Heat Shock Protein 27 Injection Leads to Caspase Activation in the Visual Pathway and Retinal T-Cell Response

 , ,
, ,
Abstract
:1. Introduction
2. Results
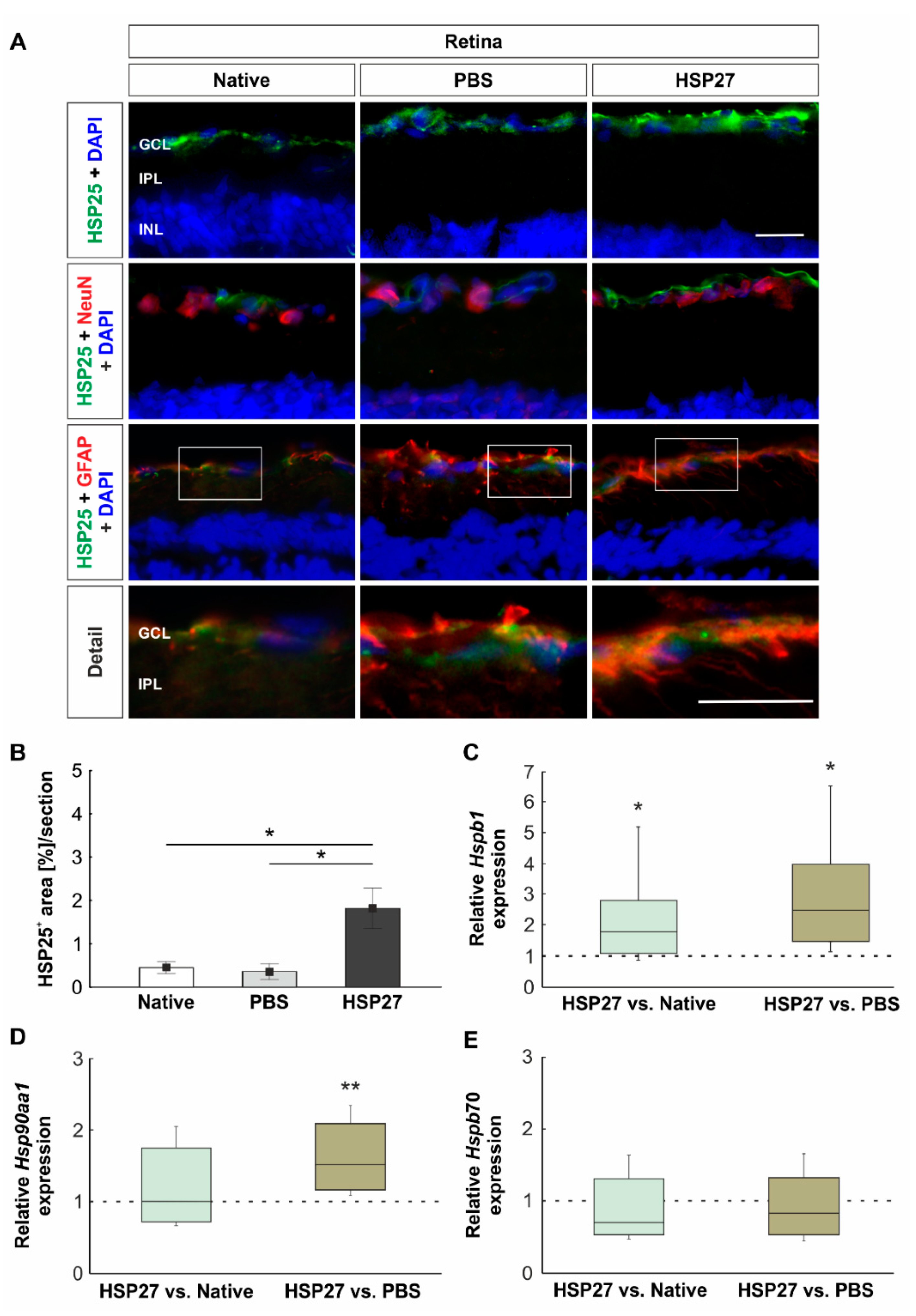
2.1. Increased Heat Shock Protein Levels on Protein and Gene Level
2.2. Unaltered Number of Retinal Ganglion Cells but Increased Apoptosis Rate
2.3. HSP27 Could Activate the NFκB Signal Pathway
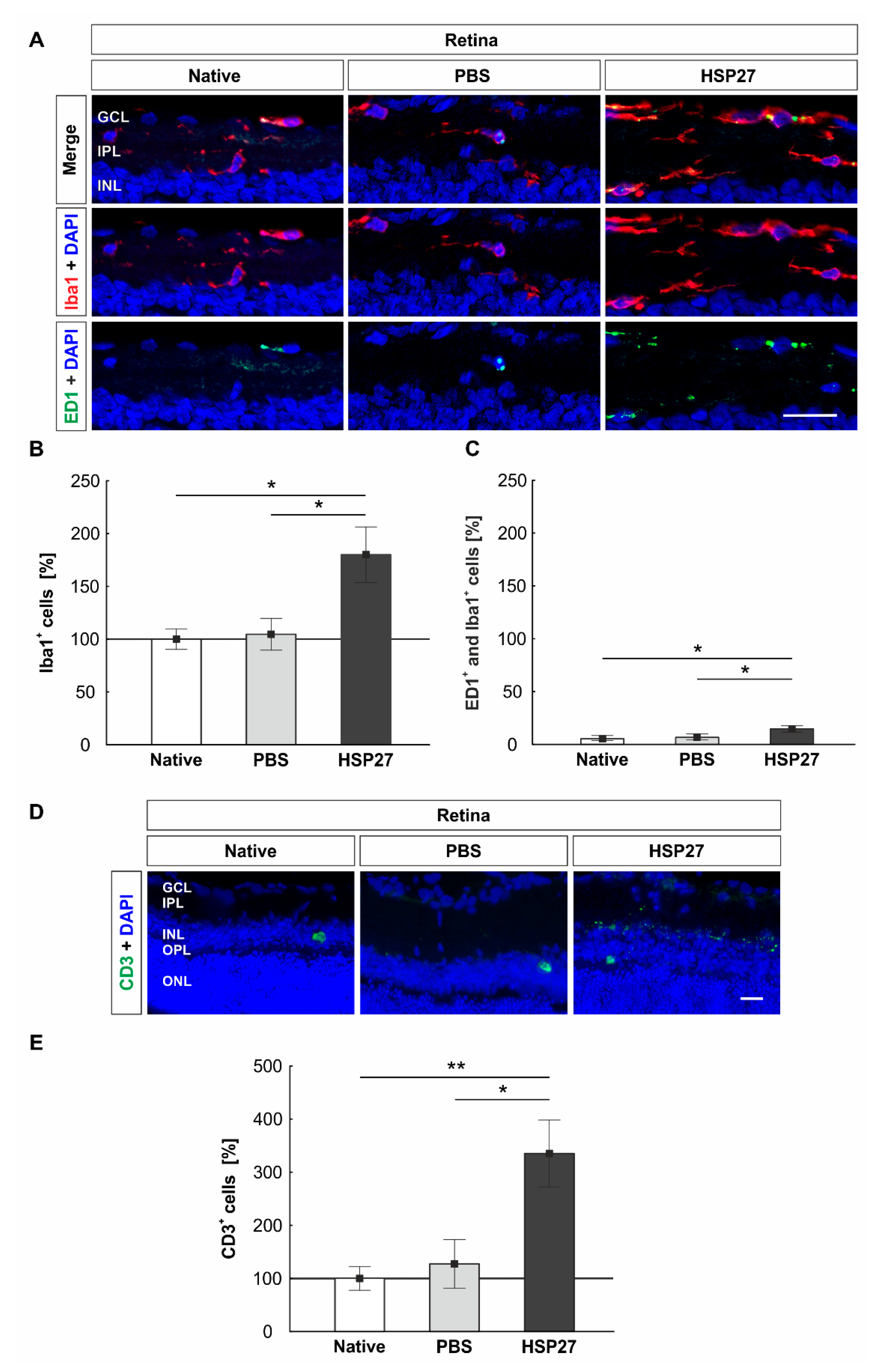
2.4. Increased Microglia/Macrophages and T-Cell Number in Retinae through HSP27
2.5. Unaltered HSP25 and NFκB Signals in Optic Nerves
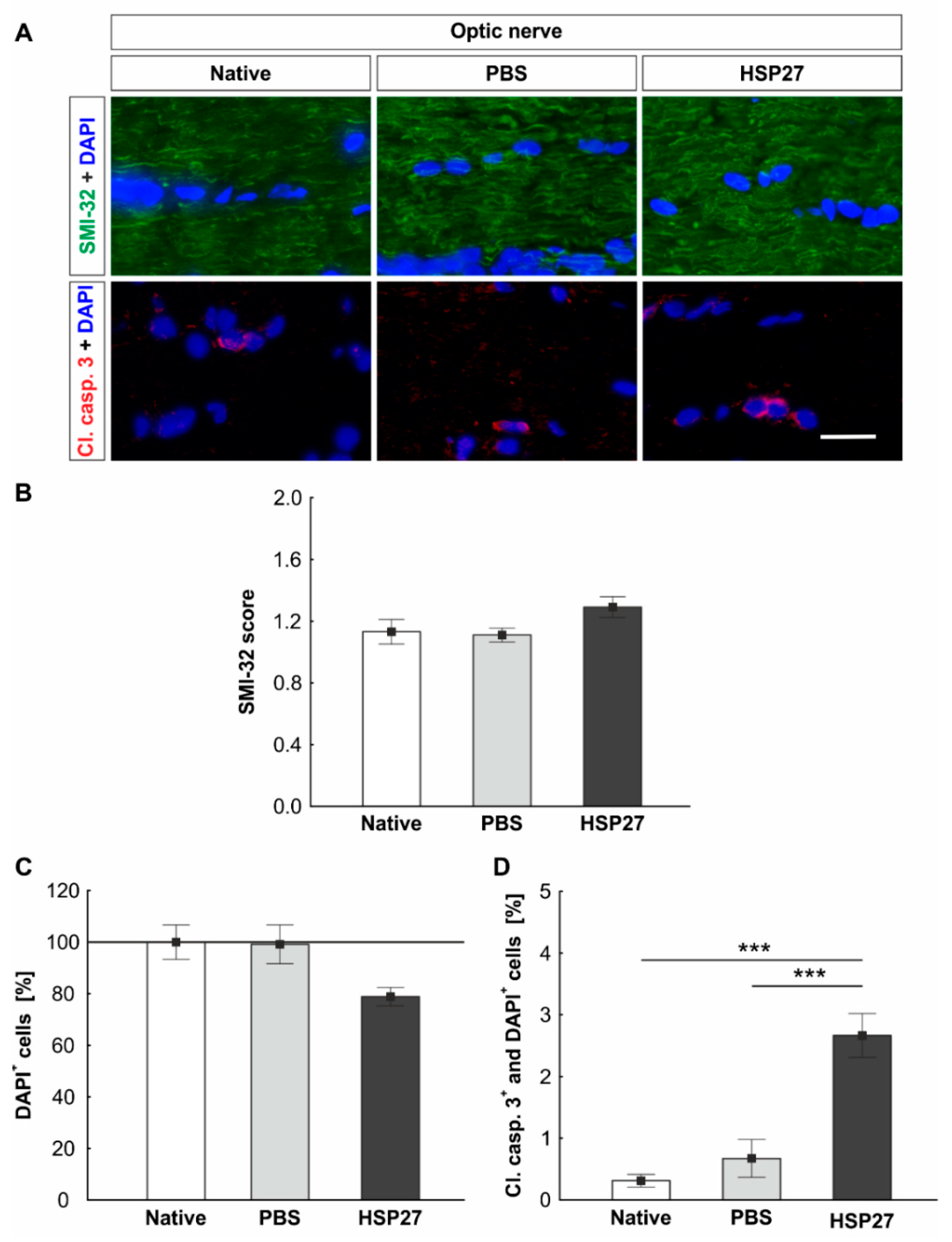
2.6. Increased Apoptosis Rate in the Optic Nerve
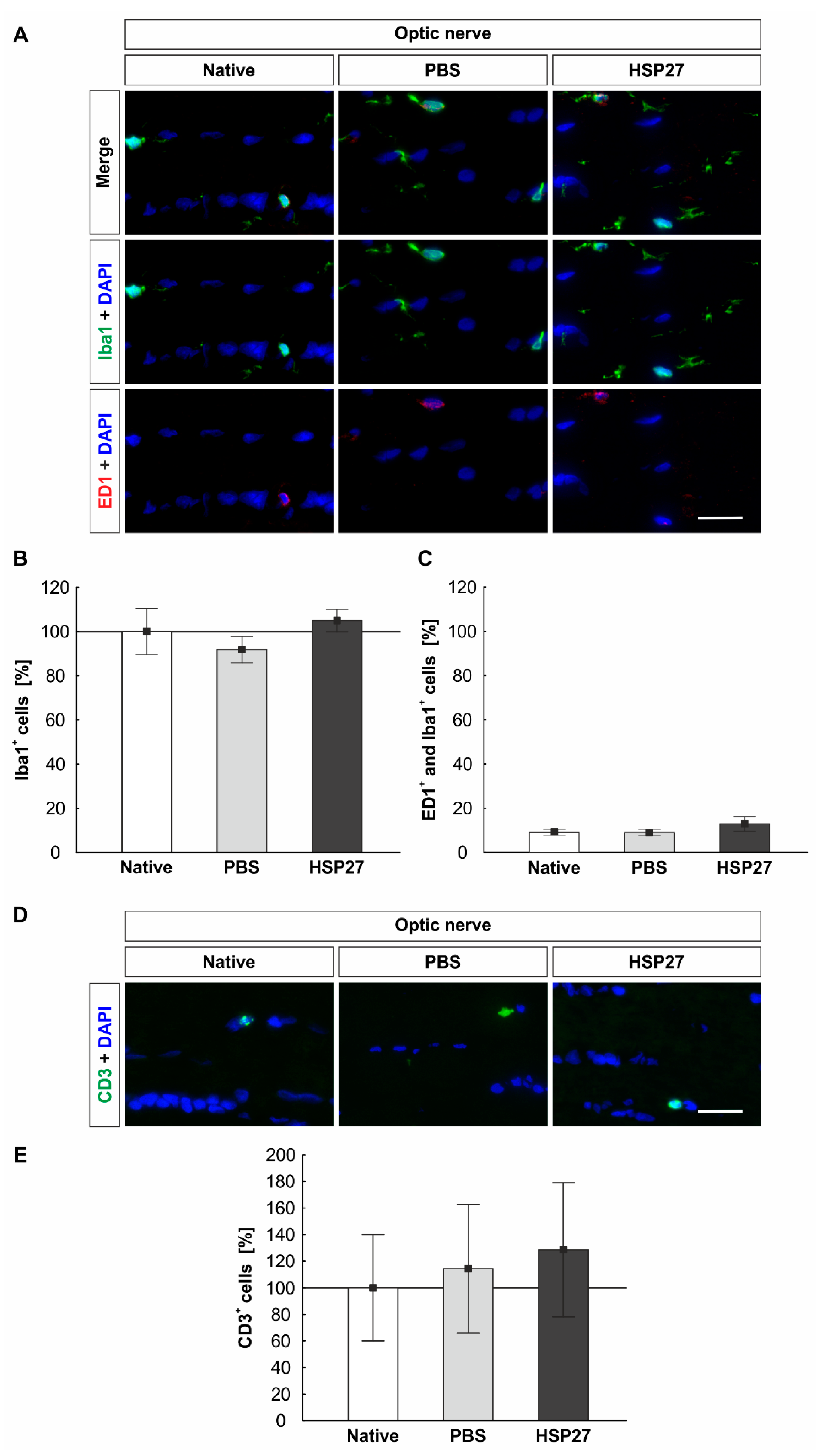
2.7. Constant Number of Microglia/Macrophages and T-Cells in the Optic Nerves
3. Discussion
3.1. Endogenous HSP as a Response to Stress
3.2. Activation of Caspase-Dependent Cell Death
3.3. NFκB Activation and Induction of an Immune Response
4. Materials and Methods
4.1. Animals
4.2. Intravitreal HSP27 Injection
4.3. Preparation of Retina and Optic Nerve
4.4. Specific Immunohistological Stainings of Retina and Optic Nerve
4.5. Quantitative Real-Time PCR Analysis of Retinal Tissue
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Brocchieri, L.; Karlin, S. Conservation among HSP60 sequences in relation to structure, function, and evolution. Protein Sci. 2000, 9, 476–486. [Google Scholar] [CrossRef] [Green Version]
- Kampinga, H.H.; Craig, E.A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 2010, 11, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Jakob, U.; Gaestel, M.; Engel, K.; Buchner, J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993, 268, 1517–1520. [Google Scholar] [CrossRef]
