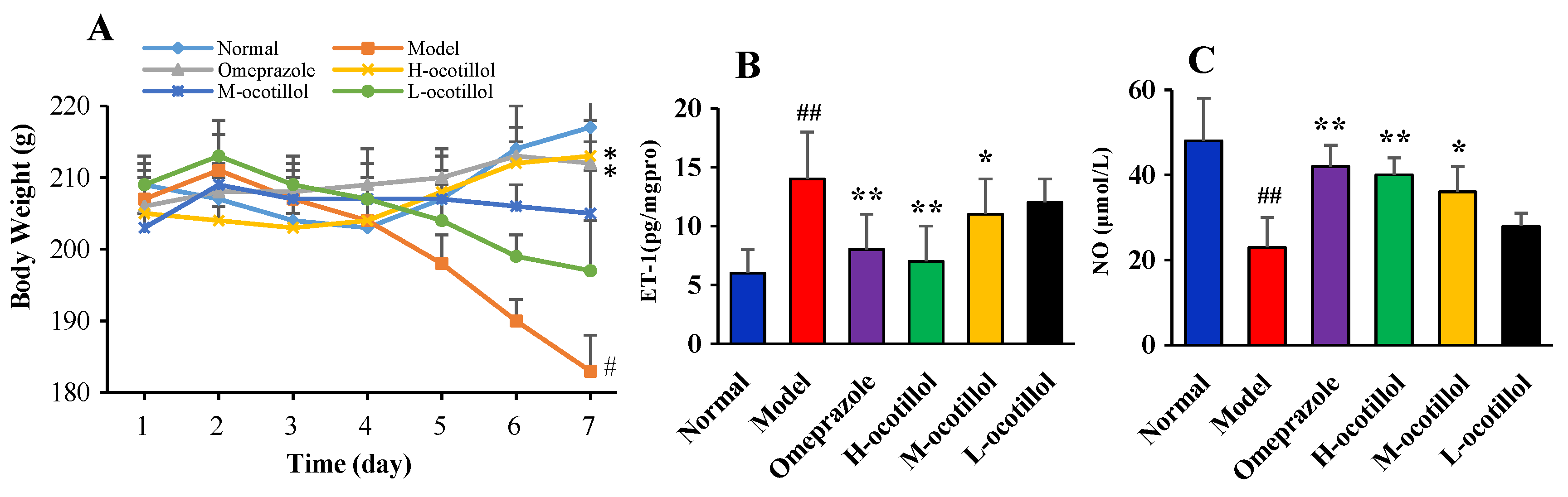
2.1.1. Body Weights of the Rats
2.1.2. Endothelin-1 (ET-1) and Nitric Oxide (NO) Levels in Serum
2.1.3. Epidermal Growth Factor(EGF), Superoxide Dismutase (SOD), and NO Levels in Gastric Mucosa
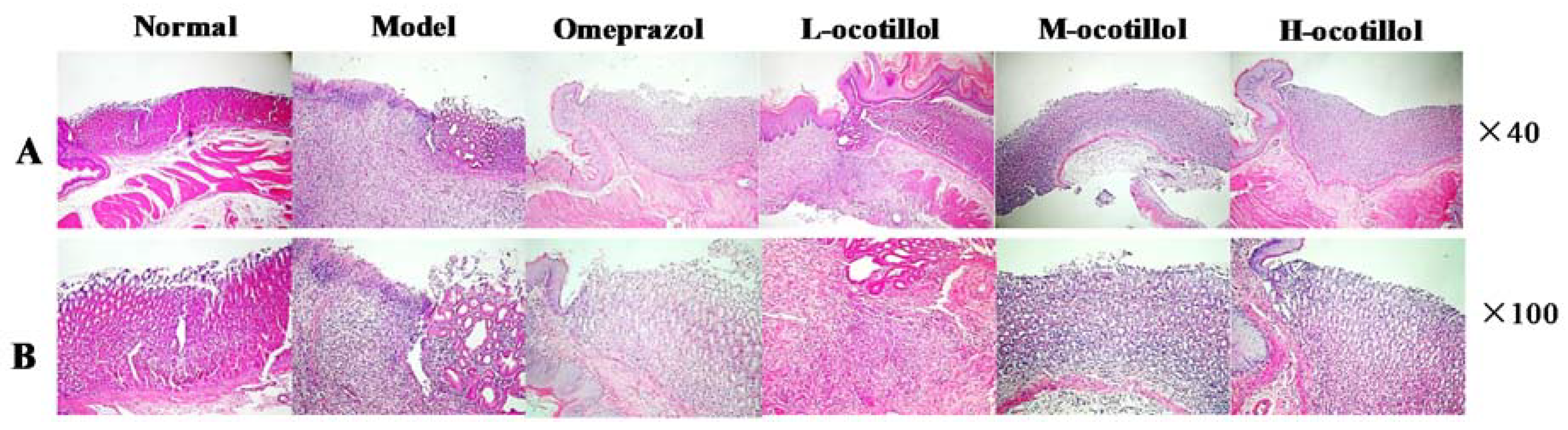
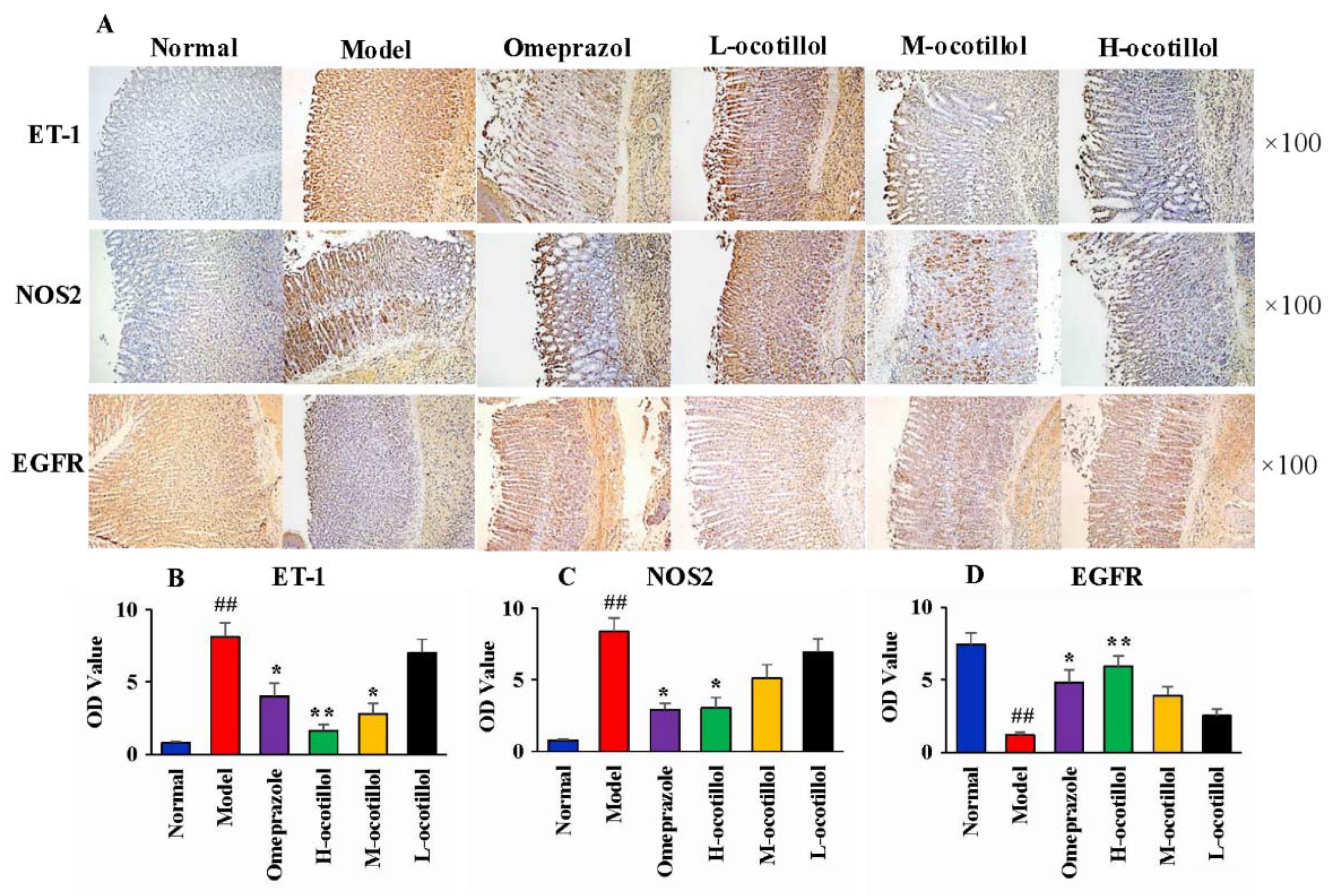
2.1.4. Histological and Immunohistochemical Analysis
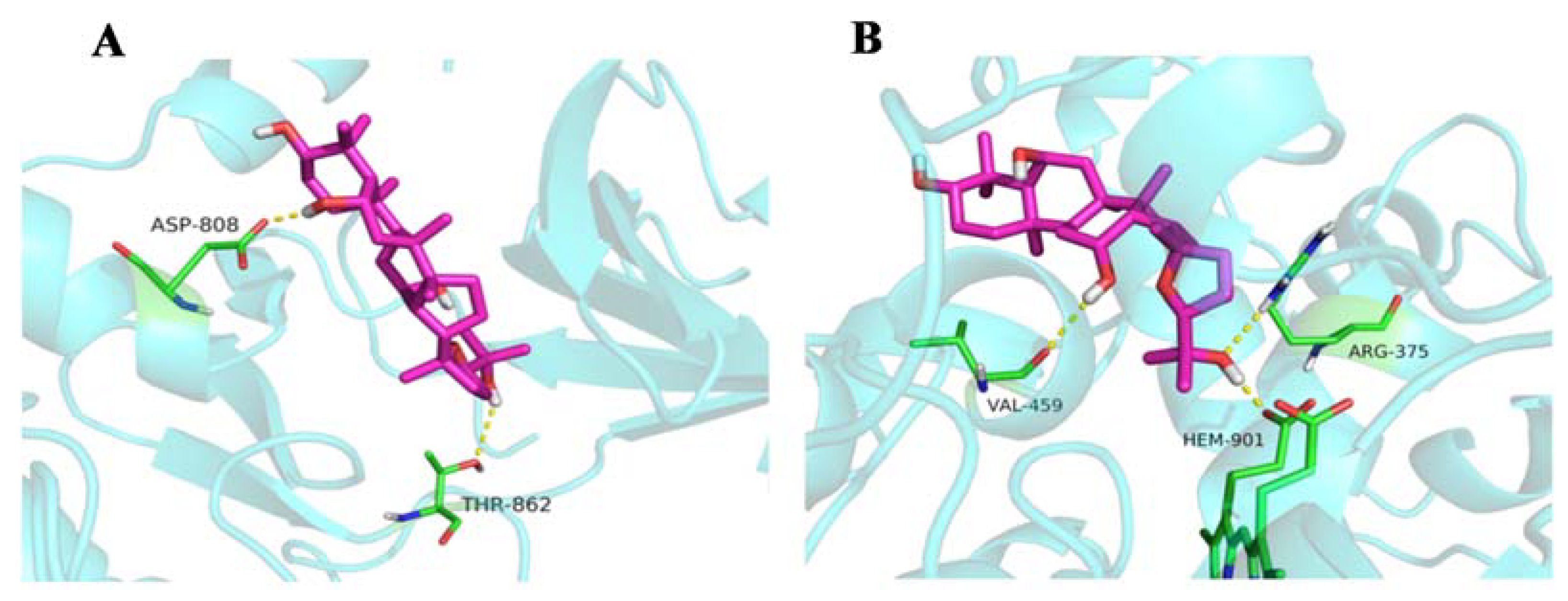
2.1.5. Molecular Docking
2.2. Metabolomics Study
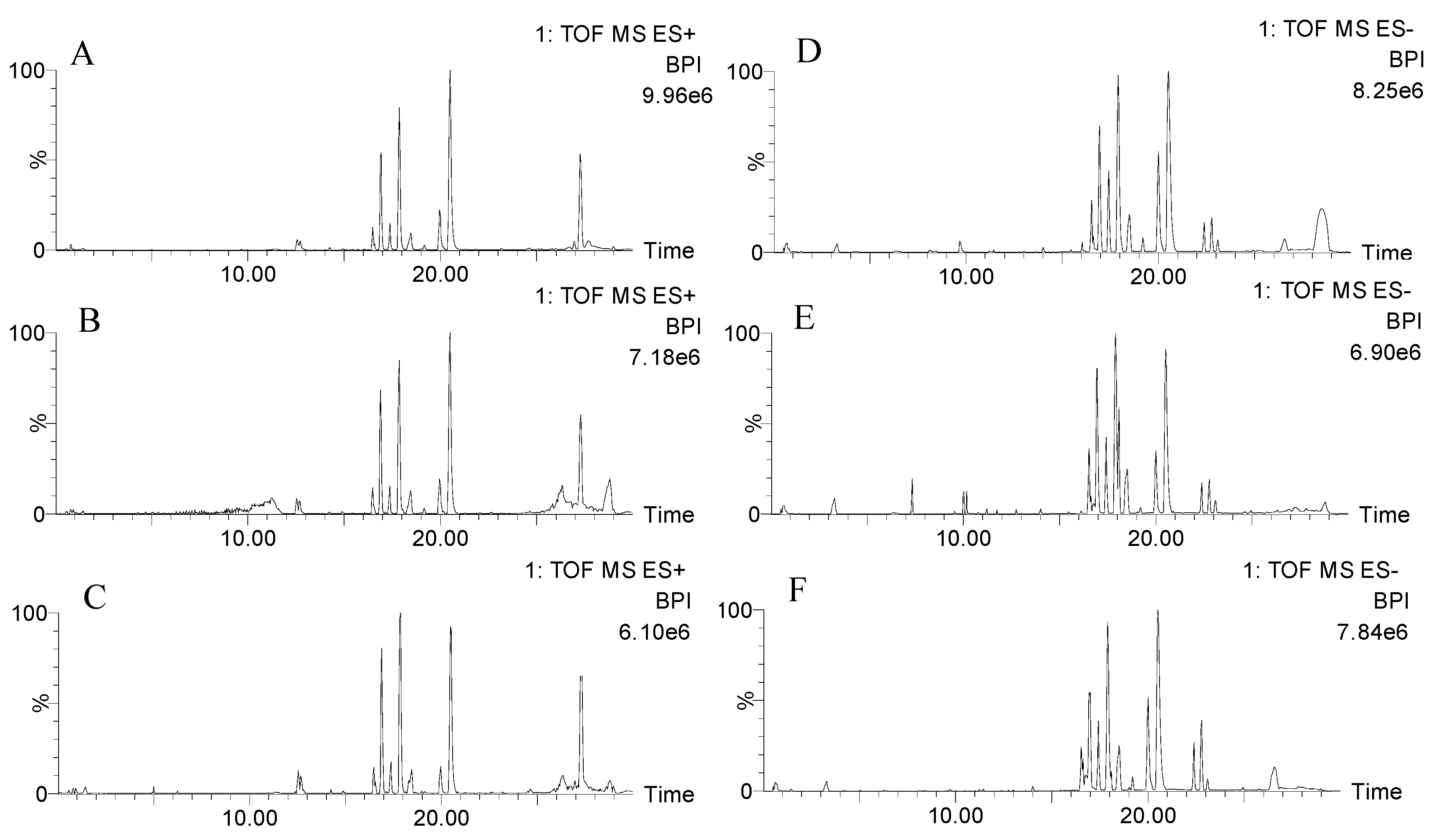
2.2.1. Validation of UPLC-QTOF-MS
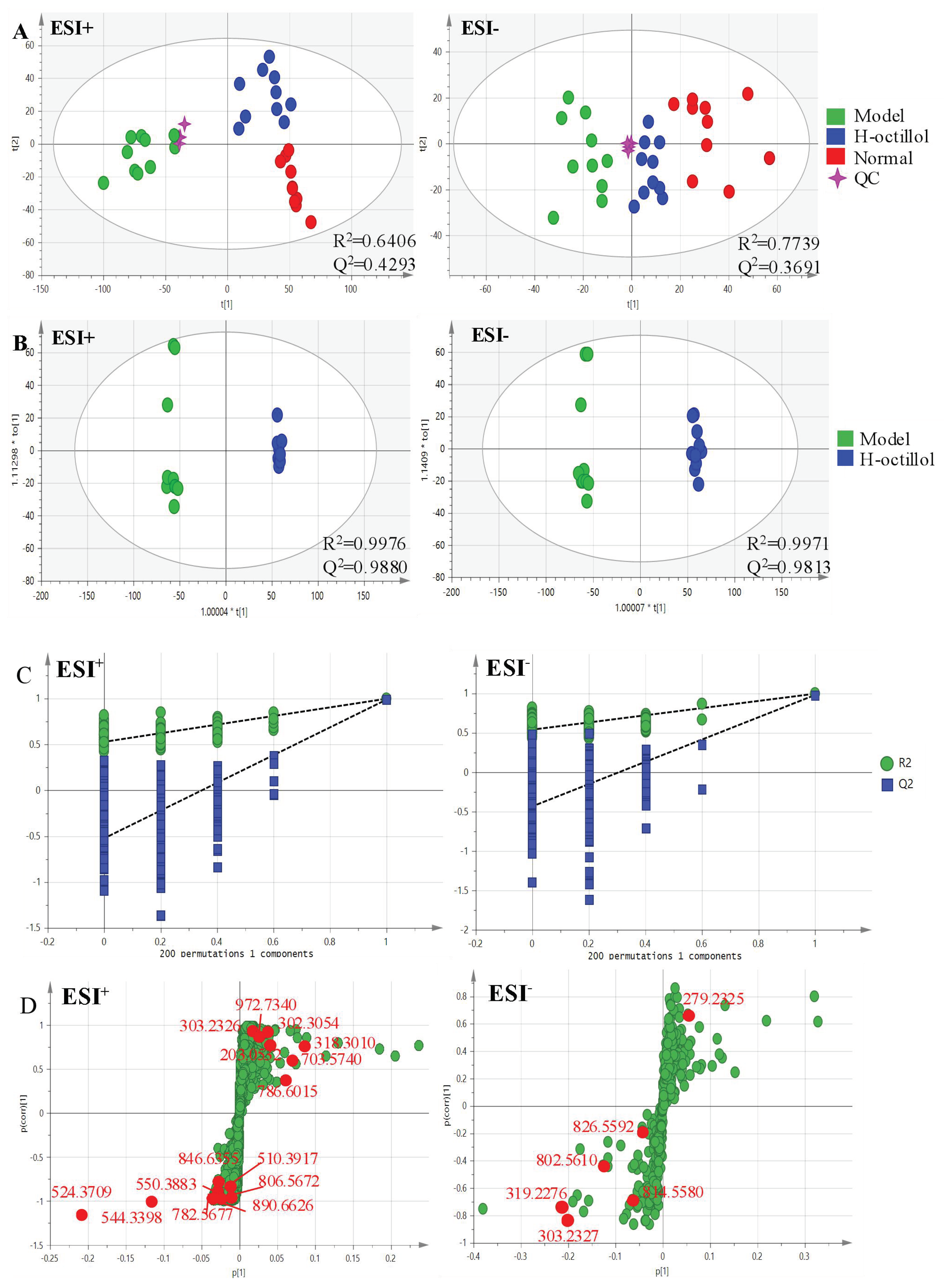
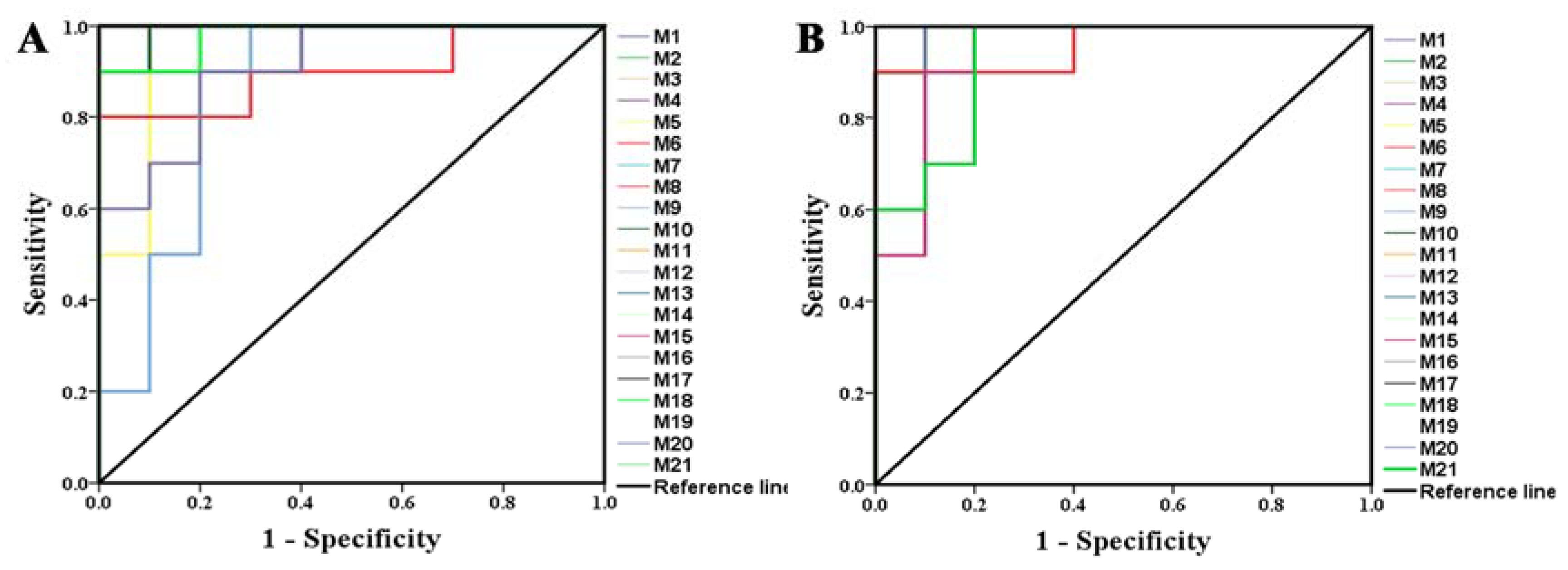
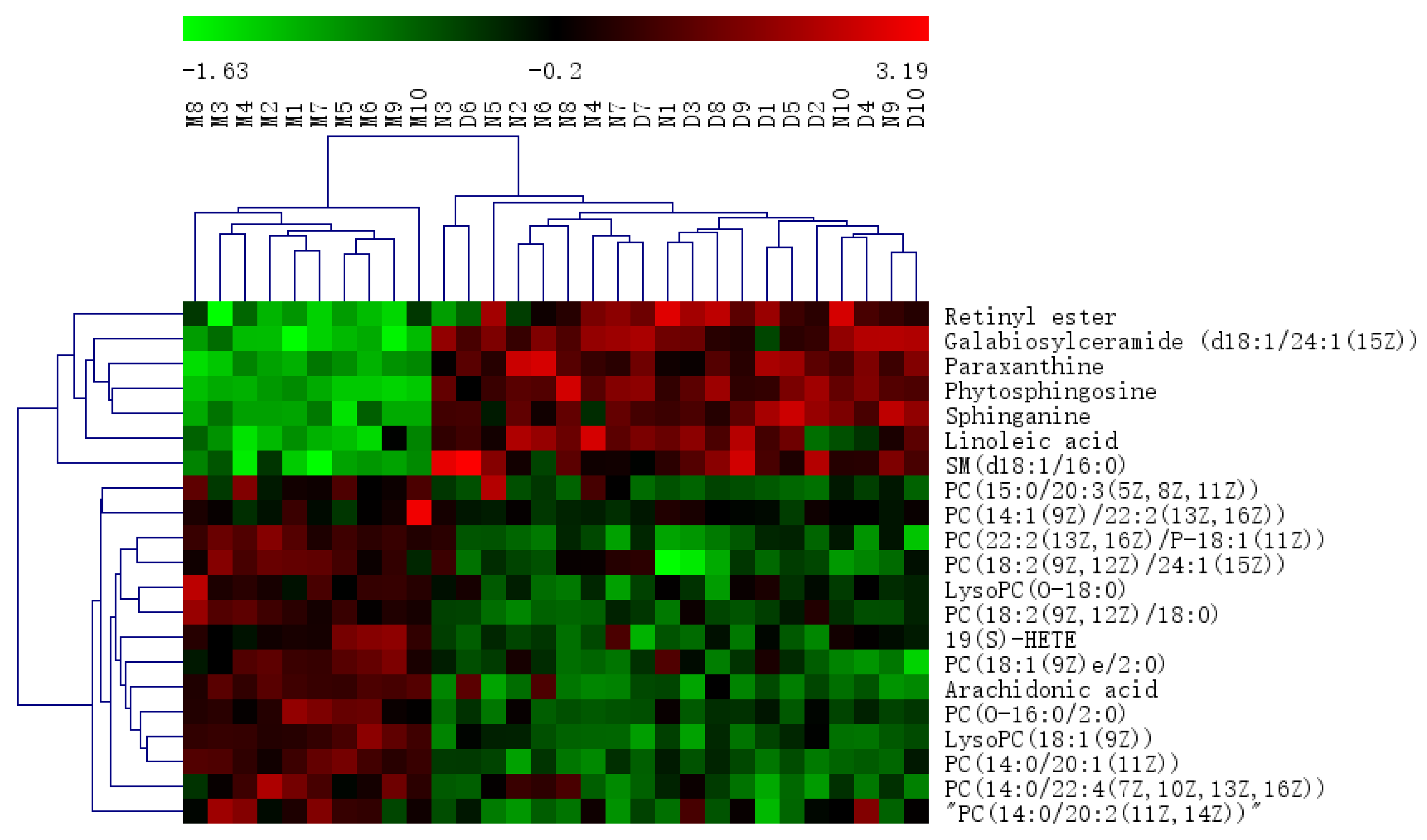
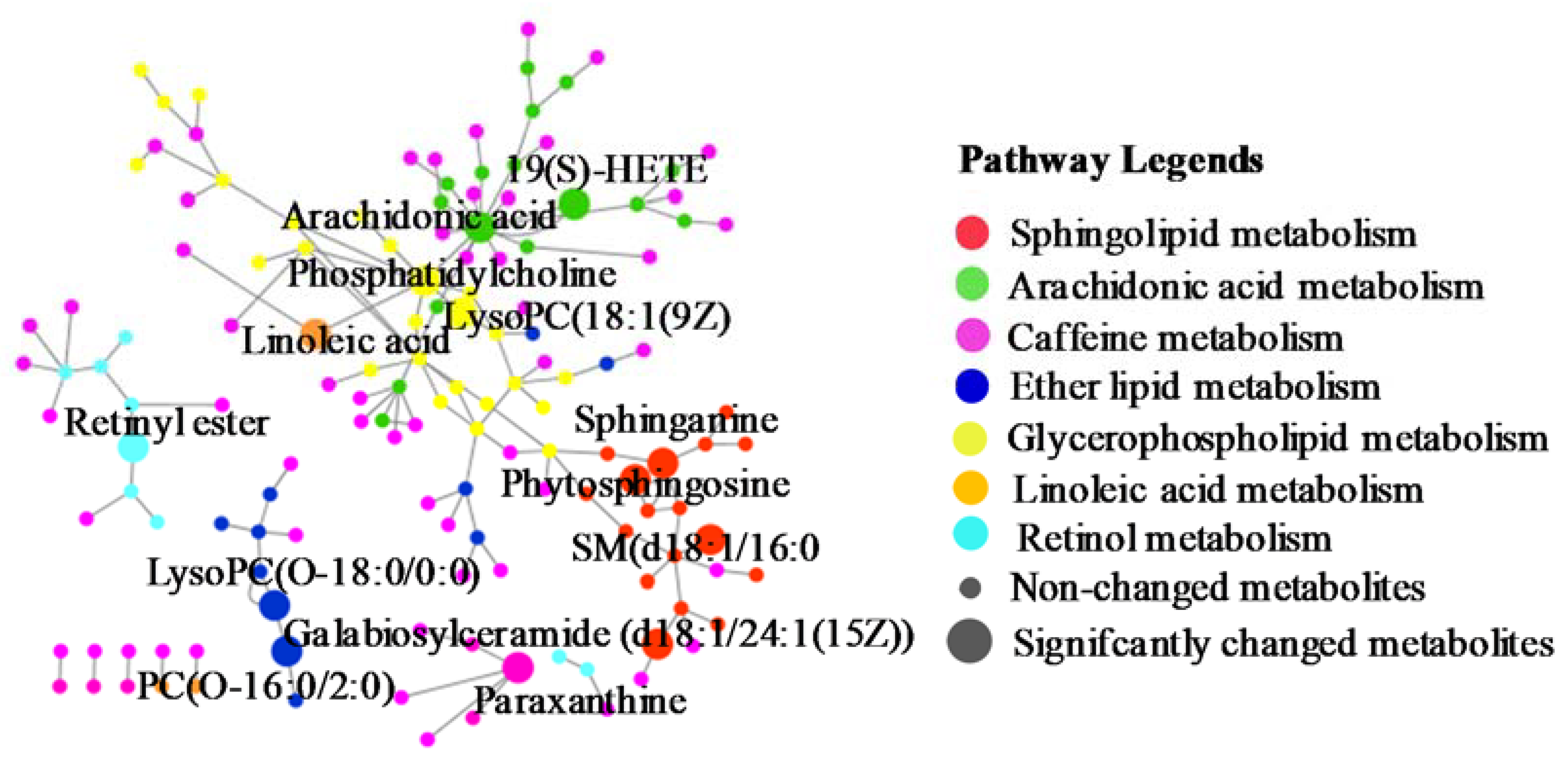
2.2.2. Identification of the Differential Metabolites and Metabolic Pathways
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. UPLC-QTOF-MS Conditions
4.3. Experimental Design
4.4. Preparation of Samples
4.5. Gastroprotective Effects
4.5.1. Body Weights
4.5.2. ET-1 and NO Levels in Serum
4.5.3. EGF, SOD, and NO Levels in Gastric Mucosa
4.5.4. Histological and Immunohistochemical Analysis
4.5.5. Molecular Docking
4.6. Data Analysis
4.7. Metabolomics Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| AUC | Area under curve |
| EGF | Epidermal growth factor |
| EGFR | Epidermal growth factor receptor |
| ELISA | Enzymeelinked immunosorbent assay |
| ESI | Electrospray ionization |
| ET-1 | Endothelin-1 |
| GU | Gastric ulcer |
| H&E | Hematoxylin and eosin |
| HETE | Hydroxyeicosatetraenoic acid |
| H-ocotillol | high-dose ocotillol |
| L-ocotillol | low-dose ocotillol |
| LysoPC | Lysophosphatidylcholine |
| M-ocotillol | moderate-dose ocotillol |
| NOS2 | Inducible nitric oxide synthase |
| OPLS-DA | Orthogonal Projections to Latent Structures Discriminant Analysis |
| PC | Phosphatidylcholine |
| PCA | Principal Component Analysis |
| PDB | Protein Data Bank |
| QC | Quality Control |
| QTOF-MS | Quadrupole Time of Flight-Mass Spectrometry |
| ROC | Receiver operating characteristic |
| RSD | Relative Standard Deviation |
| RT | Retention Time |
| SD | standard deviation |
| SM | Sphingomyelin |
| SOD | Superoxide dismutase |
| UPLC | Ultra-Performance Liquid Chromatography |
| VIP | Variable importance in the projection |
References
- Dong, Z.P.; Zhang, X.; Chen, X.; Zhang, J.Z. Significance of Serological Gastric Biopsy in Different Gastric Mucosal Lesions: An Observational Study. Clin. Lab. 2019, 65, 2141–2148. [Google Scholar] [CrossRef]
- El-Ashmawy, N.E.; Khedr, E.G.; El-Bahrawy, H.A.; Selim, H.M. Nebivolol prevents indomethacin-induced gastric ulcer in rats. J. Immunotoxicol. 2016, 13, 580–589. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.M.; Sun, J.; Niu, C.; Yu, D.D.; Chen, Z.W.; Cong, W.T.; Geng, F.N. Mechanistic evaluation of gastroprotective effects of Kangfuxin on ethanol-induced gastric ulcer in mice. Chem. Biol. Interact. 2017, 273, 115–124. [Google Scholar] [CrossRef]
