
DNA Methylation Signature for JARID2-Neurodevelopmental Syndrome

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
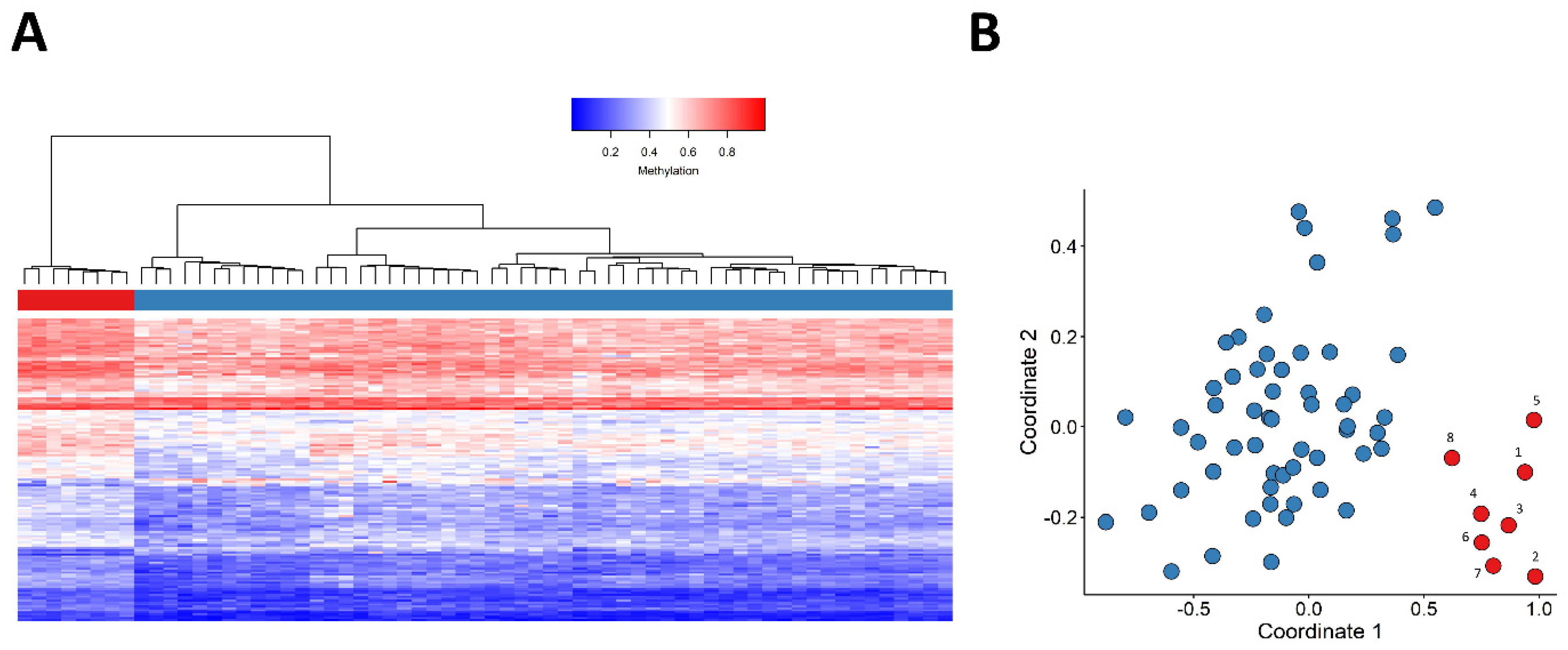
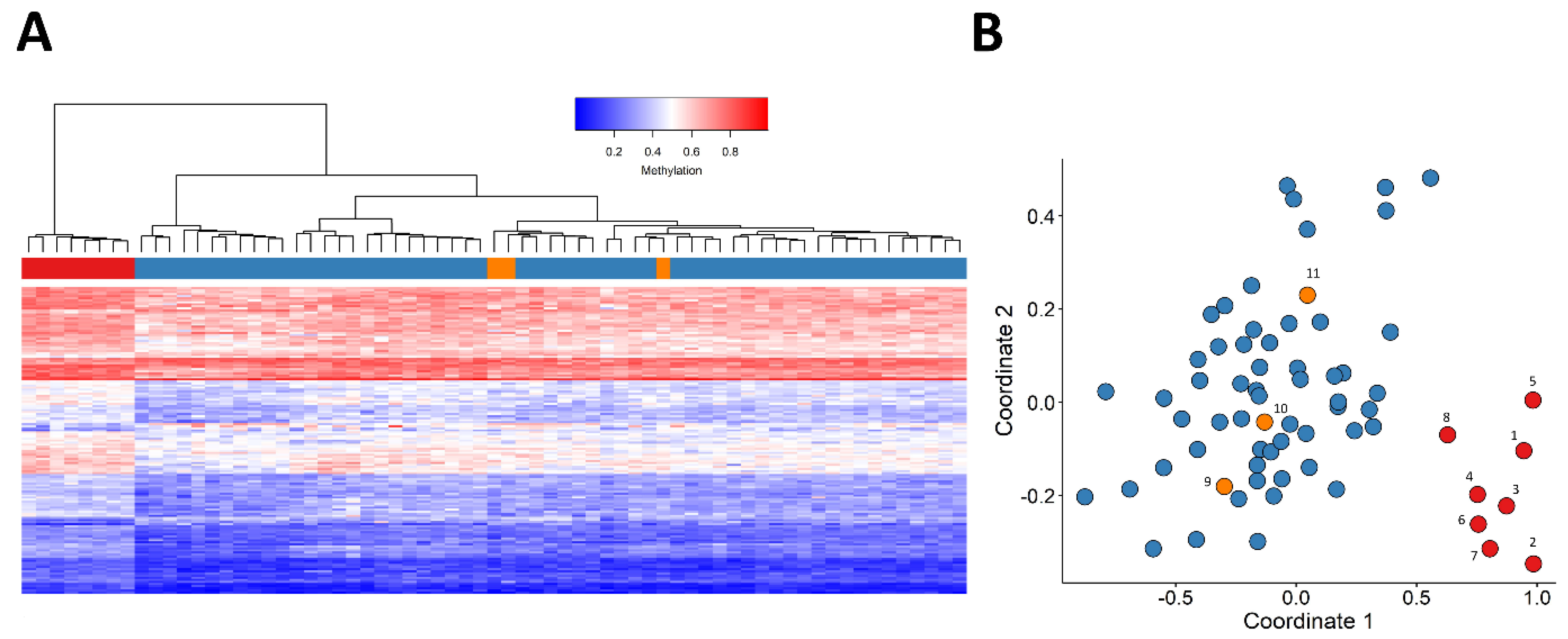
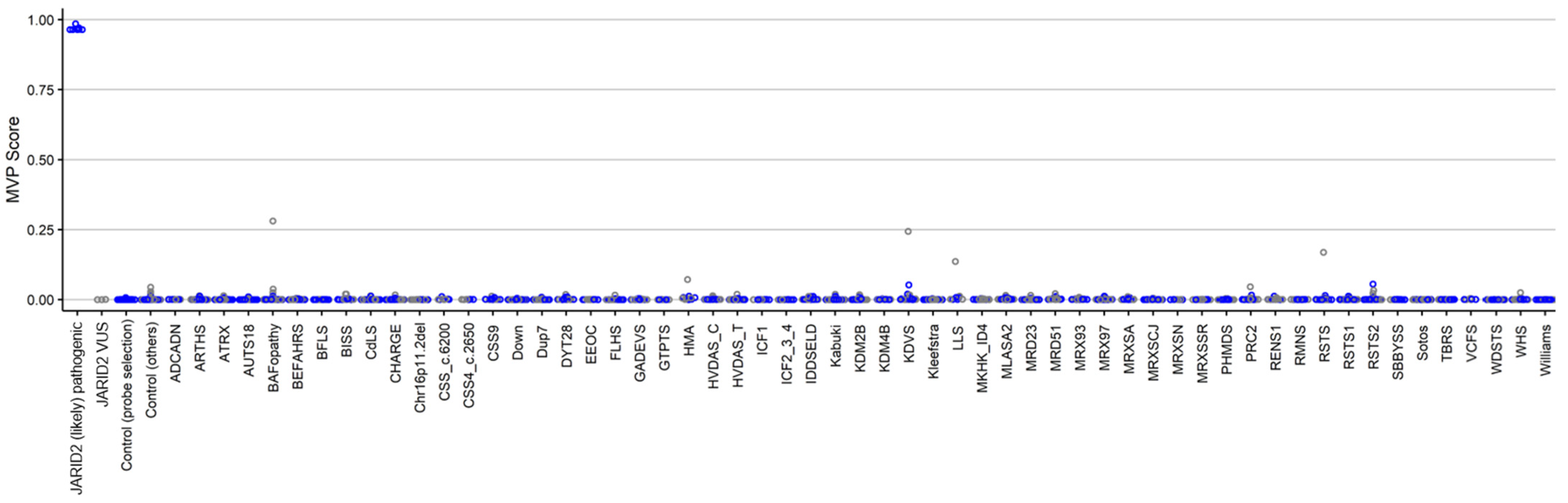
2.1. Identification and Assessment of an Episignature for the JARID2-Neurodevelopmental Syndrome
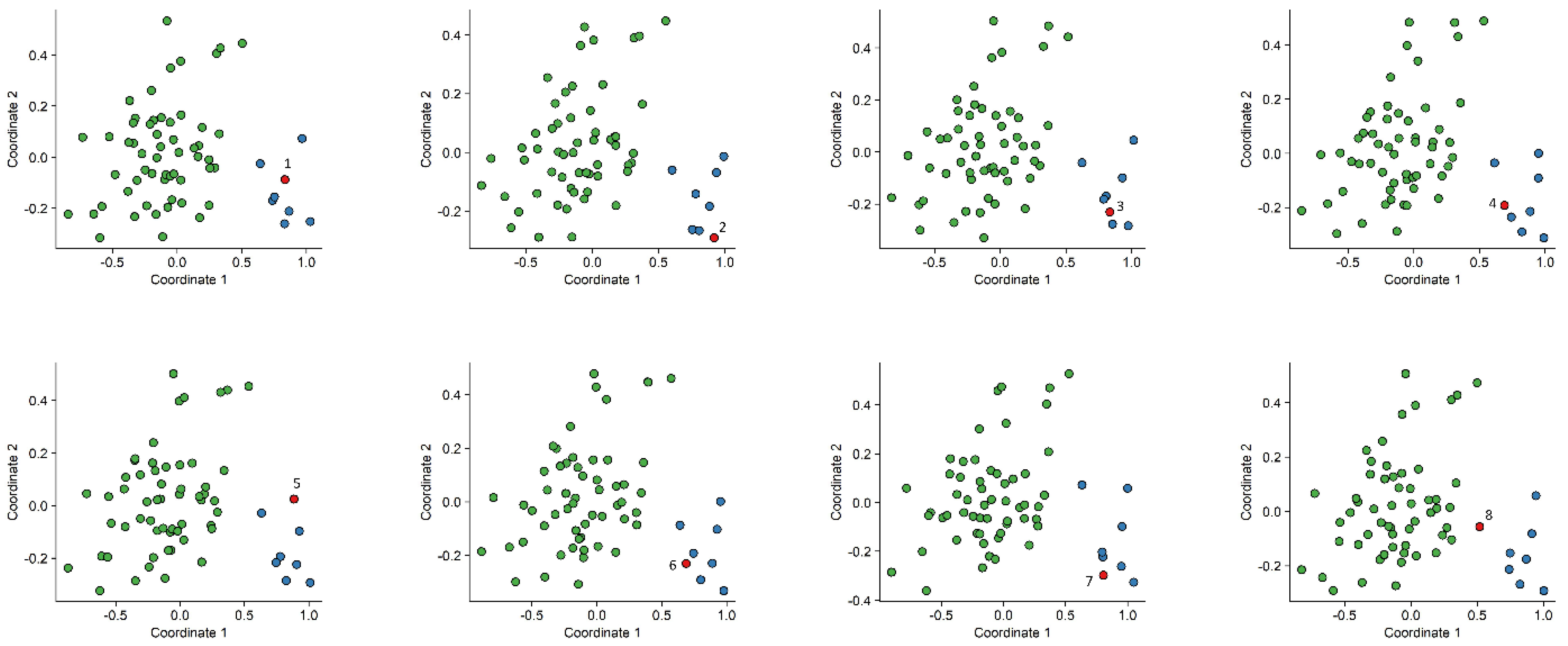
2.2. Construction of a Classification Model
2.3. Differentially Methylated Regions and Gene Set Enrichment Analysis
2.4. Gene Set Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Subjects and Study Cohort
4.2. DNA Isolation and Methylation Analysis
4.3. Quality Control of DNAm Profiles and Feature Selection
4.4. Construction of the Binary Classifier
4.5. Differentially Methylated Regions
4.6. Geneset Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Verberne, E.A.; Goh, S.; England, J.; van Ginkel, M.; Rafael-Croes, L.; Maas, S.; Polstra, A.; Zarate, Y.A.; Bosanko, K.A.; Pechter, K.B.; et al. JARID2 haploinsufficiency is associated with a clinically distinct neurodevelopmental syndrome. Genet. Med. 2021, 23, 374–383. [Google Scholar] [CrossRef]
- Barøy, T.; Misceo, D.; Strømme, P.; Stray-Pedersen, A.; Holmgren, A.; Rødningen, O.K.; Blomhoff, A.; Helle, J.R.; Stormyr, A.; Tvedt, B.; et al. Haploinsufficiency of two histone modifier genes on 6p22.3, ATXN1 and JARID2, is associated with intellectual disability. Orphanet J. Rare Dis. 2013, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Cadieux-Dion, M.; Farrow, E.; Thiffault, I.; Cohen, A.S.; Welsh, H.; Bartik, L.; Schwager, C.; Engleman, K.; Zhou, D.; Zhang, L.; et al. Phenotypic expansion and variable expressivity in individuals with JARID2-related intellectual disability: A case series. Clin. Genet. 2022, 102, 136–141. [Google Scholar] [CrossRef]