- Bruey, J.M.; Ducasse, C.; Bonniaud, P.; Ravagnan, L.; Susin, S.A.; Diaz-Latoud, C.; Gurbuxani, S.; Arrigo, A.P.; Kroemer, G.; Solary, E.; et al. Hsp27 negatively regulates cell death by interacting with cytochrome c. Nat. Cell. Biol. 2000, 2, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Havasi, A.; Li, Z.; Wang, Z.; Martin, J.L.; Botla, V.; Ruchalski, K.; Schwartz, J.H.; Borkan, S.C. Hsp27 inhibits Bax activation and apoptosis via a phosphatidylinositol 3-kinase-dependent mechanism. J. Biol. Chem. 2008, 283, 12305–12313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.; Cleveland, J.C.; Ao, L.; Li, J.; Zeng, Q.; Fullerton, D.A.; Meng, X. Human myocardium releases heat shock protein 27 (HSP27) after global ischemia: The proinflammatory effect of extracellular HSP27 through toll-like receptor (TLR)-2 and TLR4. J. Mol. Med. 2014, 20, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Salari, S.; Seibert, T.; Chen, Y.X.; Hu, T.; Shi, C.; Zhao, X.; Cuerrier, C.M.; Raizman, J.E.; O’Brien, E.R. Extracellular HSP27 acts as a signaling molecule to activate NF-kappaB in macrophages. Cell Stress Chaperones 2013, 18, 53–63. [Google Scholar] [CrossRef] [Green Version]
- EGS. European Glaucoma Society Terminology and Guidelines for Glaucoma, 4th Edition—Chapter 2: Classification and terminology Supported by the EGS Foundation: Part 1: Foreword; Introduction; Glossary; Chapter 2 Classification and Terminology. Br. J. Ophthalmol. 2017, 101, 73–127. [Google Scholar] [CrossRef] [Green Version]
- Tezel, G.; Hernandez, R.; Wax, M.B. Immunostaining of heat shock proteins in the retina and optic nerve head of normal and glaucomatous eyes. Arch. Ophthalmol. 2000, 118, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chidlow, G.; Wood, J.P.; Casson, R.J. Expression of inducible heat shock proteins Hsp27 and Hsp70 in the visual pathway of rats subjected to various models of retinal ganglion cell injury. PLoS ONE 2014, 9, e114838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalesnykas, G.; Niittykoski, M.; Rantala, J.; Miettinen, R.; Salminen, A.; Kaarniranta, K.; Uusitalo, H. The expression of heat shock protein 27 in retinal ganglion and glial cells in a rat glaucoma model. Neuroscience 2007, 150, 692–704. [Google Scholar] [CrossRef]