- Devaraj, V.C.; Krishna, B.G.; Viswanatha, G.L.; Prasad, V.S.; Babu, S.N.V. Protective effect of leaves of Raphinus sativus Linn on experimentally induced gastric ulcers in rats. Saudi Pharm. J. 2011, 19, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, L.M.; Boeing, T.; Somensi, L.B.; Cury, B.J.; Steimbach, V.M.; Silveria, A.C.; Niero, R.; Cechinel, V.; Santin, J.R.; de Andrade, S.F. Evidence of gastric ulcer healing activity of Maytenus robusta Reissek: In vitro and in vivo studies. J. Ethnopharmacol. 2015, 175, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.G.; More, S.K.; Vomhof-DeKrey, E.E.; Golovko, M.Y.; Basson, M.D. Small molecule fAK activator promotes human intestinal epithelial monolayer wound closure and mouse ulcer healing. Sci. Rep. 2019, 9, 14669. [Google Scholar] [CrossRef] [Green Version]
- Matthis, A.L.; Kaji, I.; Engevik, K.A.; Akiba, Y.; Kaunitz, J.D.; Montrose, M.H.; Aihara, E. Defcient Active Transport Activity in Healing Mucosa After Mild Gastric Epithelial Damage. Digest. Dis. Sci. 2019, 65, 119–131. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Y.; Wang, C.Z.; Li, J.N.; Xiong, L.X.; Wang, Z.Z.; Liu, J.P.; Li, P.Y. Evaluation of the gastroprotective effects of 20 (S)-ginsenoside Rg3 on gastric ulcer models in mice. J. Gins Res. 2019, 43, 550–561. [Google Scholar] [CrossRef]
- Kangwan, N.; Park, J.M.; Kim, E.H.; Hahm, K.B. Quality of healing of gastric ulcers: Natural products beyond acid suppression. World J. Gastrointest. Pathophysiol. 2014, 5, 40–47. [Google Scholar] [CrossRef]
- Huong, N.T.; Matsumoto, K.; Watanabe, H. The antistress effect of majonoside—R2, a major saponin component of Vietnamese ginseng: Neuronal mechanisms of action. Methods Find. Exp. Clin. Pharmacol. 1998, 20, 65–76. [Google Scholar] [CrossRef]
- Wang, X.X.; Wang, C.M.; Wang, J.M.; Zhao, S.Q.; Zhang, K.; Wang, J.M.; Zhang, W.; Wu, C.F.; Yang, J.Y. Pseudoginsenoside-F11 (PF11) exerts anti-neuroinflammatory effects on LPS-activated microglial cells by inhibiting TLR4-mediated TAK1/IKK/NF-κB, MAPKs and Akt signaling pathways. Neuropharmacology 2014, 79, 642–656. [Google Scholar] [CrossRef]
- Wang, P.W.; Hou, Y.; Zhang, W.; Zhang, H.T.; Che, X.H.; Gao, Y.F.; Liu, Y.L.; Yang, D.P.; Wang, J.M.; Xiang, R.W.; et al. Pseudoginsenoside-F11 Attenuates Lipopolysaccharide-Induced Acute Lung Injury by Suppressing Neutrophil Infiltration and Accelerating Neutrophil Clearance. Inflammation 2019, 42, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, H.L.; Yang, J.Y.; Xie, J.; Xu, J.Y.; Liu, C.; Wu, C.F. Pseudoginsenoside-F11 attenuates cognitive impairment by ameliorating oxidative stress and neuroinflammation in D-galactose-treated mice. Int. Immunopharmacol. 