
Transcriptomic Signatures and Upstream Regulation in Human Skeletal Muscle Adapted to Disuse and Aerobic Exercise


Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEO ID | Model and Reference | Duration (d or wk)/Recovery Time (h) | N of Subjects (Males/Females) | Mean Age (Range) | Mean BMI | Muscle | Method | Individual Study: N of UP/DOWN Genes | Meta-Analysis: N of UP/DOWN Genes |
|---|---|---|---|---|---|---|---|---|---|
| GSE113165 | Bed rest [11] | 5 d | 28 (13/15) | 53 (30–76) | 23.9 | VL | RNA seq | 339/696 | 76/205 |
| GSE126865 GSE130722 | Bed rest [12] | 10 d | 3 (3/0) 5 (5/0) | 70 (64–76) 66 (61–73) | 29.0 27.6 | VL | RNA seq | 101/310 | |
| GSE24215 | Bed rest [13] | 10 d | 10 (10/0) | 25 (24–26) | 24.0 | VL | DNA array | 47/184 | |
| GSE14901 | A leg immobilization [14] | 14 d | 24 (12/12) | 21 (19–23) | 23.6 | VL | DNA array | 479/571 | |
| GSE104999 | Bed rest [15] | 21 d | 12 (12/0) | 27 (21–33) | 24.0 | VL | DNA array | 59/170 | |
| GSE33886 | A leg immobilization [16] | 21 d | 6 (-) | 21 (21–21) | 22.0 | VL | DNA array | 0/0 | - |
| GSE147494 | Cycling training [17] | 5 wk | 40 (40/0) | 22 (20–24) | 22.4 | VL | DNA array | 134/29 | 374/119 |
| GSE35661 | Cycling training [18] | 6 wk | 24 (24/0) | 23 (22–24) | 23.0 | VL | DNA array | 563/142 | |
| GSE27536 | Cycling training [19] | 8 wk | 12 (10/2) | 65 (-) | - | VL | DNA array | 1212/386 | |
| GSE120862 | Cycling training [20] | 8 wk 1 leg 8 wk 1 leg | 7 (7/0) | 21 (21–24) | 23.0 | VL | RNA seq | 2145/1508 2127/1119 | |
| GSE1786 | Cycling training [21] | 12 wk | 6 (6/0) | 68 (65–71) | 27.0 | VL | DNA array | 194/88 | |
| GSE60591 GSE60833 GSE60833 | Knee extension training [22] | 12 wk 1 leg 12 wk 1 leg 12 wk 1 leg | 22 (11/11) 11 (6/5) 10 (5/5) | 27 (26–28) 28 (25–32) | 24.0 25.9 | VL | RNA seq | 931/322 111/155 58/97 | |
| GSE117070 | Cycling training [23] | 20 wk | 41 (-) | - | - | VL | DNA array | 30/61 | |
| GSE111551 | Running training | 18 wk | 13 (13/0) | 27 (22–30) | - | VL | DNA array | 0/0 | - |
| GSE41769 GSE71972 GSE59363 | Acute knee extension [24] Acute knee extension [25] Acute cycling [26] | 0 h 0 h 0 h | 9 (9/0) 8 (-) 7 (7/0) | 52 (47–57) 23 (19–27) 56 (54–58) | 26.0 23.7 27.4 | VL VL VL | DNA array RNA seq DNA array | 218/34 399/703 | 140/81 |
| 0/0 | - | ||||||||
| GSE43219 GSE33603 GSE107934 | Acute cycling [27] Acute cycling [28] Acute cycling [29] | 0.5 h 0.5 h 1 h | 14 (9/5) 11 (11/0) 6 (6/0) | 33 (23–48) 22 (21–23) 27 (24–30) | 26.5 24.5 24.7 | VL VL VL | DNA array DNA array RNA seq | 456/291 378/134 47/0 | 272/50 |
| GSE120862 | Acute knee extension [20] | 1 h | 7 (7/0) | 21 (21–24) | 23.0 | VL | RNA seq | 802/534 | |
| GSE164081 | Acute cycling | 1 h | 10 (10/0) | 32 (30–36) | 23.1 | VL | CAGE | 541/166 | |
| GSE27285 | Acute cycling [30] | 3 h | 8 (8/0) | 33 (27–39) | 24.0 | VL | DNA array | 462/320 | 374/80 |
| GSE33603 | Acute cycling [28] | 3 h | 11 (11/0) | 22 (21–23) | 24.5 | VL | DNA array | 932/592 | |
| GSE71972 | Acute knee extension [25] | 3 h | 8 (-) | 23 (19–27) | 23.7 | VL | RNA seq | 1411/635 | |
| GSE164081 | Acute cycling | 3 h | 10 (10/0) | 32 (30–36) | 23.1 | VL | CAGE | 769/325 | |
| GSE43856 | Acute cycling and running [31] | 3 h | 8 (8/0) | 25 (21–29) | 23.1 | VL | DNA array | 0/0 | - |
| GSE59363 | Acute cycling [26] | 3 h | 7 (7/0) | 56 (54–58) | 27.4 | VL | DNA array | 0/0 | - |
| GSE107934 | Acute cycling [29] | 4 h | 6 (6/0) | 27 (24–30) | 24.7 | VL | RNA seq | 193/20 | 153/74 |
| GSE86931 | Acute cycling [32] | 4 h | 2 (2/0) | 24 (22–25) | 22.0 | VL | RNA seq | 101/45 | |
| GSE87748 | Acute cycling [33] | 4 h | 10 (10/0) | 23 (21–24) | 23.1 | VL | RNA seq | 183/173 | |
| GSE120862 | Acute knee extension [20] | 4 h | 7 (7/0) | 21 (21–24) | 23.0 | VL | RNA seq | 1295/909 |
2. Results
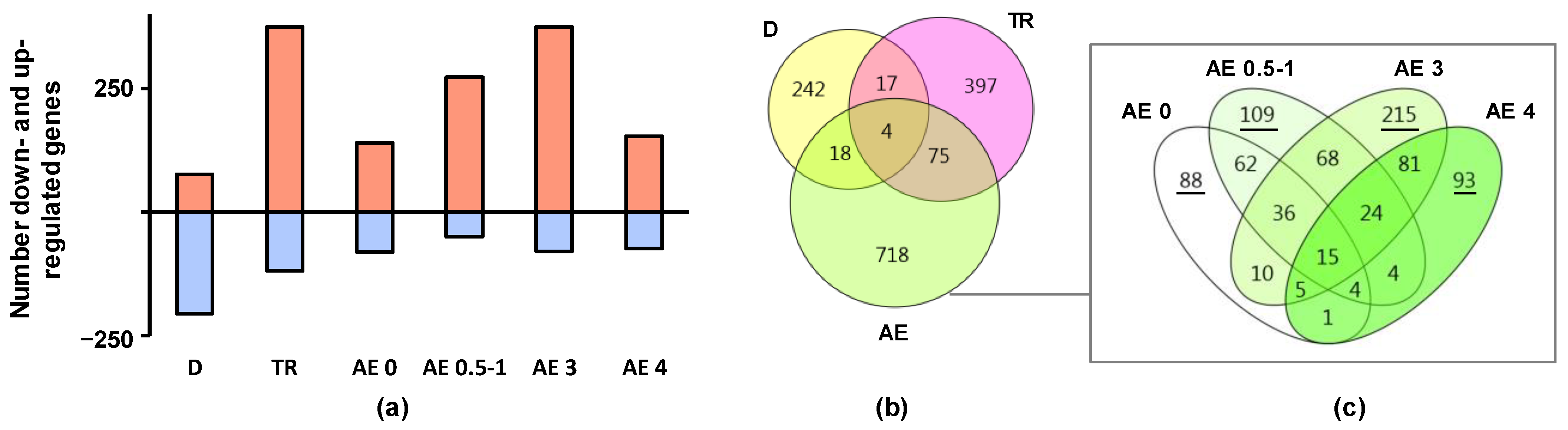
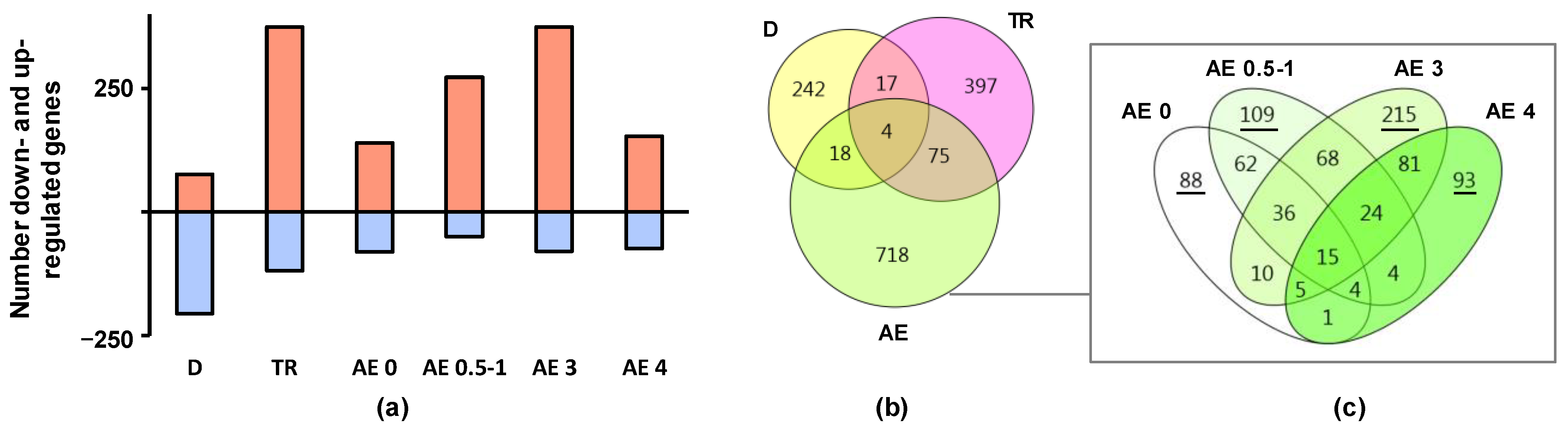
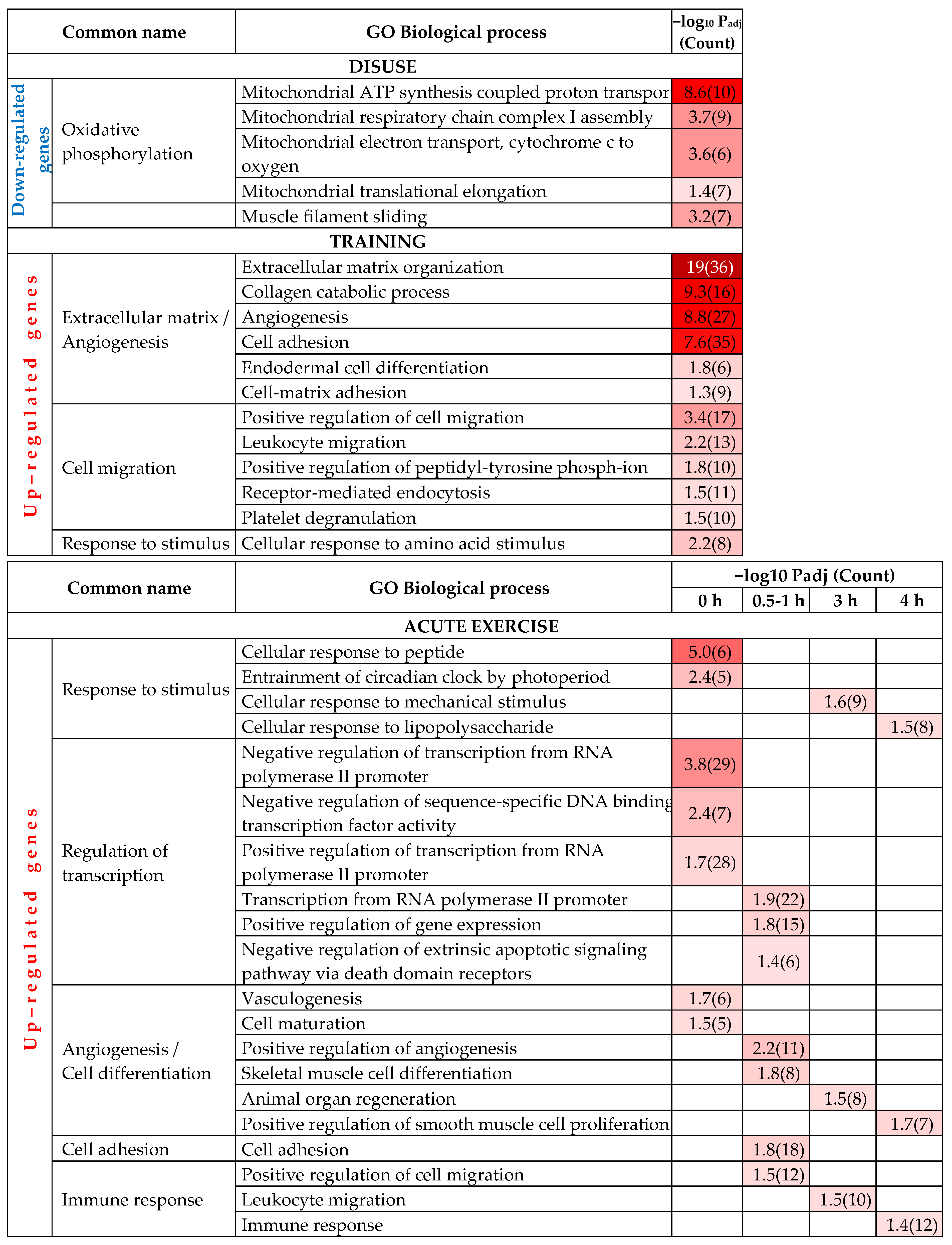
2.1. Regulation of Biological Processes
2.2. Expression of Genes Encoding Specific Sets of Proteins
2.3. Transcription Factors Induced by Disuse and Exercise
3. Discussion
3.1. The Role of Transcription in Phenotypic Changes Induced by Disuse and Training