- Kasinath, V.; Beck, C.; Sauer, P.; Poepsel, S.; Kosmatka, J.; Faini, M.; Toso, D.; Aebersold, R.; Nogales, E. JARID2 and AEBP2 regulate PRC2 in the presence of H2AK119ub1 and other histone modifications. Science 2021, 371, eabc3393. [Google Scholar] [CrossRef] [PubMed]
- Pasini, D.; Cloos, P.A.; Walfridsson, J.; Olsson, L.; Bukowski, J.P.; Johansen, J.V.; Bak, M.; Tommerup, N.; Rappsilber, J.; Helin, K. JARID2 regulates binding of the Polycomb repressive complex 2 to target genes in ES cells. Nature 2010, 464, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Mysliwiec, M.R.; Carlson, C.D.; Tietjen, J.; Hung, H.; Ansari, A.Z.; Lee, Y. Jarid2 (Jumonji, AT rich interactive domain 2) regulates NOTCH1 expression via histone modification in the developing heart. J. Biol. Chem. 2012, 287, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aref-Eshghi, E.; Kerkhof, J.; Pedro, V.P.; France, G.D.; Barat-Houari, M.; Ruiz-Pallares, N.; Andrau, J.C.; Lacombe, D.; Van-Gils, J.; Fergelot, P.; et al. Evaluation of DNA methylation episignatures for diagnosis and phenotype correlations in 42 Mendelian neurodevelopmental disorders. Am. J. Hum. Genet. 2020, 106, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.A.; McConkey, H.; Kerkhof, J.; Barat-Houari, M.; Bargiacchi, S.; Biamino, E.; Bralo, M.P.; Cappuccio, G.; Ciolfi, A.; Clarke, A.; et al. Novel diagnostic DNA methylation episignatures expand and refine the epigenetic landscapes of Mendelian disorders. Hum. Genet. Genom. Adv. 2022, 3, 100075. [Google Scholar] [CrossRef]
- Bjornsson, H.T. The Mendelian disorders of the epigenetic machinery. Genome Res. 2015, 25, 1473–1481. [Google Scholar] [CrossRef] [Green Version]
