
Efficiency of Orexin-A for Inflammatory Flare and Mucosal Healing in Experimental Colitis: Comparison with the Anti-TNF Alpha Infliximab
 , ,
, ,
Abstract
:1. Introduction
2. Results
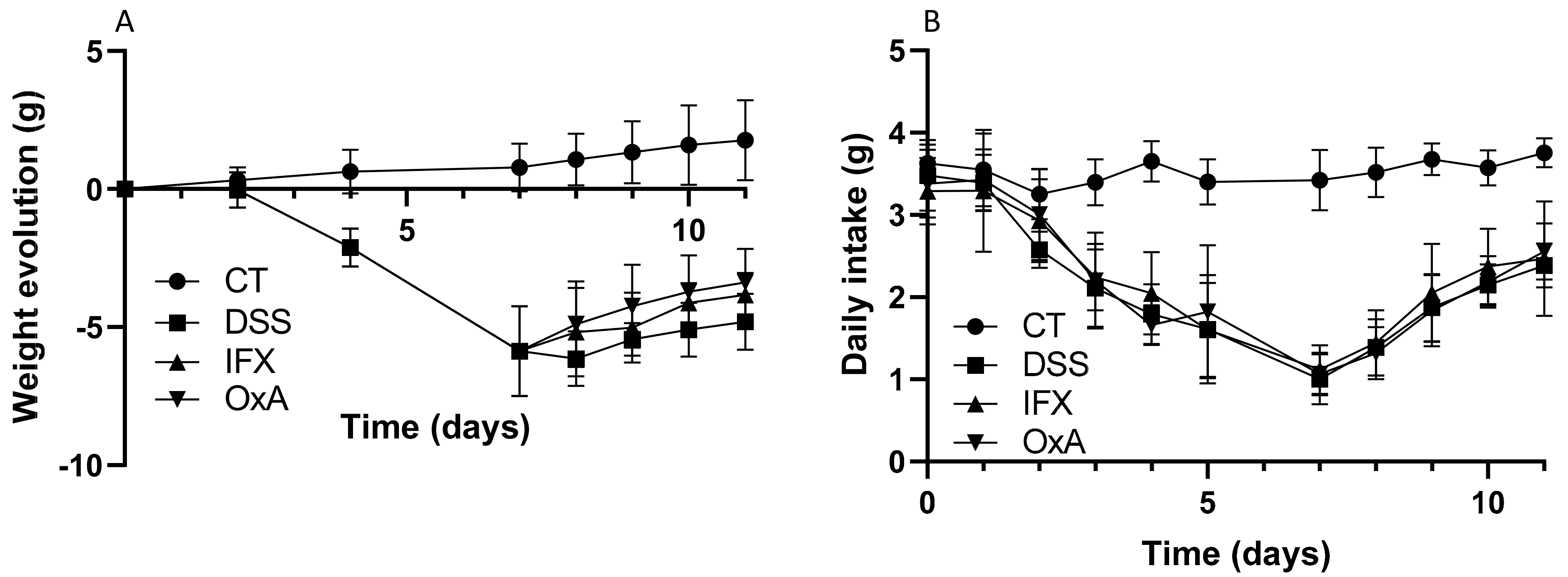
2.1. Effect of DSS, IFX and OxA on Body Weight
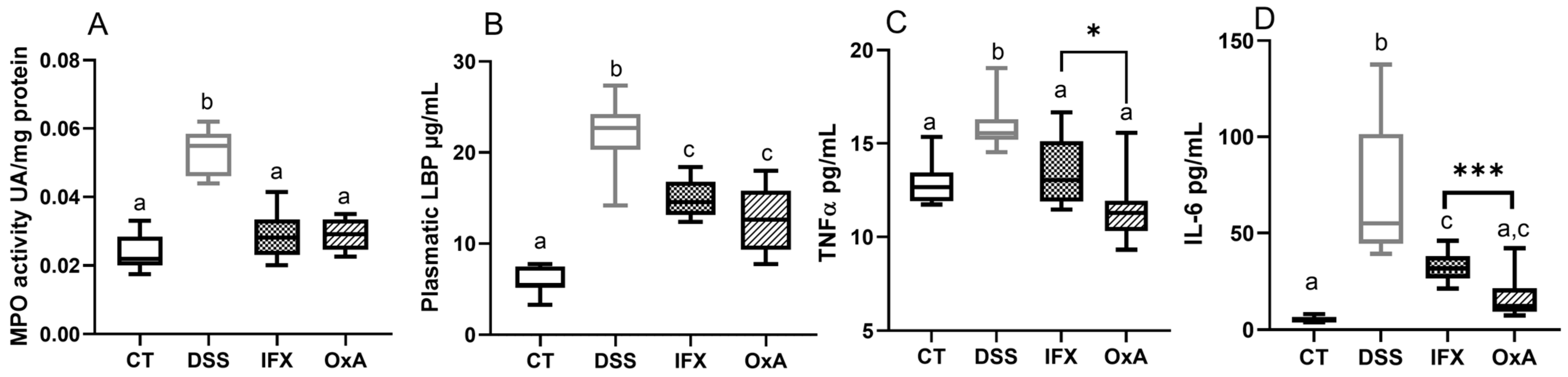
2.2. Effect of DSS, IFX and OxA on Inflammatory Markers
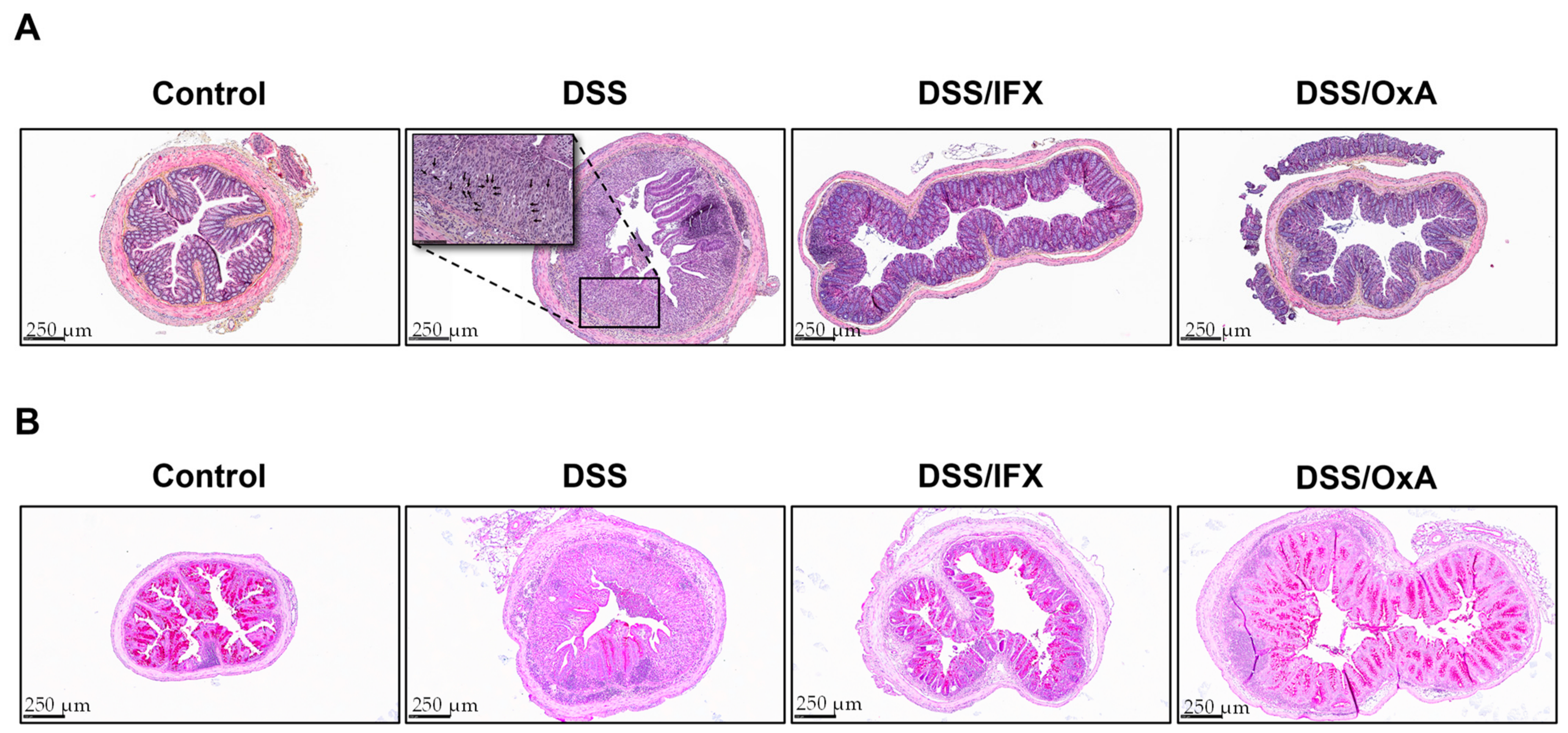
2.3. Histological Changes Induced by DSS and Effects of IFX and OxA Treatments
2.4. Effect of DSS, IFX and OxA on Colonic mRNA Expression of Different Genes Involved in Colonic Inflammation and Healing
3. Discussion
4. Materials and Methods
4.1. Animals
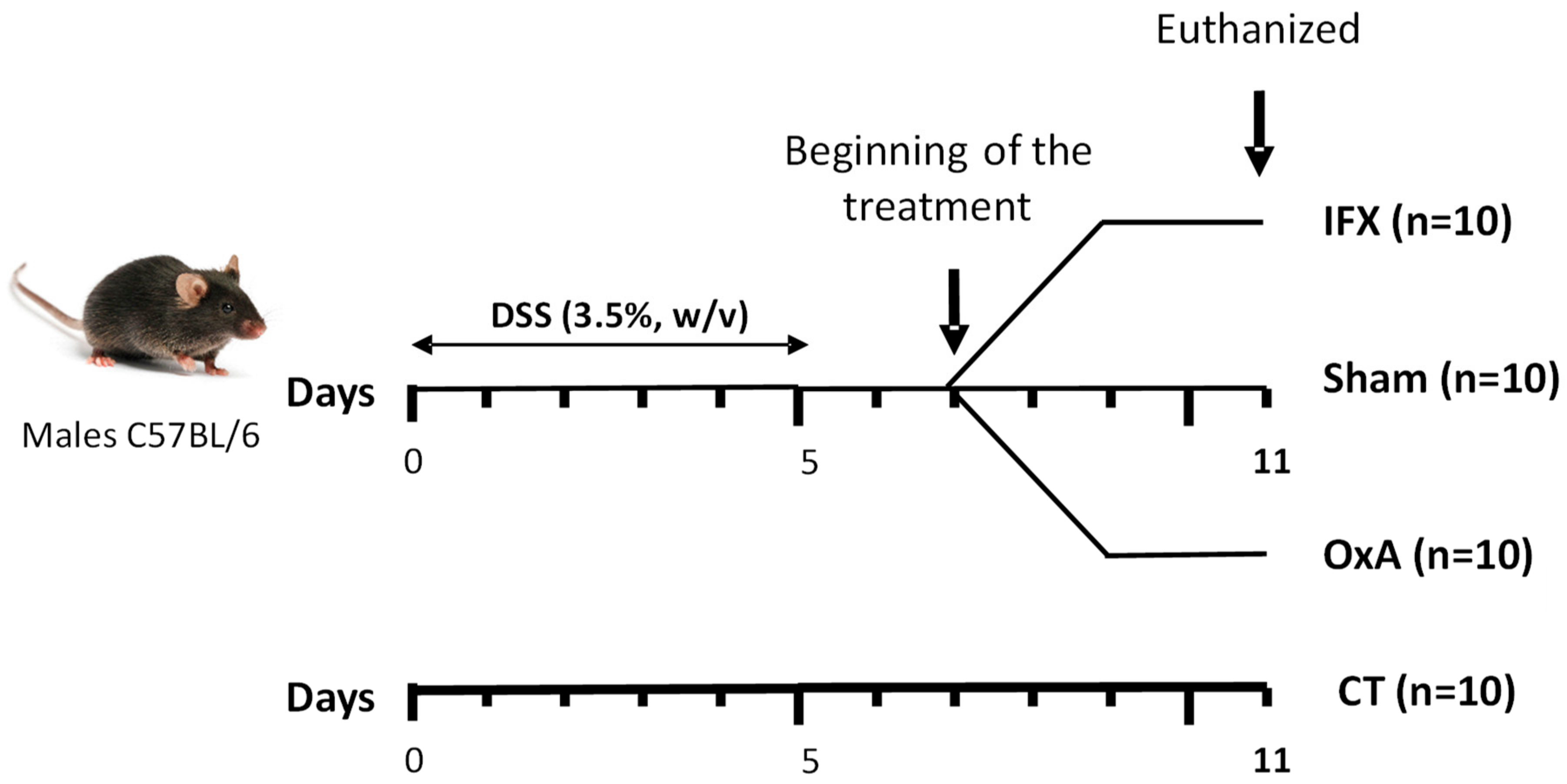
4.2. Experimental Design
4.3. Body Composition
4.4. Tissue Collection
4.5. Quantification of Gene Expression via Real-Time Polymerase Chain Reaction (qRTPCR)
4.6. Histological Analysis
4.7. Determination of Local and Systemic Inflammatory Markers
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Démaris, A.; Widigson, E.S.K.; Ilvemark, J.F.K.F.; Steenholdt, C.; Seidelin, J.B.; Huisinga, W.; Michelet, R.; Aulin, L.B.S.; Kloft, C. Ulcerative Colitis and Acute Severe Ulcerative Colitis Patients Are Overlooked in Infliximab Population Pharmacokinetic Models: Results from a Comprehensive Review. Pharmaceutics 2022, 14, 2095. [Google Scholar] [CrossRef] [PubMed]