- Chen, H.; Cho, K.S.; Vu, T.H.K.; Shen, C.H.; Kaur, M.; Chen, G.; Mathew, R.; McHam, M.L.; Fazelat, A.; Lashkari, K.; et al. Commensal microflora-induced T cell responses mediate progressive neurodegeneration in glaucoma. Nat. Commun. 2018, 9, 3209. [Google Scholar] [CrossRef]
- Wax, M.B.; Tezel, G.; Yang, J.; Peng, G.; Patil, R.V.; Agarwal, N.; Sappington, R.M.; Calkins, D.J. Induced autoimmunity to heat shock proteins elicits glaucomatous loss of retinal ganglion cell neurons via activated T-cell-derived fas-ligand. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 12085–12096. [Google Scholar] [CrossRef] [Green Version]
- Joachim, S.C.; Grus, F.H.; Kraft, D.; White-Farrar, K.; Barnes, G.; Barbeck, M.; Ghanaati, S.; Cao, S.; Li, B.; Wax, M.B. Complex antibody profile changes in an experimental autoimmune glaucoma animal model. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4734–4742. [Google Scholar] [CrossRef]
- Grotegut, P.; Kuehn, S.; Dick, H.B.; Joachim, S.C. Destructive effect of intravitreal heat shock protein 27 application on retinal ganglion cells and neurofilament. Int. J. Mol. Sci. 2020, 21, 549. [Google Scholar] [CrossRef] [Green Version]
- Imai, Y.; Ibata, I.; Ito, D.; Ohsawa, K.; Kohsaka, S. A novel gene iba1 in the major histocompatibility complex class III region encoding an EF hand protein expressed in a monocytic lineage. Biochem. Biophys. Res. Commun. 1996, 224, 855–862. [Google Scholar] [CrossRef]
- McLean, P.J.; Kawamata, H.; Shariff, S.; Hewett, J.; Sharma, N.; Ueda, K.; Breakefield, X.O.; Hyman, B.T. TorsinA and heat shock proteins act as molecular chaperones: Suppression of alpha-synuclein aggregation. J. Neurochem. 2002, 83, 846–854. [Google Scholar] [CrossRef]