3.2. Transcription Factors Induced by Disuse and Exercise
3.3. Limitations and Further Directions
3.4. Conclusions
4. Materials and Methods
4.1. Search Strategy and Eligibility Criteria
4.2. Data Synthesis and Analysis
4.2.1. DNA Microarray Data Processing
4.2.2. RNA Sequencing Data Processing
4.2.3. Differentially Expressed Genes Related with a Specific Intervention (Meta-Analysis)
4.2.4. Bioinformatics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Demontis, F.; Piccirillo, R.; Goldberg, A.L.; Perrimon, N. The influence of skeletal muscle on systemic aging and lifespan. Aging Cell 2013, 12, 943–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervenka, I.; Agudelo, L.Z.; Ruas, J.L. Kynurenines: Tryptophan’s metabolites in exercise, inflammation, and mental health. Science 2017, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.; Nathan, J.A.; Goldberg, A.L. Muscle wasting in disease: Molecular mechanisms and promising therapies. Nat. Rev. Drug Discov. 2015, 14, 58–74. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Saltin, B. Exercise as medicine—Evidence for prescribing exercise as therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. 3), 1–72. [Google Scholar] [CrossRef] [Green Version]
- Malatras, A.; Duguez, S.; Duddy, W. Muscle Gene Sets: A versatile methodological aid to functional genomics in the neuromuscular field. Skelet. Muscle 2019, 9, 10. [Google Scholar] [CrossRef]
- Soul, J.; Hardingham, T.E.; Boot-Handford, R.P.; Schwartz, J.M. SkeletalVis: An exploration and meta-analysis data portal of cross-species skeletal transcriptomics data. Bioinformatics 2019, 35, 2283–2290. [Google Scholar] [CrossRef] [Green Version]
- Reibe, S.; Hjorth, M.; Febbraio, M.A.; Whitham, M. GeneXX: An online tool for the exploration of transcript changes in skeletal muscle associated with exercise. Physiol. Genom. 2018, 50, 376–384. [Google Scholar] [CrossRef]
- Pillon, N.J.; Gabriel, B.M.; Dollet, L.; Smith, J.A.B.; Sardon, P.L.; Botella, J.; Bishop, D.J.; Krook, A.; Zierath, J.R. Transcriptomic profiling of skeletal muscle adaptations to exercise and inactivity. Nat. Commun. 2020, 11, 470. [Google Scholar] [CrossRef] [Green Version]
- Kolde, R.; Laur, S.; Adler, P.; Vilo, J. Robust rank aggregation for gene list integration and meta-analysis. Bioinformatics 2012, 28, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Mahmassani, Z.S.; Reidy, P.T.; McKenzie, A.I.; Stubben, C.; Howard, M.T.; Drummond, M.J. Age-dependent skeletal muscle transcriptome response to bed rest-induced atrophy. J. Appl. Physiol. 2019, 126, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Trevino, M.B.; Zhang, X.; Standley, R.A.; Wang, M.; Han, X.; Reis, F.C.G.; Periasamy, M.; Yu, G.; Kelly, D.P.; Goodpaster, B.H.; et al. Loss of mitochondrial energetics is associated with poor recovery of muscle function but not mass following disuse atrophy. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E899–E910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibegovic, A.C.; Sonne, M.P.; Hojbjerre, L.; Bork-Jensen, J.; Jacobsen, S.; Nilsson, E.; Faerch, K.; Hiscock, N.; Mortensen, B.; Friedrichsen, M.; et al. Insulin resistance induced by physical inactivity is associated with multiple transcriptional changes in skeletal muscle in young men. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E752–E763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abadi, A.; Glover, E.I.; Isfort, R.J.; Raha, S.; Safdar, A.; Yasuda, N.; Kaczor, J.J.; Melov, S.; Hubbard, A.; Qu, X.; et al. Limb immobilization induces a coordinate down-regulation of mitochondrial and other metabolic pathways in men and women. PLoS ONE 2009, 4, e6518. [Google Scholar] [CrossRef] [Green Version]
- Rullman, E.; Fernandez-Gonzalo, R.; Mekjavic, I.B.; Gustafsson, T.; Eiken, O. MEF2 as upstream regulator of the transcriptome signature in human skeletal muscle during unloading. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R799–R809. [Google Scholar] [CrossRef] [Green Version]