- Aref-Eshghi, E.; Rodenhiser, D.I.; Schenkel, L.C.; Lin, H.; Skinner, C.; Ainsworth, P.; Paré, G.; Hood, R.L.; Bulman, D.E.; Kernohan, K.D.; et al. Genomic DNA Methylation Signatures Enable Concurrent Diagnosis and Clinical Genetic Variant Classification in Neurodevelopmental Syndromes. Am. J. Hum. Genet. 2018, 102, 156–174. [Google Scholar] [CrossRef] [Green Version]
- Aref-Eshghi, E.; Bend, E.G.; Colaiacovo, S.; Caudle, M.; Chakrabarti, R.; Napier, M.; Brick, L.; Brady, L.; Carere, D.A.; Levy, M.A.; et al. Diagnostic Utility of Genome-wide DNA Methylation Testing in Genetically Unsolved Individuals with Suspected Hereditary Conditions. Am. J. Hum. Genet. 2019, 104, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Peters, T.J.; Buckley, M.J.; Statham, A.L.; Pidsley, R.; Samaras, K.; Lord, R.V.; Clark, S.J.; Molloy, P.L. De novo identification of differentially methylated regions in the human genome. Epigenetics Chromatin 2015, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Sadikovic, B.; Aref-Eshghi, E.; Levy, M.A.; Rodenhiser, D. DNA methylation signatures in mendelian developmental disorders as a diagnostic bridge between genotype and phenotype. Epigenomics 2019, 11, 563–575. [Google Scholar] [CrossRef]
- Butcher, D.T.; Cytrynbaum, C.; Turinsky, A.L.; Siu, M.T.; Inbar-Feigenberg, M.; Mendoza-Londono, R.; Chitayat, D.; Walker, S.; Machado, J.; Caluseriu, O.; et al. CHARGE and Kabuki Syndromes: Gene-Specific DNA Methylation Signatures Identify Epigenetic Mechanisms Linking These Clinically Overlapping Conditions. Am. J. Hum. Genet. 2017, 100, 773–788. [Google Scholar] [CrossRef]
- Aref-Eshghi, E.; Schenkel, L.C.; Lin, H.; Skinner, C.; Ainsworth, P.; Paré, G.; Rodenhiser, D.; Schwartz, C.; Sadikovic, B. The defining DNA methylation signature of Kabuki syndrome enables functional assessment of genetic variants of unknown clinical significance. Epigenetics 2017, 12, 923–933. [Google Scholar] [CrossRef] [Green Version]