- Mehlen, P.; Coronas, V.; Ljubic-Thibal, V.; Ducasse, C.; Granger, L.; Jourdan, F.; Arrigo, A.P. Small stress protein Hsp27 accumulation during dopamine-mediated differentiation of rat olfactory neurons counteracts apoptosis. Cell Death Differ. 1999, 6, 227–233. [Google Scholar] [CrossRef]
- Huot, J.; Houle, F.; Spitz, D.R.; Landry, J. HSP27 phosphorylation-mediated resistance against actin fragmentation and cell death induced by oxidative stress. Cancer Res. 1996, 56, 273–279. [Google Scholar] [PubMed]
- Krueger-Naug, A.M.; Hopkins, D.A.; Armstrong, J.N.; Plumier, J.C.; Currie, R.W. Hyperthermic induction of the 27-kDa heat shock protein (Hsp27) in neuroglia and neurons of the rat central nervous system. J. Comp. Neurol. 2000, 428, 495–510. [Google Scholar] [CrossRef]
- Matozaki, M.; Saito, Y.; Yasutake, R.; Munira, S.; Kaibori, Y.; Yukawa, A.; Tada, M.; Nakayama, Y. Involvement of Stat3 phosphorylation in mild heat shock-induced thermotolerance. Exp. Cell Res. 2019, 377, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Fischer, D.; Andreadaki, A.; Bartelt-Kirbach, B.; Golenhofen, N. Induction and phosphorylation of the small heat shock proteins HspB1/Hsp25 and HspB5/alphaB-crystallin in the rat retina upon optic nerve injury. Cell Stress Chaperones 2016, 21, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Fileta, J.B.; Filippopoulos, T.; Ray, A.; Dobberfuhl, A.; Grosskreutz, C.L. Hsp27 phosphorylation in experimental glaucoma. Invest. Ophthalmol. Vis. Sci. 2007, 48, 4129–4135. [Google Scholar] [CrossRef]
- Kuehn, S.; Meissner, W.; Grotegut, P.; Theiss, C.; Dick, H.B.; Joachim, S.C. Intravitreal S100B Injection Leads to Progressive Glaucoma Like Damage in Retina and Optic Nerve. Front. Cell. Neurosci. 2018, 12, 312. [Google Scholar] [CrossRef] [Green Version]
- Ueki, Y.; Reh, T.A. Activation of BMP-Smad1/5/8 signaling promotes survival of retinal ganglion cells after damage in vivo. PLoS ONE 2012, 7, e38690. [Google Scholar] [CrossRef]