- Lammers, G.; Poelkens, F.; van Duijnhoven, N.T.; Pardoel, E.M.; Hoenderop, J.G.; Thijssen, D.H.; Hopman, M.T. Expression of genes involved in fatty acid transport and insulin signaling is altered by physical inactivity and exercise training in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E1245–E1251. [Google Scholar] [CrossRef] [Green Version]
- Knuiman, P.; Hangelbroek, R.; Boekschoten, M.; Hopman, M.; Mensink, M. Impact of protein supplementation during endurance training on changes in skeletal muscle transcriptome. BMC Genom. 2020, 21, 397. [Google Scholar] [CrossRef]
- Keller, P.; Vollaard, N.B.; Gustafsson, T.; Gallagher, I.J.; Sundberg, C.J.; Rankinen, T.; Britton, S.L.; Bouchard, C.; Koch, L.G.; Timmons, J.A. A transcriptional map of the impact of endurance exercise training on skeletal muscle phenotype. J. Appl. Physiol. 2011, 110, 46–59. [Google Scholar] [CrossRef] [Green Version]
- Turan, N.; Kalko, S.; Stincone, A.; Clarke, K.; Sabah, A.; Howlett, K.; Curnow, S.J.; Rodriguez, D.A.; Cascante, M.; O’Neill, L.; et al. A systems biology approach identifies molecular networks defining skeletal muscle abnormalities in chronic obstructive pulmonary disease. PLoS Comput. Biol. 2011, 7, e1002129. [Google Scholar] [CrossRef] [Green Version]
- Popov, D.V.; Makhnovskii, P.A.; Shagimardanova, E.I.; Gazizova, G.R.; Lysenko, E.A.; Gusev, O.A.; Vinogradova, O.L. Contractile activity-specific transcriptome response to acute endurance exercise and training in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2019, 316, e605–e614. [Google Scholar] [CrossRef]
- Radom-Aizik, S.; Hayek, S.; Shahar, I.; Rechavi, G.; Kaminski, N.; Ben-Dov, I. Effects of aerobic training on gene expression in skeletal muscle of elderly men. Med. Sci. Sports Exerc. 2005, 37, 1680–1696. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, M.E.; Huss, M.; Solnestam, B.W.; Kjellqvist, S.; Lundeberg, J.; Sundberg, C.J. The human skeletal muscle transcriptome: Sex differences, alternative splicing, and tissue homogeneity assessed with RNA sequencing. FASEB J. 2014, 28, 4571–4581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.; Ricciardi, S.; Pearson, T.; Bharudin, I.; Davidsen, P.K.; Bonomo, M.; Brina, D.; Scagliola, A.; Simpson, D.M.; Beynon, R.J.; et al. The role of Eif6 in skeletal muscle homeostasis revealed by endurance training co-expression networks. Cell Rep. 2017, 21, 1507–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catoire, M.; Mensink, M.; Boekschoten, M.V.; Hangelbroek, R.; Muller, M.; Schrauwen, P.; Kersten, S. Pronounced effects of acute endurance exercise on gene expression in resting and exercising human skeletal muscle. PLoS ONE 2012, 7, e51066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, S.A.; Hocker, A.D.; Mangum, J.E.; Luttrell, M.J.; Turnbull, D.W.; Struck, A.J.; Ely, M.R.; Sieck, D.C.; Dreyer, H.C.; Halliwill, J.R. Evidence of a broad histamine footprint on the human exercise transcriptome. J. Physiol. 2016, 594, 5009–5023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, J.S.; Zhao, X.; Irmler, M.; Liu, X.; Hoene, M.; Scheler, M.; Li, Y.; Beckers, J.; de Angelis, M.H.; Haring, H.U.; et al. Type 2 diabetes alters metabolic and transcriptional signatures of glucose and amino acid metabolism during exercise and recovery. Diabetologia 2015, 58, 1845–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, C.S.; Mielke, C.; Cordova, J.M.; Langlais, P.R.; Bowen, B.; Miranda, D.; Coletta, D.K.; Mandarino, L.J. Gene and MicroRNA Expression Responses to Exercise; Relationship with Insulin Sensitivity. PLoS ONE 2015, 10, e0127089. [Google Scholar] [CrossRef]
- Crane, J.D.; Ogborn, D.I.; Cupido, C.; Melov, S.; Hubbard, A.; Bourgeois, J.M.; Tarnopolsky, M.A. Massage therapy attenuates inflammatory signaling after exercise-induced muscle damage. Sci. Transl. Med. 2012, 4, 119ra13. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, J.M.; D’Lugos, A.C.; Naymik, M.A.; Siniard, A.L.; Wolfe, A.J.; Curtis, D.R.; Huentelman, M.J.; Carroll, C.C. Transcriptome response of human skeletal muscle to divergent exercise stimuli. J. Appl. Physiol. 2018, 124, 1529–1540. [Google Scholar] [CrossRef]