2019, 67, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H. Gut microbiota-mediated pharmacokinetics of ginseng saponins. J. Gins. Res. 2018, 42, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Jeong, J.J.; Van Le, T.H.; Eun, S.H.; Nguyen, M.D.; Park, J.H.; Kim, D.H. Ocotillol, a Majonoside R2 Metabolite, Ameliorates 2,4,6- Trinitrobenzenesulfonic Acid-Induced Colitis in Mice by Restoring the Balance of Th17/Treg Cells. J. Agric. Food Chem. 2015, 63, 7024–7031. [Google Scholar] [CrossRef]
- Wang, H.B.; Yu, P.F.; Bai, J.; Zhang, J.Q.; Kong, L.; Zhang, F.X.; Du, G.Y.; Pei, S.Q.; Zhang, L.X.; Jiang, Y.T.; et al. Ocotillol Enhanced the Antitumor Activity of Doxorubicin via p53-Dependent Apoptosis. Evid. Based Complement. Alternat. Med. 2013, 2013, 468537. [Google Scholar] [CrossRef]
- Yoon, D.; Lee, M.J.; Kim, S.; Kim, S. Applications of NMR spectroscopy based metabolomics: A review. J. Korean Magn. Reso. Soc. 2013, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Cao, C.; Chen, S.; Hu, L.Y.; Bao, W.; Shi, H.D.; Zhao, X.J.; Sun, C.H. Serum Metabolomics Analysis of Quercetin against Acrylamide-Induced Toxicity in Rats. J. Agric. Food Chem. 2016, 64, 9237–9245. [Google Scholar] [CrossRef]
- Zhang, L.M.; Dong, M.Y.; Xu, G.Y.; Tian, Y.; Tang, H.R.; Wang, Y.L. Metabolomics Reveals that Dietary Ferulic Acid and Quercetin Modulate Metabolic Homeostasis in Rats. J. Agric. Food Chem. 2018, 66, 1723–1731. [Google Scholar] [CrossRef]
- Wang, C.Z.; Lin, H.Q.; Yang, N.; Wang, H.; Zhao, Y.; Li, P.Y.; Liu, J.P.; Wang, F. Effects of Platycodins Folium on Depression in Mice Based on a UPLC-Q/ TOF-MS Serum Assay and Hippocampus Metabolomics. Molecules 2019, 24, 1712. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Liu, Y.; Li, W.; Wang, Z.; Guo, P.; Li, L.; Li, N.J. Metabolic profiling of the effects of ginsenoside Re in an Alzheimer’s disease mouse model. Behav. Brain Res. 2018, 337, 160–172. [Google Scholar] [CrossRef]
- Liu, G.; Lee, D.P.; Schmidt, E.; Prasad, G.L. Pathway Analysis of Global Metabolomic Profles Identifed Enrichment of Caffeine, Energy, and Arginine Metabolism in Smokers but Not Moist Snuff Consumers. Bioinform. Biol. Insights 2019, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bahr, T.M.; Hughes, G.J.; Armstrong, M.; Reisdorph, R.; Coldren, C.D.; Edwards, M.G.; Schnell, C.; Kedl, R.; LaFlamme, D.J.; Reisdorph, N.; et al. Peripheral Blood Mononuclear Cell Gene Expression in Chronic Obstructive Pulmonary Disease. Am. J. Resp. Cell Mol. 2013, 49, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Li, C.H.; Wu, Q.Q.; Yao, Z.M.; Wu, J.; Huang, P.; Wang, D.L.; Li, Z.G. Metabolic profiling of chronic obstructive pulmonary disease model rats and the interventional effects of HuaTanJiangQi decoction using UHPLC-Q-TOF/MSE. J. Pharm. Biomed. Anal. 2020, 180, 113078. [Google Scholar] [CrossRef]