- Gao, L.; Zheng, Q.J.; Ai, L.Q.; Chen, K.J.; Zhou, Y.G.; Ye, J.; Liu, W. Exploration of the glutamate-mediated retinal excitotoxic damage: A rat model of retinal neurodegeneration. Int. J. Ophthalmol. 2018, 11, 1746–1754. [Google Scholar]
- Lam, T.T.; Abler, A.S.; Kwong, J.M.; Tso, M.O. N-methyl-D-aspartate (NMDA)--induced apoptosis in rat retina. Invest. Ophthalmol. Vis. Sci. 1999, 40, 2391–2397. [Google Scholar]
- Batulan, Z.; Pulakazhi Venu, V.K.; Li, Y.; Koumbadinga, G.; Alvarez-Olmedo, D.G.; Shi, C.; O’Brien, E.R. Extracellular Release and Signaling by Heat Shock Protein 27: Role in Modifying Vascular Inflammation. Front. Immunol. 2016, 7, 285. [Google Scholar] [CrossRef] [Green Version]
- Thuringer, D.; Jego, G.; Wettstein, G.; Terrier, O.; Cronier, L.; Yousfi, N.; Hebrard, S.; Bouchot, A.; Hazoume, A.; Joly, A.L.; et al. Extracellular HSP27 mediates angiogenesis through Toll-like receptor 3. FASEB J 2013, 27, 4169–4183. [Google Scholar] [CrossRef]
- Aliprantis, A.O.; Yang, R.B.; Weiss, D.S.; Godowski, P.; Zychlinsky, A. The apoptotic signaling pathway activated by Toll-like receptor-2. Embo J. 2000, 19, 3325–3336. [Google Scholar] [CrossRef] [Green Version]
- Han, K.J.; Su, X.; Xu, L.G.; Bin, L.H.; Zhang, J.; Shu, H.B. Mechanisms of the TRIF-induced interferon-stimulated response element and NF-kappaB activation and apoptosis pathways. J. Biol. Chem. 2004, 279, 15652–15661. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Budihardjo, I.; Zou, H.; Slaughter, C.; Wang, X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 1998, 94, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Fileta, J.B.; Dobberfuhl, A.; Filippopolous, T.; Guo, Y.; Kwon, G.; Grosskreutz, C.L. Calcineurin cleavage is triggered by elevated intraocular pressure, and calcineurin inhibition blocks retinal ganglion cell death in experimental glaucoma. Proc. Natl. Acad. Sci. USA 2005, 102, 12242–12247. [Google Scholar] [CrossRef] [Green Version]