- Schenkel, L.C.; Schwartz, C.; Skinner, C.; Rodenhiser, D.I.; Ainsworth, P.J.; Pare, G.; Sadikovic, B. Clinical Validation of Fragile X Syndrome Screening by DNA Methylation Array. J. Mol. Diagn. 2016, 18, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Bend, E.G.; Aref-Eshghi, E.; Everman, D.B.; Rogers, R.C.; Cathey, S.S.; Prijoles, E.J.; Lyons, M.J.; Davis, H.; Clarkson, K.; Gripp, K.W.; et al. Gene domain-specific DNA methylation episignatures highlight distinct molecular entities of ADNP syndrome. Clin. Epigenetics 2019, 11, 64. [Google Scholar] [CrossRef]
- Sadikovic, B.; Levy, M.A.; Kerkhof, J.; Aref-Eshghi, E.; Schenkel, L.; Stuart, A.; McConkey, H.; Henneman, P.; Venema, A.; Schwartz, C.E.; et al. Clinical epigenomics: Genome-wide DNA methylation analysis for the diagnosis of Mendelian disorders. Genet. Med. 2021, 23, 1065–1074. [Google Scholar] [CrossRef]
- Grantham, R. Amino acid difference formula to help explain protein evolution. Science 1974, 185, 862–864. [Google Scholar] [CrossRef]
- Patsialou, A.; Wilsker, D.; Moran, E. DNA-binding properties of ARID family proteins. Nucleic Acids Res. 2005, 33, 66–80. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Margueron, R.; Ku, M.; Chambon, P.; Bernstein, B.E.; Reinberg, D. Jarid2 and PRC2, partners in regulating gene expression. Genes Dev. 2010, 24, 368–380. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, M.; Nomura, J.; Ji, X.; Tamada, K.; Arai, T.; Takahashi, E.; Bućan, M.; Takumi, T. Functional significance of rare neuroligin 1 variants found in autism. PLoS Genet. 2017, 13, e1006940. [Google Scholar] [CrossRef] [Green Version]