- Ahmad, M.I.; Ijaz, M.U.; Hussain, M.; Khan, I.A.; Mehmood, N.; Siddiqi, S.M.; Liu, C.C.; Zhao, D.; Xu, X.L.; Zhou, G.H.; et al. High fat diet incorporated with meat proteins changes biomarkers of lipid metabolism, antioxidant activities, and the serum metabolomic profile in Glrx1−/− mice†. Food Funct. 2020, 11, 236–252. [Google Scholar] [CrossRef]
- Cheng, M.M.; Wu, H.; Wu, H.F.; Liu, X.C.; Zhou, A. Metabolic profiling of copper-laden hepatolenticular degeneration model rats and the interventional effects of Gandou decoction using UPLC-Q-TOF/MS. J. Pharm. Biomed. Anal. 2019, 164, 187–195. [Google Scholar] [CrossRef]
- Zhou, X.S.; Wang, R.P.; Zhang, T.; Liu, F.; Zhang, W.; Wang, G.L.; Gu, G.R.; Han, Q.Q.; Xu, D.M.; Yao, C.L.; et al. Identification of Lysophosphatidylcholines and Sphingolipids as Potential Biomarkers for Acute Aortic Dissection via Serum Metabolomics. Eur. J. Vasc. Endovasc. Surg. 2019, 57, 434–441. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.; Hoogerland, J.A.; Wessel, H.; Heegsma, J.; Derks, T.G.J.; Veer, E.V.; Mithieux, G.; Rajas, F.; Oosterveer, M.H.; Faber, K.N. Glycogen storage disease type 1a is associated with disturbed vitamin A metabolism and elevated serum retinol levels. Hum. Mol. Genet. 2020, 29, 264–273. [Google Scholar] [CrossRef]
- Wang, T.Q.; Fu, X.J.; Chen, Q.F.; Patra, J.K.; Wang, D.D.; Wang, Z.G.; Gai, Z.B. Arachidonic Acid Metabolism and Kidney Inflammation. Int. J. Mol. Sci. 2019, 20, 3683. [Google Scholar] [CrossRef] [Green Version]
- Xia, F.B.; Li, A.; Chai, Y.S.; Xiao, X.; Wan, J.B.; Li, P.; Wang, Y.T. UPLC/Q-TOFMS-Based Metabolomics Approach to Reveal the Protective Role of Other Herbs in An-Gong-Niu-Huang Wan Against the Hepatorenal Toxicity of Cinnabar and Realgar. Front. Pharmacol. 2018, 9, 618. [Google Scholar] [CrossRef]
- Wang, D.; Li, R.S.; Wei, S.Z.; Gao, S.J.; Xu, Z.; Liu, H.H.; Wang, R.L.; Li, H.T.; Cai, H.D.; Wang, J.; et al. Metabolomics combined with network pharmacology exploration reveals the modulatory properties of Astragali Radix extract in the treatment of liver fibrosis. Chin. Med. 2019, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Z.; Hu, H.; Li, Z.G.; Li, R.H.; Xu, F.; Zhao, C.Y.; An, Y.; Liu, Y.G.; Wang, Z.L.; Zhang, B.B.; et al. Pharmacokinetic Characterizations of Ginsenoside Ocotillol, RT5 and F11, the Promising Agents for Alzheimer’s Disease from American Ginseng, in Rats and Beagle Dogs. Pharmacology 2019, 104, 7–20. [Google Scholar] [CrossRef]
- Fu, K.Q.; Lin, H.Y.; Miyamoto, Y.; Wu, C.F.; Yang, J.Y.; Uno, K.; Nitta, A. Pseudoginsenoside-F11 inhibits methamphetamine-induced behaviors by regulating dopaminergic and GABAergic neurons in the nucleus accumbens. Psychopharmacology 2016, 233, 831–840. [Google Scholar] [CrossRef]