- Rowlands, D.S.; Thomson, J.S.; Timmons, B.W.; Raymond, F.; Fuerholz, A.; Mansourian, R.; Zwahlen, M.C.; Metairon, S.; Glover, E.; Stellingwerff, T.; et al. Transcriptome and translational signaling following endurance exercise in trained skeletal muscle: Impact of dietary protein. Physiol. Genom. 2011, 43, 1004–1020. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, O.; Sabapathy, S.; Ashton, K.J.; Desbrow, B.; Peake, J.M.; Lazarus, R.; Wessner, B.; Cameron-Smith, D.; Wagner, K.H.; Haseler, L.J.; et al. Time course-dependent changes in the transcriptome of human skeletal muscle during recovery from endurance exercise: From inflammation to adaptive remodeling. J. Appl. Physiol. 2014, 116, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Popov, D.V.; Makhnovskii, P.A.; Kurochkina, N.S.; Lysenko, E.A.; Vepkhvadze, T.F.; Vinogradova, O.L. Intensity-dependent gene expression after aerobic exercise in endurance-trained skeletal muscle. Biol. Sport 2018, 35, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Pattamaprapanont, P.; Garde, C.; Fabre, O.; Barres, R. Muscle Contraction Induces Acute Hydroxymethylation of the Exercise-Responsive Gene Nr4a3. Front. Endocrinol. 2016, 7, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.; Ross, R.S. A mini review: Proteomics approaches to understand disused vs. exercised human skeletal muscle. Physiol. Genom. 2018, 50, 746–757. [Google Scholar] [CrossRef]
- Makhnovskii, P.A.; Zgoda, V.G.; Bokov, R.O.; Shagimardanova, E.I.; Gazizova, G.R.; Gusev, O.A.; Lysenko, E.A.; Kolpakov, F.A.; Vinogradova, O.L.; Popov, D.V. Regulation of proteins in human skeletal muscle: The role of transcription. Sci.Rep. 2020, 10, 3514. [Google Scholar] [CrossRef] [Green Version]
- Schild, M.; Ruhs, A.; Beiter, T.; Zugel, M.; Hudemann, J.; Reimer, A.; Krumholz-Wagner, I.; Wagner, C.; Keller, J.; Eder, K.; et al. Basal and exercise induced label-free quantitative protein profiling of m. vastus lateralis in trained and untrained individuals. J. Proteom. 2015, 122, 119–132. [Google Scholar] [CrossRef]
- Bamman, M.M.; Clarke, M.S.; Feeback, D.L.; Talmadge, R.J.; Stevens, B.R.; Lieberman, S.A.; Greenisen, M.C. Impact of resistance exercise during bed rest on skeletal muscle sarcopenia and myosin isoform distribution. J. Appl. Physiol. 1998, 84, 157–163. [Google Scholar] [CrossRef]
- Hortobagyi, T.; Dempsey, L.; Fraser, D.; Zheng, D.; Hamilton, G.; Lambert, J.; Dohm, L. Changes in muscle strength, muscle fibre size and myofibrillar gene expression after immobilization and retraining in humans. J. Physiol. 2000, 524 Pt 1, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Hvid, L.; Aagaard, P.; Justesen, L.; Bayer, M.L.; Andersen, J.L.; Ortenblad, N.; Kjaer, M.; Suetta, C. Effects of aging on muscle mechanical function and muscle fiber morphology during short-term immobilization and subsequent retraining. J. Appl. Physiol. 2010, 109, 1628–1634. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, N.; Glover, E.I.; Phillips, S.M.; Isfort, R.J.; Tarnopolsky, M.A. Sex-based differences in skeletal muscle function and morphology with short-term limb immobilization. J. Appl. Physiol. 2005, 99, 1085–1092. [Google Scholar] [CrossRef]
- Hackney, K.J.; Ploutz-Snyder, L.L. Unilateral lower limb suspension: Integrative physiological knowledge from the past 20 years (1991–2011). Eur. J. Appl. Physiol. 2012, 112, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Saltin, B.; Henriksson, J.; Nygaard, E.; Andersen, P.; Jansson, E. Fiber types and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Ann. N. Y. Acad. Sci. 1977, 301, 3–29. [Google Scholar] [CrossRef] [PubMed]