- Vezain, M.; Lecuyer, M.; Rubio, M.; Dupé, V.; Ratié, L.; David, V.; Pasquier, L.; Odent, S.; Coutant, S.; Tournier, I.; et al. A de novo variant in ADGRL2 suggests a novel mechanism underlying the previously undescribed association of extreme microcephaly with severely reduced sulcation and rhombencephalosynapsis. Acta Neuropathol. Commun. 2018, 6, 109. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [Green Version]
- Aref-Eshghi, E.; Bend, E.G.; Hood, R.L.; Schenkel, L.C.; Carere, D.A.; Chakrabarti, R.; Nagamani, S.; Cheung, S.W.; Campeau, P.M.; Prasad, C.; et al. BAFopathies’ DNA methylation epi-signatures demonstrate diagnostic utility and functional continuum of Coffin-Siris and Nicolaides-Baraitser syndromes. Nat. Commun. 2018, 9, 4885. [Google Scholar] [CrossRef]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef] [Green Version]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Collins, J.R.; Alvord, W.G.; Roayaei, J.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. The DAVID Gene Functional Classification Tool: A novel biological module-centric algorithm to functionally analyze large gene lists. Genome Biol. 2007, 8, R183. [Google Scholar] [CrossRef] [Green Version]
- WEB-Based Gene SeT AnaLysis Toolkit. WebGestalt (WEB-Based GEne SeT AnaLysis Toolkit). 2019. Available online: http://www.webgestalt.org (accessed on 1 March 2022).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient # | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variant | (15291644_15388348)x1 | (15330889_15419256)x1 | (15374392_15405436)x1 | c.2866dupG p.(Glu956GlyfsTer72) | c.2731 + 1G > C | (15383717_15462037)x1 | (15383717_15462037)x1 | deletion exons 6-18 * | c.2363G > A p.(Arg788Gln) | c.1930G > A p.(Glu644Lys) | c.2363G > A p.(Arg788Gln) |