- Wang, Z.J.; Sun, L.; Peng, W.; Ma, S.; Zhu, C.; Fu, F.; Heinbockel, T. Ginseng derivative ocotillol enhances neuronal activity through increased glutamate release: A possible mechanism underlying increased spontaneous locomotor activity of mice. Neuroscience 2011, 195, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Shi, N.; Shi, H.L.; Ye, H.; Li, N.; Sun, P.; Bai, D.F.; Yuan, H.P. Correlations of Acute Cerebral Hemorrhage Complicated with Stress Ulcer Bleeding with Acute Physiology and Chronic Health Evalua (APACHE) II Score, Endothelin (ET), Tumor Necrosis Factor-alpha (TNF-a), and Blood Lipids. Med. Sci. Monit. 2018, 24, 9120–9126. [Google Scholar] [CrossRef]
- Romana-Souza, B.; Santos, J.S.; Bandeira, L.G.; Monte-Alto-Costa, A. Monte-Alto-Costa, Selective inhibition of COX-2 improves cutaneous wound healing of pressure ulcers in mice through reduction of iNOS expression. Life Sci. 2016, 153, 82–92. [Google Scholar] [CrossRef]
- Schanuel, F.S.; Saguie, B.O.; Monte-Alto-Costa, A. Olive oil promotes wound healing of mice pressure injuries through NOS-2 and Nrf2. Appl. Physiol. Nutr. Metab. 2019, 44, 1199–1208. [Google Scholar] [CrossRef]
- Suo, H.Y.; Zhao, X.; Qian, Y.; Sun, P.; Zhu, K.; Li, J.; Sun, B.Z. Lactobacillus fermentum Suo Attenuates HCl/Ethanol Induced Gastric Injury in Mice through Its Antioxidant Effects. Nutrients 2016, 8, 155. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Bao, Y.R.; Li, T.J.; Chang, X.; Yang, G.L.; Meng, X.S. Multipathway integrated adjustment mechanism of Glycyrrhiza triterpenes curing gastric ulcer in rats. Pharmacogn. Mag. 2017, 13, 209–215. [Google Scholar]
- Tunaru, S.; Chennupati, R.; Nusing, R.M.; Offermanns, S. Arachidonic Acid Metabolite 19(S)-HETE Induces Vasorelaxation and Platelet Inhibition by Activating Prostacyclin (IP) Receptor. PLoS ONE 2016, 11, e0163633. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Li, X.Q.; Dai, Y.J.; Wang, Q.H.; Wang, G.Y.; Liu, D.P.; Gu, X.Z.; Yu, D.R.; Ma, Y.L.; et al. Treatment Mechanism of Gardeniae Fructus and Its Carbonized Product Against Ethanol-Induced Gastric Lesions in Rats. Front. Pharmacol. 2019, 10, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maksem, J.; Jacobson, N.; Neiderhiser, D.H. Lysophosphatidylcholine-induced gastric injury and ulceration in the guinea pig. Am. J. Pathol. 1984, 115, 288–295. [Google Scholar] [PubMed]
- Tarnawski, A.; Hollander, D.; Gergely, H. Protection of the gastric mucosa by linoleic acid-a nutrient essential fatty acid. Med. Clin. Exp. 1987, 10, 132–135. [Google Scholar]