- Haus, J.M.; Carrithers, J.A.; Carroll, C.C.; Tesch, P.A.; Trappe, T.A. Contractile and connective tissue protein content of human skeletal muscle: Effects of 35 and 90 days of simulated microgravity and exercise countermeasures. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1722–R1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeMoine, J.K.; Haus, J.M.; Trappe, S.W.; Trappe, T.A. Muscle proteins during 60-day bedrest in women: Impact of exercise or nutrition. Muscle Nerve 2009, 39, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Schoenrock, B.; Zander, V.; Dern, S.; Limper, U.; Mulder, E.; Veraksits, A.; Viir, R.; Kramer, A.; Stokes, M.J.; Salanova, M.; et al. Bed rest, exercise countermeasure and reconditioning effects on the human resting muscle tone system. Front. Physiol. 2018, 9, 810. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Huenchullan, S.; Mclennan, S.V.; Verhoeven, A.; Twigg, S.M.; Tam, C.S. The emerging role of skeletal muscle extracellular matrix remodelling in obesity and exercise. Obes. Rev. 2017, 18, 776–790. [Google Scholar] [CrossRef]
- Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal muscle extracellular matrix—What do we know about its composition, regulation, and physiological roles? A narrative review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- Whitham, M.; Parker, B.L.; Friedrichsen, M.; Hingst, J.R.; Hjorth, M.; Hughes, W.E.; Egan, C.L.; Cron, L.; Watt, K.I.; Kuchel, R.P.; et al. Extracellular vesicles provide a means for tissue crosstalk during exercise. Cell Metab. 2018, 27, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, H.; Deminice, R.; Yoshihara, T.; Powers, S.K. Mitochondrial dysfunction induces muscle atrophy during prolonged inactivity: A review of the causes and effects. Arch. Biochem. Biophys. 2019, 662, 49–60. [Google Scholar] [CrossRef]
- Lanza, I.R.; Short, D.K.; Short, K.R.; Raghavakaimal, S.; Basu, R.; Joyner, M.J.; McConnell, J.P.; Nair, K.S. Endurance exercise as a countermeasure for aging. Diabetes 2008, 57, 2933–2942. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Jain, S.S.; McFarlan, J.T.; Snook, L.A.; Chabowski, A.; Bonen, A. Exercise- and training-induced upregulation of skeletal muscle fatty acid oxidation are not solely dependent on mitochondrial machinery and biogenesis. J. Physiol. 2013, 591, 4415–4426. [Google Scholar] [CrossRef] [PubMed]
- Narici, M.V.; de Boer, M.D. Disuse of the musculo-skeletal system in space and on earth. Eur. J. Appl. Physiol. 2011, 111, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Rudrappa, S.S.; Wilkinson, D.J.; Greenhaff, P.L.; Smith, K.; Idris, I.; Atherton, P.J. Human skeletal muscle disuse atrophy: Effects on muscle protein synthesis, breakdown, and insulin resistance-A qualitative review. Front. Physiol. 2016, 7, 361. [Google Scholar] [CrossRef] [PubMed]
- Crossland, H.; Skirrow, S.; Puthucheary, Z.A.; Constantin-Teodosiu, D.; Greenhaff, P.L. The impact of immobilisation and inflammation on the regulation of muscle mass and insulin resistance: Different routes to similar end-points. J. Physiol. 2019, 597, 1259–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murgia, M.; Nagaraj, N.; Deshmukh, A.S.; Zeiler, M.; Cancellara, P.; Moretti, I.; Reggiani, C.; Schiaffino, S.; Mann, M. Single muscle fiber proteomics reveals unexpected mitochondrial specialization. EMBO Rep. 2015, 16, 387–395. [Google Scholar] [CrossRef]
- Holm, L.; Van, H.G.; Rose, A.J.; Miller, B.F.; Doessing, S.; Richter, E.A.; Kjaer, M. Contraction intensity and feeding affect collagen and myofibrillar protein synthesis rates differently in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E257–E269. [Google Scholar] [CrossRef] [Green Version]
- Karamanidis, K.; Arampatzis, A. Mechanical and morphological properties of human quadriceps femoris and triceps surae muscle-tendon unit in relation to aging and running. J. Biomech. 2006, 39, 406–417. [Google Scholar] [CrossRef]
- Kubo, K.; Kanehisa, H.; Kawakami, Y.; Fukunaga, T. Elastic properties of muscle-tendon complex in long-distance runners. Eur. J. Appl. Physiol. 2000, 81, 181–187. [Google Scholar] [CrossRef]
- Noakes, T.D. Effect of exercise on serum enzyme activities in humans. Sports Med. 1987, 4, 245–267. [Google Scholar] [CrossRef]