- Solt, L.A.; May, M.J. The IkappaB kinase complex: Master regulator of NF-kappaB signaling. Immunol. Res. 2008, 42, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [Green Version]
- Bosco, A.; Steele, M.R.; Vetter, M.L. Early microglia activation in a mouse model of chronic glaucoma. J. Comp. Neurol. 2011, 519, 599–620. [Google Scholar] [CrossRef] [Green Version]
- Ebneter, A.; Casson, R.J.; Wood, J.P.; Chidlow, G. Microglial activation in the visual pathway in experimental glaucoma: Spatiotemporal characterization and correlation with axonal injury. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6448–6460. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Lorenz, K.; Beck, S.; Keilani, M.M.; Wasielica-Poslednik, J.; Pfeiffer, N.; Grus, F.H. Course of serum autoantibodies in patients after acute angle-closure glaucoma attack. Clin. Exp. Ophthalmol. 2017, 45, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G.; Seigel, G.M.; Wax, M.B. Autoantibodies to small heat shock proteins in glaucoma. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2277–2287. [Google Scholar]
- Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Grus, F.H. Antibodies to alpha B-crystallin, vimentin, and heat shock protein 70 in aqueous humor of patients with normal tension glaucoma and IgG antibody patterns against retinal antigen in aqueous humor. Curr. Eye Res. 2007, 32, 501–509. [Google Scholar] [CrossRef]
- Noristani, R.; Kuehn, S.; Stute, G.; Reinehr, S.; Stellbogen, M.; Dick, H.B.; Joachim, S.C. Retinal and Optic Nerve Damage is Associated with Early Glial Responses in an Experimental Autoimmune Glaucoma Model. J. Mol. Neurosci. Mn 2016, 58, 470–482. [Google Scholar] [CrossRef]
- Reinehr, S.; Reinhard, J.; Gandej, M.; Kuehn, S.; Noristani, R.; Faissner, A.; Dick, H.B.; Joachim, S.C. Simultaneous Complement Response via Lectin Pathway in Retina and Optic Nerve in an Experimental Autoimmune Glaucoma Model. Front. Cell. Neurosci. 2016, 10, 140. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Secondary Antibodies | ||||