- Schepp, W.; Steffen, B.; Ruoff, H.J.; Schusdziarra, V.; Classen, M. Modulation of rat gastric mucosal prostaglandin E2 release by dietary linoleic acid: Effects on gastric acid secretion and stress-induced mucosal damage. Gastroenterology 1988, 95, 18–25. [Google Scholar] [CrossRef]
- Baek, S.W.; Kim, N.K.; Jin, H.J.; Koh, C.W.; Kim, C.K.; Kwon, O.H.; Kim, J.S.; Cho, M.H.; Park, C.K. Anti-ulcer Actions of Phytosphingosine Hydrochloride in Different Experimental Rat Ulcer Models. Arzneimittelforsch 2005, 55, 461–465. [Google Scholar] [CrossRef]
- Kanfer, J.N.; Young, O.M.; Shapiro, D.; Brady, R.O. The metabolism of sphingomyelin. I. Purification and properties of a sphingomyelin-cleaving enzyme from rat liver tissue. J. Biol. Chem. 1966, 241, 1081–1084. [Google Scholar]
- Nakashita, M.; Suzuki, H.; Miura, S.; Taki, T.; Uehara, K.; Mizushima, T.; Nagata, H.; Hibi, T. Attenuation of Acetic Acid-Induced Gastric Ulcer Formation in Rats by Glucosylceramide Synthase Inhibitors. Digest. Dis. Sci. 2013, 58, 354–362. [Google Scholar] [CrossRef]
- Harrison, E.H. Mechanisms of Transport and Delivery of Vitamin A and Carotenoids to the Retinal Pigment Epithelium. Mol. Nutr. Food Res. 2019, 63, 1801046. [Google Scholar] [CrossRef] [Green Version]
- Rosa, C.A.; Paggiaro, A.O.; Carvalho, V.F. Effect of Hydrogel Enriched with Alginate, Fatty Acids, and Vitamins A and E on Pressure Injuries: A Case Series. Plast. Surg. Nurs. 2019, 39, 87–94. [Google Scholar] [CrossRef]
- Kafi, R.; Kwak, H.S.R.; Schumacher, W.E.; Cho, S.Y.; Hanft, V.N.; Hamilton, T.A.; King, A.L.; Neal, J.D.; Varani, J.; Fisher, G.J.; et al. Improvement of Naturally Aged Skin with Vitamin A (Retinol). Arch. Dermatol. 2007, 143, 606–612. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Pi, Z.F.; Men, L.H.; Chen, W.J.; Liu, Z.Q.; Liu, Z.Y. Urinary metabonomic study of Panax ginseng in deficiency of vital energy rat using ultra performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. J. Ethnopharmacol. 2016, 184, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.Y.; Yao, T.; Zhang, B.Y.; Li, Y.; Kang, N.; Cao, S.J.; Ding, L.Q.; Chen, L.X.; Qiu, F. Withapubesides A-D: Natural inducible nitric oxide synthase (iNOS) inhibitors from Physalis pubescens. Org. Biomol. Chem. 2017, 15, 10016–10023. [Google Scholar] [CrossRef] [PubMed]
- Garcin, E.D.; Arvai, A.S.; Rosenfeld, R.J.; Kroeger, M.D.; Crane, B.R.; Andersson, G.; Andrews, G.; Hamley, P.J.; Mallinder, P.R.; Nicholls, D.J.; et al. Anchored plasticity opens doors for selective inhibitor design in nitric oxide synthase. Nat. Chem. Biol. 2008, 4, 700–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, T.; Seto, M.; Banno, H.; Kawakita, Y.; Oorui, M.; Taniguchi, T.; Ohta, Y.; Tamura, T.; Nakayama, A.; Miki, H.; et al. Design and synthesis of novel human epidermal growth factor receptor 2 (HER2)/epidermal growth factor receptor (EGFR) dual inhibitors bearing a Pyrrolo 3,2-d pyrimidine Scaffold. J. Med. Chem. 2011, 54, 8030–8050. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT | Mass | Compound Name | VIP | Formula | Adducts | Δm | HMDB ID | Pathways | Content Level |