- McKee, T.J.; Perlman, G.; Morris, M.; Komarova, S.V. Extracellular matrix composition of connective tissues: A systematic review and meta-analysis. Sci. Rep. 2019, 9, 10542. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, T. Vascular remodelling in human skeletal muscle. Biochem. Soc. Trans. 2011, 39, 1628–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arentson-Lantz, E.J.; English, K.L.; Paddon-Jones, D.; Fry, C.S. Fourteen days of bed rest induces a decline in satellite cell content and robust atrophy of skeletal muscle fibers in middle-aged adults. J. Appl. Physiol. 2016, 120, 965–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigelso, A.; Gram, M.; Wiuff, C.; Andersen, J.L.; Helge, J.W.; Dela, F. Six weeks’ aerobic retraining after two weeks’ immobilization restores leg lean mass and aerobic capacity but does not fully rehabilitate leg strength in young and older men. J. Rehabil. Med. 2015, 47, 552–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, H.E.; Dudley, G.A.; Hather, B.; Tesch, P.A. Work capacity and metabolic and morphologic characteristics of the human quadriceps muscle in response to unloading. Clin. Physiol. 1993, 13, 337–347. [Google Scholar] [CrossRef]
- Hather, B.M.; Adams, G.R.; Tesch, P.A.; Dudley, G.A. Skeletal muscle responses to lower limb suspension in humans. J. Appl. Physiol. 1992, 72, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Rudnick, J.; Puttmann, B.; Tesch, P.A.; Alkner, B.; Schoser, B.G.; Salanova, M.; Kirsch, K.; Gunga, H.C.; Schiffl, G.; Luck, G.; et al. Differential expression of nitric oxide synthases (NOS 1-3) in human skeletal muscle following exercise countermeasure during 12 weeks of bed rest. FASEB J. 2004, 18, 1228–1230. [Google Scholar] [CrossRef]
- Salanova, M.; Schiffl, G.; Puttmann, B.; Schoser, B.G.; Blottner, D. Molecular biomarkers monitoring human skeletal muscle fibres and microvasculature following long-term bed rest with and without countermeasures. J. Anat. 2008, 212, 306–318. [Google Scholar] [CrossRef]
- Olfert, I.M.; Baum, O.; Hellsten, Y.; Egginton, S. Advances and challenges in skeletal muscle angiogenesis. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H326–H336. [Google Scholar] [CrossRef] [Green Version]
- Arner, E.; Daub, C.O.; Vitting-Seerup, K.; Andersson, R.; Lilje, B.; Drablos, F.; Lennartsson, A.; Ronnerblad, M.; Hrydziuszko, O.; Vitezic, M.; et al. Transcribed enhancers lead waves of coordinated transcription in transitioning mammalian cells. Science 2015, 347, 1010–1014. [Google Scholar] [CrossRef] [Green Version]
- Migliavacca, E.; Tay, S.K.H.; Patel, H.P.; Sonntag, T.; Civiletto, G.; McFarlane, C.; Forrester, T.; Barton, S.J.; Leow, M.K.; Antoun, E.; et al. Mitochondrial oxidative capacity and NAD(+) biosynthesis are reduced in human sarcopenia across ethnicities. Nat. Commun. 2019, 10, 5808. [Google Scholar] [CrossRef]
- Donges, C.E.; Burd, N.A.; Duffield, R.; Smith, G.C.; West, D.W.; Short, M.J.; Mackenzie, R.; Plank, L.D.; Shepherd, P.R.; Phillips, S.M.; et al. Concurrent resistance and aerobic exercise stimulates both myofibrillar and mitochondrial protein synthesis in sedentary middle-aged men. J. Appl. Physiol. 2012, 112, 1992–2001. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.B.; Phillips, S.M.; Atherton, P.J.; Patel, R.; Yarasheski, K.E.; Tarnopolsky, M.A.; Rennie, M.J. Differential effects of resistance and endurance exercise in the fed state on signalling molecule phosphorylation and protein synthesis in human muscle. J. Physiol. 2008, 586, 3701–3717. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, D.M.; West, D.W.; Churchward-Venne, T.A.; Breen, L.; Baker, S.K.; Phillips, S.M. Influence of aerobic exercise intensity on myofibrillar and mitochondrial protein synthesis in young men during early and late postexercise recovery. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1025–E1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, Y.; Ni, M.; Cao, H.; Signer, R.A.J.; Li, D.; Li, M.; Gu, Z.; Hu, Z.; Dickerson, K.E.; et al. Regulation of mitochondrial biogenesis in erythropoiesis by mTORC1-mediated protein translation. Nat. Cell Biol. 2017, 19, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Sinvani, H.; Haimov, O.; Svitkin, Y.; Sonenberg, N.; Tamarkin-Ben-Harush, A.; Viollet, B.; Dikstein, R. Translational tolerance of mitochondrial genes to metabolic energy stress involves TISU and eIF1-eIF4GI cooperation in start codon selection. Cell Metab. 