- Nakase, H.; Hirano, T.; Wagatsuma, K.; Ichimiya, T.; Yamakawa, T.; Yokoyama, Y.; Hayashi, Y.; Hirayama, D.; Kazama, T.; Yoshii, S.; et al. Artificial intelligence-assisted endoscopy changes the definition of mucosal healing in ulcerative colitis. Dig. Endosc. 2021, 33, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Nakase, H. Optimizing the Use of Current Treatments and Emerging Therapeutic Approaches to Achieve Therapeutic Success in Patients with Inflammatory Bowel Disease. Gut Liver 2020, 14, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Hindryckx, P.; Jairath, V.; D’Haens, G. Acute severe ulcerative colitis: From pathophysiology to clinical management. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 654–664. [Google Scholar] [CrossRef]
- Fernández-Clotet, A.; Castro-Poceiro, J.; Panés, J. JAK Inhibition: The Most Promising Agents in the IBD Pipeline? Curr. Pharm. Des. 2019, 25, 32–40. [Google Scholar] [CrossRef]
- Hemperly, A.; Vande Casteele, N. Clinical Pharmacokinetics and Pharmacodynamics of Infliximab in the Treatment of Inflammatory Bowel Disease. Clin. Pharmacokinet. 2018, 57, 929–942. [Google Scholar] [CrossRef]
- Couvineau, A.; Dayot, S.; Nicole, P.; Gratio, V.; Rebours, V.; Couvelard, A.; Voisin, T. The Anti-tumoral Properties of Orexin/Hypocretin Hypothalamic Neuropeptides: An Unexpected Therapeutic Role. Front. Endocrinol. 2018, 27, 573. [Google Scholar] [CrossRef]
- Jacobson, L.H.; Hoyer, D.; de Lecea, L. Hypocretins (orexins): The ultimate translational neuropeptides. J. Intern. Med. 2022, 291, 533–556. [Google Scholar] [CrossRef]
- Kukkonen, J.P.; Turunen, P.M. Cellular Signaling Mechanisms of Hypocretin/Orexin. Front. Neurol. Neurosci. 2021, 45, 91–102. [Google Scholar] [CrossRef]