|---|---|---|---|---|---|
| Name and host | Company | Dilution | Name | Company | Dilution |
| Retinal Cross-sections | |||||
| Bax (rabbit) | Abcam | 1:100 | Donkey anti-rabbit Alexa Fluor 555 | Abcam | 1:400 |
| CD3-FITC (directly labeled antibody, mouse) | BD Biosciences | 1:1000 | − | − | − |
| ED1 (mouse) | Millipore | 1:250 | Goat anti-mouse Alexa Fluor 488 | Invitrogen | 1:400 |
| GFAP (chicken) | Millipore | 1:400 | Donkey anti-chicken Cy3 | Millipore | 1:500 |
| HSP25 (rabbit) | Enzo Life Sciences | 1:100 | Donkey anti-rabbit Alexa Fluor 488 | Jackson ImmunoResearch | 1:500 |
| Iba1 (rabbit) | Wako | 1:500 | Goat anti-rabbit Cy3 | Linaris | 1:300 |
| NeuN (chicken) | Millipore | 1:500 | Donkey anti-chicken Cy3 | Millipore | 1:500 |
| NFκB (rabbit) | Santa Cruz | 1:500 | Goat anti-rabbit Alexa Fluor 488 | Invitrogen | 1:500 |
| RBPMS (guinea pig) | Millipore | 1:250 | Donkey anti-guinea pig Alexa 488 | Jackson ImmunoResearch | 1:400 |
| Longitudinal Optic NerveSections | |||||
| CD3-FITC (directly labeled antibody, mouse) | BD Biosciences | 1:100 | − | − | − |
| Cl. caspase3 (rabbit) | Sigma-Aldrich | 1:300 | Donkey anti-rabbit Alexa Fluor 555 | Invitrogen | 1:400 |
| ED1 (mouse) | Millipore | 1:200 | Goat anti-mouse Alexa Fluor 555 | Invitrogen | 1:500 |
| HSP25 (rabbit) | Enzo Life Sciences | 1:100 | Donkey anti-rabbit Alexa Fluor 488 | Jackson ImmunoResearch | 1:500 |
| Iba1 (rabbit) | Wako | 1:400 | Goat anti-rabbit Alexa Fluor 488 | Invitrogen | 1:500 |
| SMI-32 (mouse) | Biolegend | 1:2000 | Goat anti-mouse Alexa Fluor 488 | Invitrogen | 1:400 |
| Gene | Primer Sequence | GenBank Accession Number |
|---|---|---|
| Actb-F | cccgcgagtacaaccttct | NM_031144.3 |
| Actb-R | cgtcatccatggcgaact | |
| Akt1-F | gacgtagccattgtgaaggag | NM_033230.2 |
| Akt1-R | ccatcattcttgaggaggaagt | |