2015, 21, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Henstridge, D.C.; Bruce, C.R.; Drew, B.G.; Tory, K.; Kolonics, A.; Estevez, E.; Chung, J.; Watson, N.; Gardner, T.; Lee-Young, R.S.; et al. Activating HSP72 in rodent skeletal muscle increases mitochondrial number and oxidative capacity and decreases insulin resistance. Diabetes 2014, 63, 1881–1894. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Xu, L.; Alberobello, A.T.; Gavrilova, O.; Bagattin, A.; Skarulis, M.; Liu, J.; Finkel, T.; Mueller, E. Celastrol protects against obesity and metabolic dysfunction through activation of a HSF1-PGC1alpha transcriptional axis. Cell Metab. 2015, 22, 695–708. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, M.E.; Marabita, F.; Gomez-Cabrero, D.; Rundqvist, H.; Ekstrom, T.J.; Tegner, J.; Sundberg, C.J. An integrative analysis reveals coordinated reprogramming of the epigenome and the transcriptome in human skeletal muscle after training. Epigenetics 2014, 9, 1557–1569. [Google Scholar] [CrossRef]
- Tomiga, Y.; Ito, A.; Sudo, M.; Ando, S.; Eshima, H.; Sakai, K.; Nakashima, S.; Uehara, Y.; Tanaka, H.; Soejima, H.; et al. One week, but not 12 hours, of cast immobilization alters promotor DNA methylation patterns in the nNOS gene in mouse skeletal muscle. J. Physiol. 2019, 597, 5145–5159. [Google Scholar] [CrossRef]
- Nitert, M.D.; Dayeh, T.; Volkov, P.; Elgzyri, T.; Hall, E.; Nilsson, E.; Yang, B.T.; Lang, S.; Parikh, H.; Wessman, Y.; et al. Impact of an exercise intervention on DNA methylation in skeletal muscle from first-degree relatives of patients with type 2 diabetes. Diabetes 2012, 61, 3322–3332. [Google Scholar] [CrossRef] [Green Version]
- Hai, T.; Curran, T. Cross-family dimerization of transcription factors Fos/Jun and ATF/CREB alters DNA binding specificity. Proc. Natl. Acad. Sci. USA 1991, 88, 3720–3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, J.R.; Keating, A.E. Comprehensive identification of human bZIP interactions with coiled-coil arrays. Science 2003, 300, 2097–2101. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Martinez, J.A.; Reinke, A.W.; Bhimsaria, D.; Keating, A.E.; Ansari, A.Z. Combinatorial bZIP dimers display complex DNA-binding specificity landscapes. Elife 2017, 6, e19272. [Google Scholar] [CrossRef] [PubMed]
- Reinke, A.W.; Baek, J.; Ashenberg, O.; Keating, A.E. Networks of bZIP protein-protein interactions diversified over a billion years of evolution. Science 2013, 340, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altarejos, J.Y.; Montminy, M. CREB and the CRTC co-activators: Sensors for hormonal and metabolic signals. Nat. Rev. Mol. Cell Biol. 2011, 12, 141–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, N.E.; Kelly, K.A.; Hawkins, R.; Bramah-Lawani, M.; Amelio, A.L.; Nwachukwu, J.C.; Nettles, K.W.; Conkright, M.D. Creb coactivators direct anabolic responses and enhance performance of skeletal muscle. EMBO J. 2014, 33, 1027–1043. [Google Scholar] [CrossRef] [PubMed]
- Uchida, S.; Teubner, B.J.W.; Hevi, C.; Hara, K.; Kobayashi, A.; Dave, R.M.; Shintaku, T.; Jaikhan, P.; Yamagata, H.; Suzuki, T.; et al. CRTC1 Nuclear translocation following learning modulates memory strength via exchange of chromatin remodeling complexes on the Fgf1 gene. Cell Rep. 2017, 18, 352–366. [Google Scholar] [CrossRef]
- Tsai, L.N.; Ku, T.K.; Salib, N.K.; Crowe, D.L. Extracellular signals regulate rapid coactivator recruitment at AP-1 sites by altered phosphorylation of both CREB binding protein and c-jun. Mol. Cell Biol. 2008, 28, 4240–4250. [Google Scholar] [CrossRef] [Green Version]
- Kasper, L.H.; Lerach, S.; Wang, J.; Wu, S.; Jeevan, T.; Brindle, P.K. CBP/p300 double null cells reveal effect of coactivator level and diversity on CREB transactivation. EMBO J. 2010, 29, 3660–3672. [Google Scholar] [CrossRef]
- Xu, W.; Kasper, L.H.; Lerach, S.; Jeevan, T.; Brindle, P.K. Individual CREB-target genes dictate usage of distinct cAMP-responsive coactivation mechanisms. EMBO J. 2007, 26, 2890–2903. [Google Scholar] [CrossRef]