- Couvineau, A.; Voisin, T.; Nicole, P.; Gratio, V.; Abad, C.; Tan, Y.V. Orexins as Novel Therapeutic Targets in Inflammatory and Neurodegenerative Diseases. Front. Endocrinol. 2019, 22, 709. [Google Scholar] [CrossRef]
- Messal, N.; Fernandez, N.; Dayot, S.; Gratio, V.; Nicole, P.; Prochasson, C.; Chantret, I.; LeGuilloux, G.; Jarry, A.; Couvelard, A.; et al. Ectopic expression of OX1R in ulcerative colitis mediates anti-inflammatory effect of orexin-A. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3618–3628. [Google Scholar] [CrossRef] [PubMed]
- Voisin, T.; El Firar, A.; Fasseu, M.; Rouyer-Fessard, C.; Descatoire, V.; Walker, F.; Paradis, V.; Bedossa, P.; Henin, D.; Lehy, T.; et al. Aberrant expression of OX1 receptors for orexins in colon cancers and liver metastases: An openable gate to apoptosis. Cancer Res. 2011, 71, 3341–3351. [Google Scholar] [CrossRef] [PubMed]
- Becquet, L.; Abad, C.; Leclercq, M.; Miel, C.; Jean, L.; Riou, G.; Couvineau, A.; Boyer, O.; Tan, Y.V. Systemic administration of orexin A ameliorates established experimental autoimmune encephalomyelitis by diminishing neuroinflammation. J. Neuroinflammation 2019, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Irukayama-Tomobe, Y.; Murakoshi, N.; Kiyama, M.; Ishikawa, Y.; Hosokawa, N.; Tominaga, H.; Uchida, S.; Kimura, S.; Kanuka, M.; et al. Peripherally administered orexin improves survival of mice with endotoxin shock. eLife 2016, 30, e21055. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Lletjós, S.; Andriamihaja, M.; Blais, A.; Grauso, M.; Lepage, P.; Davila, A.M.; Gaudichon, C.; Leclerc, M.; Blachier, F.; Lan, A. Mucosal healing progression after acute colitis in mice. World J. Gastroenterol. 2019, 25, 3572–3589. [Google Scholar] [CrossRef] [PubMed]
- Eichele, E.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef] [PubMed]
- Katsandegwaza, B.; Horsnell, W.; Katherine Smith, K. Inflammatory Bowel Disease: A Review of Pre-Clinical Murine Models of Human Disease. Int. J. Mol. Sci. 2022, 23, 9344. [Google Scholar] [CrossRef]
- Gajendran, M.; Loganathan, P.; Jimenez, G.; Catinella, A.P.; Ng, N.; Umapathy, C.; Ziade, N.; Hashash, J.G. A comprehensive review and update on ulcerative colitis. Dis. Mon. 2019, 65, 100851. [Google Scholar] [CrossRef]
- Knight, D.M.; Trinh, H.; Le, J.; Siegel, S.; Shealy, D.; McDonough, M.; Scallon, B.; Moore, M.A.; Vilcek, J.; Daddona, P.; et al. Construction and initial characterization of a mouse-human chimeric anti-TNF antibody. Mol. Immunol. 1993, 30, 1443–1453. [Google Scholar] [CrossRef]
- Derkx, B.; Taminiau, J.; Radema, S.; Stronkhorst, A.; Wortel, C.; Tytgat, G.; van Deventer, S. Tumour-necrosis-factor antibody treatment in Crohn’s disease. Lancet 1993, 342, 173–174. [Google Scholar] [CrossRef]
- Nalagatla, N.; Falloon, K.; Tran, G.; Borren, N.Z.; Avalos, D.; Luther, J.; Colizzo, F.; Garber, J.; Khalili, H.; Melia, J.; et al. Effect of Accelerated Infliximab Induction on Short- and Long-term Outcomes of Acute Severe Ulcerative Colitis: A Retrospective Multicenter Study and Meta-analysis. Clin. Gastroenterol. Hepatol. 2019, 17, 502–509.e1. [Google Scholar] [CrossRef] [PubMed]