| Genomic position (hg19) | - | - | - | (15511546dup) | (15507648G > C) | - | - | - | (15501555G > A) | (15497386G > A) | (15501555G > A) |
| Variant type | Del | Del | Del | FS | SS | Del | Del | Del | Mis | Mis | Mis |
| Inheritance | dn | dn | dn | dn | dn | mat | NA | dn | dn | dn | mat † |
| Classification | P | P | P | P | LP | P | P | P | VUS | VUS | VUS |
| Gender | M | F | F | M | M | F | F | M | F | M | M |
| Age (years) | 14 | 21 | 18 | 14 | 5 | 6 | 36 | 9 | 43 | 12 | 8 |
| Intellectual functioning | Mild ID (IQ 61-74) | Borderline intellectual functioning (IQ 82) | Mild ID (IQ 50) | Moderate ID(IQ NA) | IQ NA | Normal | Learning difficulties (IQ NA) | Mild ID (IQ NA) | Learning difficulties (IQ 79) | Mild ID (IQ 66) | Normal |
| Developmental delay | + | + | + | + | + | + | + | + | + | + | + |
| Behavior abnormalities | - | + | - | + | - | - | - | + | + | - | - |
| Autistic features | + | - | + | + | - | - | - | + | + | - | - |
| ASD diagnosis | - | - | - | + | - | - | - | + | + | - | - |
| Hypotonia | - | - | - | - | + | - | - | + (mild) | - | - | + (later spasticity) |
| Gait disturbance | - | - | - | - | - | + (previously rigid walking pattern) | - | + | - | - | + (due to spasticity) |
| MRI abnormalities | NA | NA | NA | Small posterior fossa cyst or mega cisterna magna | Arachnoid cyst | NA | NA | Normal spinal cord MRI | NA | NA | Brain MRI: lack of myelinization. Normal spinal MRI. |
| Dysmorphic features | |||||||||||
| - Broad forehead | + | - | - | - | + | - | - | + | - | - | + |
| - High anterior hair line | - | + | + | + | - | + | - | + | - | - | + |
| - Prominent supraorbital ridges | - | - | - | - | - | - | - | + | - | - | - |
| - Deeply set eyes | - | + | + | - | + | - | - | + | - | - | + |
| - Infraorbital dark circles | + | - | + | - | - | + | - | + | - | - | + |