| Casp3-F | ccgacttcctgtatgcttactcta | NM_012922.2 |
| Casp3-R | catgacccgtcccttgaa | |
| Casp8-F | agagcctgagggaaagatgtc | NM_022277.1 |
| Casp8-R | tcacatcatagttcacgccagt | |
| Casp9-F | cgtggtggtcatcctctctc | NM_031632.1 |
| Casp9-R | gagcatccatctgtgccata | |
| Hspb1-F | gaggagctcacagttaagaccaa | NM_031970.4 |
| Hspb1-R | ttcatcctgcctttcttcgt | |
| Hspb70-F | catatccaatatctttgaggtgga | XM_017601842.1 |
| Hspb70-R | tggggaagacttcacagtca | |
| Hsp90aa1-F | gggagctcatttccaactcc | NM_175761.2 |
| Hsp90aa1-R | gggttcggtcttgcttgtt | |
| Mapk14-F | gaacttcgcaaatgtatttattggt | NM_031020.2 |
| Mapk14-R | cgagtccaaaaccagcatc | |
| Nfκb-F | ctggcagctcttctcaaagc | NM_001276711.1 |
| Nfκb-R | caggtcatagagaggctcaa | |
| Pou4f1-F | ctggccaacctcaagatcc | XM_008770931 |
| Pou4f1-R | cgtgagcgactcgaacct | |
| Ppid-F | tgctggaccaaacacaaatg | M19553.1 |
| Ppid-R | cttcccaaagaccacatgct | |
| Prkd1-F | tgctccttcaggactcctct | NM_001276715.1 |
| Prkd1-R | gaagccacattcagggaact |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grotegut, P.; Hoerdemann, P.J.; Reinehr, S.; Gupta, N.; Dick, H.B.; Joachim, S.C. Heat Shock Protein 27 Injection Leads to Caspase Activation in the Visual Pathway and Retinal T-Cell Response. Int. J. Mol. Sci. 2021, 22, 513. https://doi.org/10.3390/ijms22020513
Grotegut P, Hoerdemann PJ, Reinehr S, Gupta N, Dick HB, Joachim SC. Heat Shock Protein 27 Injection Leads to Caspase Activation in the Visual Pathway and Retinal T-Cell Response. International Journal of Molecular Sciences. 2021; 22(2):513. https://doi.org/10.3390/ijms22020513
Chicago/Turabian StyleGrotegut, Pia, Philipp Johannes Hoerdemann, Sabrina Reinehr, Nupur Gupta, H. Burkhard Dick, and Stephanie C. Joachim. 2021. "Heat Shock Protein 27 Injection Leads to Caspase Activation in the Visual Pathway and Retinal T-Cell Response" International Journal of Molecular Sciences 22, no. 2: 513. https://doi.org/10.3390/ijms22020513