|---|---|---|---|---|---|---|---|---|---|---|
| 1* | 0.60 | 203.0532 | Paraxanthine | 2.77 | C7H8N4O2 | M+Na | 0 | HMDB0001860 | CM [22] | CM < CD |
| 2a | 11.39 | 972.7340 | Galabiosylceramide (d18:1/24:1(15Z)) | 1.53 | C54H101NO13 | M+H | −1.13 | HMDB0004837 | SphM [23] | CM < CD |
| 3a | 11.40 | 846.6355 | PC(22:2(13Z,16Z)/P-18:1(11Z)) | 2.62 | C48H90NO7P | M+Na | 0.24 | HMDB0008621 | AM, LM, GlyM [24,25,26] | CD < CM |
| 4a | 11.49 | 890.6626 | PC(18:2(9Z,12Z)/24:1(15Z)) | 1.99 | C50H94NO8P | M+Na | 1.24 | HMDB0008158 | AM, LM,GlyM [24,25,26] | CD < CM |
| 5* | 12.72 | 318.3010 | Phytosphingosine | 6.98 | C18H39NO3 | M+H | 0.63 | HMDB0004610 | SphM [25,27] | CM<CD |
| 6* | 14.94 | 302.3054 | Sphinganine | 2.95 | C18H39NO2 | M+H | −1.65 | HMDB0000269 | SphM [25,27] | CM<CD |
| 7a | 16.90 | 544.3398 | LysoPC(18:1(9Z)) | 7.92 | C26H52NO7P | M+Na | 3.49 | HMDB0002815 | GlyM [24,25] | CD<CM |
| 8a | 18.04 | 303.2326 | Retinyl ester | 1.91 | C20H30O2 | M+H | 0.66 | HMDB0003598 | RM [28] | CM<CD |
| 9a | 18.07 | 814.5580 | PC(15:0/20:3(5Z,8Z,11Z)) | 1.09 | C43H80NO8P | M+FA-H | −2.21 | HMDB0007947 | AM, LM,GlyM [24,25,26] | CD<CM |
| 10* | 18.11 | 319.2276 | 19(S)-HETE | 7.39 | C20H32O3 | M-H | 0.94 | HMDB0011136 | AM [29] | CD<CM |
| 11a | 20.54 | 524.3709 | PC(O-16:0/2:0) | 18.53 | C26H54NO7P | M+H | −1.33 | HMDB0062195 | EM [30] | CD<CM |
| 12a | 20.90 | 550.3883 | PC(18:1(9Z)e/2:0) | 2.10 | C28H56NO7P | M+H | 1.82 | HMDB0011148 | AM, LM,GlyM [24,25,26] | CD<CM |
| 13a | 21.15 | 510.3917 | LysoPC(O-18:0/0:0) | 1.76 | C26H56NO6P | M+H | −1.37 | HMDB0011149 | EM [31] | CD<CM |
| 14* | 22.81 | 303.2327 | Arachidonic acid | 7.16 | C20H32O2 | M-H | 0.99 | HMDB0001043 | AM [24] | CD<CM |
| 15* | 23.13 | 279.2325 | Linoleic acid | 1.40 | C18H32O2 | M-H | 0.36 | HMDB0000673 | LM [28] | CM<CD |
| 16a | 24.68 | 806.5672 | PC(14:1(9Z)/22:2(13Z,16Z)) | 1.87 | C44H82NO8P | M+Na | −0.50 | HMDB0007921 | AM, LM,GlyM [24,25,26] | CD<CM |
| 17a | 26.33 | 786.6015 | PC(18:2(9Z,12Z)/18:0) | 5.38 | C44H84NO8P | M+H | 0.25 | HMDB0008135 | AM, LM,GlyM [24,25,26] | CM<CD |
| 18a | 27.41 | 703.5740 | SM(d18:1/16:0) | 6.72 | C39H80N2O6P | M+H | −1.99 | HMDB0010169 | SphM [27] | CM<CD |
| 19a | 27.54 | 782.5677 | PC(14:0/20:1(11Z)) | 3.68 | C42H82NO8P | M+Na | 0.13 | HMDB0007879 | AM, LM,GlyM [24,25,26] | CD<CM |
| 20a | 27.67 | 826.5592 | PC(14:0/22:4(7Z,10Z,13Z,16Z)) | 1.50 | C44H80NO8P | M+FA-H | −0.73 | HMDB0007889 | AM, LM,GlyM [24,25,26] | CD<CM |
| 21a | 27.91 | 802.5610 | PC(14:0/20:2(11Z,14Z)) | 2.76 | C42H80NO8P | M+FA-H | 1.50 | HMDB0007880 | AM, LM,GlyM [24,25,26] | CD<CM |