| - Midface hypoplasia | - | + | - | - | - | - | - | - | - | - | + |
| - Depressed nasal bridge | - | - | + | - | - | - | - | slight | - | - | - |
| - Bulbous nasal tip | - | - | + | - | - | + | + | - | - | - | + |
| - Short philtrum | - | + | + | + | - | - | - | - | - | - | + |
| - Full lips | - | - | + | + | - | - | - | - | - | - | + |
| Other anomalies | Pes plano valgus, mild hypermetropia | Submucous cleft palate, bifid uvula | Fetal finger pads, slight tapering of digit II and V bilateral. | 2 café au lait macules | Right cryptorchidism, congenital torticollis | Supernumerary tooth | - | Kyfoscoliosis, bladder spasticity | Strabismus convergens, camptodactyly digiti V of the hands, syndactyly dig 2–3 of the feet | Severe global spasticity, neurogenic bladder |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verberne, E.A.; van der Laan, L.; Haghshenas, S.; Rooney, K.; Levy, M.A.; Alders, M.; Maas, S.M.; Jansen, S.; Lieden, A.; Anderlid, B.-M.; et al. DNA Methylation Signature for JARID2-Neurodevelopmental Syndrome. Int. J. Mol. Sci. 2022, 23, 8001. https://doi.org/10.3390/ijms23148001
Verberne EA, van der Laan L, Haghshenas S, Rooney K, Levy MA, Alders M, Maas SM, Jansen S, Lieden A, Anderlid B-M, et al. DNA Methylation Signature for JARID2-Neurodevelopmental Syndrome. International Journal of Molecular Sciences. 2022; 23(14):8001. https://doi.org/10.3390/ijms23148001
Chicago/Turabian StyleVerberne, Eline A., Liselot van der Laan, Sadegheh Haghshenas, Kathleen Rooney, Michael A. Levy, Mariëlle Alders, Saskia M. Maas, Sandra Jansen, Agne Lieden, Britt-Marie Anderlid, and et al. 2022. "DNA Methylation Signature for JARID2-Neurodevelopmental Syndrome" International Journal of Molecular Sciences 23, no. 14: 8001. https://doi.org/10.3390/ijms23148001