- Andersson, R.; Sandelin, A. Determinants of enhancer and promoter activities of regulatory elements. Nat. Rev. Genet. 2020, 21, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Akberdin, I.R.; Vertyshev, A.Y.; Pintus, S.S.; Popov, D.V.; Kolpakov, F.A. A mathematical model linking Ca2+-dependent signaling pathway and gene expression regulation in human skeletal muscle. Math. Biol. Bioinform. 2020, 15, 20–39. [Google Scholar] [CrossRef] [Green Version]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The human transcription factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popov, D.V.; Lysenko, E.A.; Vepkhvadze, T.F.; Kurochkina, N.S.; Maknovskii, P.A.; Vinogradova, O.L. Promoter-specific regulation of ppargc1a gene expression in human skeletal muscle. J. Mol. Endocrinol. 2015, 55, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Metabolic Pathway/Protein Class | D | TR | TR + AE | AE 0 h | AE 0.5–1 h | AE 3 h | AE 4 h | ||
|---|---|---|---|---|---|---|---|---|---|
| Fiber type specific sarcomeric proteins/genes: 58 | Protein concentration | ↔↓ [34] | ↔↑ [34,35] | ↔↓ [36] | |||||
| MF1 CSA | ↓ [37,38,39,40,41] | ↔ [42] | |||||||
| MF2 CSA | ↓ [37,38,39,40,41] | ↔ [42] | |||||||
| Genes Up | 1 | 1 | |||||||
| Genes Down | 13 | 2 | 2 | ||||||
| MF1 specific | 12 | 1 | 1 | 1 | |||||
| MF2 specific | 1 | 1 | |||||||
| Extracellular matrix | Collagen proteins/genes: 27 | Protein concentration | ↔ [43,44,45] | ↑ [35,46] | |||||
| Protein content | ↓ | ↑ | |||||||
| Genes Up | 2 | 15 | 15 | 1 | |||||
| Genes Down | |||||||||
| ECM Glycoprotein proteins/genes: 122 | Protein concentration | ||||||||
| Genes Up | 26 | 31 | 1 | 4 | 2 | 1 | |||
| Genes Down | 4 | 1 | 2 | ||||||
| Proteoglycan proteins/genes: 19 | Protein concentration | ↔↑ [35] | |||||||
| Genes Up | 1 | 6 | 6 | 1 | |||||
| Genes Down | |||||||||
| ECM Regulator proteins/genes: 115 | Protein concentration | ↑ [46,47] | |||||||
| Genes Up | 19 | 26 | 3 | 6 | 6 | 2 | |||
| Genes Down | 2 | 1 | 1 | 1 | |||||
| Cytokine, growth and secreted factor proteins/genes: 179 | Protein concentration | ↑ [1,48] | |||||||
| Genes Up | 5 | 20 | 37 | 3 | 11 | 14 | 7 | ||
| Genes Down | 4 | 3 | 8 | 3 | 2 | 2 | 1 | ||
| Transcriptional regulator proteins/genes: 1527 | Protein concentration | ||||||||
| Genes Up | 6 | 11 | 130 | 42 | 66 | 61 | 26 | ||
| Genes Down | 4 | 21 | 58 | 10 | 7 | 16 | 16 | ||
| Glycolysis/ Gluconeogenesis proteins/genes: 35 | Protein concentration | ↔↓ [34,41,49] | ↔↑ [35,36,50] | ↔↓ [36] | |||||
| Genes Up | 1 | 4 | 1 | 3 | |||||
| Genes Down | 2 | ||||||||
| Lipid transport and beta oxidation proteins/genes: 26 | Protein concentration | ↓ [41,49] | ↑ [35,51] | ||||||
| Genes Up | 3 | 3 | |||||||
| Genes Down | 5 | ||||||||
| Mitochondrion | Citrate cycle (TCA cycle) proteins/genes: 27 | Protein concentration | ↓ [34,41,49] | ↑ [35,36,50] | ↔↓ [36] | ||||
| Genes Up | 1 | 1 | |||||||
| Genes Down | 3 | ||||||||
| Oxidative phosphorylation proteins/genes: 123 | Protein concentration | ↓ [34,41,49] | ↑ [35,36,50] | ↔↓ [36] | |||||
| Genes Up | 2 | 2 | 0 | ||||||
| Genes Down | 29 | ||||||||
| Mitochondrial proteins/genes: 1097 | Protein concentration | ↓ [34,41,49] | ↑ [35,36,50] | ↔↓ [36] | |||||
| Genes Up | 3 | 18 | 35 | 2 | 6 | 11 | 8 | ||
| Genes Down | 88 | 3 | 9 | 1 | 3 | 4 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makhnovskii, P.A.; Bokov, R.O.; Kolpakov, F.A.; Popov, D.V. Transcriptomic Signatures and Upstream Regulation in Human Skeletal Muscle Adapted to Disuse and Aerobic Exercise. Int. J. Mol. Sci. 2021, 22, 1208. https://doi.org/10.3390/ijms22031208
Makhnovskii PA, Bokov RO, Kolpakov FA, Popov DV. Transcriptomic Signatures and Upstream Regulation in Human Skeletal Muscle Adapted to Disuse and Aerobic Exercise. International Journal of Molecular Sciences. 2021; 22(3):1208. https://doi.org/10.3390/ijms22031208
Chicago/Turabian StyleMakhnovskii, Pavel A., Roman O. Bokov, Fedor A. Kolpakov, and Daniil V. Popov. 2021. "Transcriptomic Signatures and Upstream Regulation in Human Skeletal Muscle Adapted to Disuse and Aerobic Exercise" International Journal of Molecular Sciences 22, no. 3: 1208. https://doi.org/10.3390/ijms22031208