| No. | M and N | M and Ocotillol | ||
|---|---|---|---|---|
| AUC | p | AUC | p | |
| 1 | 1.000 | <0.001 | 1.000 | <0.001 |
| 2 | 1.000 | <0.001 | 1.000 | <0.001 |
| 3 | 1.000 | <0.001 | 1.000 | <0.001 |
| 4 | 0.960 | 0.0001 | 0.990 | <0.001 |
| 5 | 1.000 | <0.001 | 1.000 | <0.001 |
| 6 | 1.000 | <0.001 | 1.000 | <0.001 |
| 7 | 1.000 | <0.001 | 1.000 | <0.001 |
| 8 | 0.900 | 0.002 | 0.960 | 0.001 |
| 9 | 0.860 | 0.007 | 1.000 | <0.001 |
| 10 | 0.990 | <0.001 | 0.990 | <0.001 |
| 11 | 1.000 | <0.001 | 1.000 | <0.001 |
| 12 | 0.980 | <0.001 | 0.990 | <0.001 |
| 13 | 1.000 | <0.001 | 1.000 | <0.001 |
| 14 | 1.000 | <0.001 | 1.000 | <0.001 |
| 15 | 0.950 | 0.001 | 0.940 | 0.001 |
| 16 | 1.000 | <0.001 | 1.000 | <0.001 |
| 17 | 1.000 | <0.001 | 1.000 | <0.001 |
| 18 | 0.980 | <0.001 | 0.990 | <0.001 |
| 19 | 1.000 | <0.001 | 1.000 | <0.001 |
| 20 | 0.910 | 0.002 | 0.990 | <0.001 |
| 21 | 0.980 | <0.001 | 0.930 | 0.001 |
| Pathway Name | Match Status | p | −log (p) | Holm p | FDR | Impact |
|---|---|---|---|---|---|---|
| Sphingolipid metabolism (SphM) | 4/21 | 1.8325 × 10−5 | 10.9070 | 0.0015 | 0.0015 | 0.1968 |
| Linoleic acid metabolism (LM) | 2/5 | 6.7558 × 10−4 | 7.2999 | 0.0560 | 0.0284 | 1.0000 |
| Arachidonic acid metabolism (AM) | 3/36 | 0.0030 | 5.7997 | 0.2484 | 0.0848 | 0.3329 |
| Ether lipid metabolism (EM) | 2/20 | 0.0119 | 4.4286 | 0.9664 | 0.2506 | 0.2289 |
| Glycerophospholipid metabolism (GlyM) | 2/36 | 0.0366 | 3.3077 | 1.0000 | 0.5124 | 0.1118 |
| Caffeine metabolism (CM) | 1/12 | 0.0990 | 2.3130 | 1.0000 | 1.0000 | 0.6923 |
| Retinol metabolism (RM) | 1/16 | 0.1299 | 2.0411 | 1.0000 | 1.0000 | 0.1617 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Yuan, Y.; Pan, H.; Hsu, A.C.-Y.; Chen, J.; Liu, J.; Li, P.; Wang, F. Protective Effect of Ocotillol, the Derivate of Ocotillol-Type Saponins in Panax Genus, against Acetic Acid-Induced Gastric Ulcer in Rats Based on Untargeted Metabolomics. Int. J. Mol. Sci. 2020, 21, 2577. https://doi.org/10.3390/ijms21072577
Wang C, Yuan Y, Pan H, Hsu AC-Y, Chen J, Liu J, Li P, Wang F. Protective Effect of Ocotillol, the Derivate of Ocotillol-Type Saponins in Panax Genus, against Acetic Acid-Induced Gastric Ulcer in Rats Based on Untargeted Metabolomics. International Journal of Molecular Sciences. 2020; 21(7):2577. https://doi.org/10.3390/ijms21072577
Chicago/Turabian StyleWang, Cuizhu, Yuze Yuan, He Pan, Alan Chen-Yu Hsu, Jinluan Chen, Jinping Liu, Pingya Li, and Fang Wang. 2020. "Protective Effect of Ocotillol, the Derivate of Ocotillol-Type Saponins in Panax Genus, against Acetic Acid-Induced Gastric Ulcer in Rats Based on Untargeted Metabolomics" International Journal of Molecular Sciences 21, no. 7: 2577. https://doi.org/10.3390/ijms21072577