- Atreya, R.; Neurath, M.F.; Siegmund, B. Personalizing Treatment in IBD: Hype or Reality in 2020? Can We Predict Response to Anti-TNF? Front. Med. 2020, 7, 517. [Google Scholar] [CrossRef]
- Stidham, R.W.; Lee, T.C.; Higgins, P.D.; Deshpande, A.R.; Sussman, D.A.; Singal, A.G.; Elmunzer, B.J.; Saini, S.D.; Vijan, S.; Waljee, A.K. Systematic review with network meta-analysis: The efficacy of anti-tumour necrosis factor-alpha agents for the treatment of ulcerative colitis. Aliment. Pharmacol. Ther. 2014, 39, 660–671. [Google Scholar] [CrossRef]
- Arihara, Z.; Takahashi, K.; Murakami, O.; Totsune, K.; Sone, M.; Satoh, F.; Ito, S.; Mouri, T. Immunoreactive orexin-A in human plasma. Peptides 2001, 22, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, S.; Nishijima, T.; Takahashi, S.; Yamauchi, K.; Arihara, Z.; Takahashi, K. Clinical significance of daytime plasma orexin-A-like immunoreactivity concentrations in patients with obstructive sleep apnea hypopnea syndrome. Respiration 2004, 71, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Nicole, P.; Couvineau, P.; Jamin, N.; Voisin, T.; Couvineau, A. Crucial role of the orexin-B C-terminus in the induction of OX1 receptor-mediated apoptosis: Analysis by alanine scanning, molecular modelling and site-directed mutagenesis. Br. J. Pharmacol. 2015, 172, 5211–5223. [Google Scholar] [CrossRef] [PubMed]
- Sokołowska, P.; Urbańska, A.; Namiecińska, M.; Biegańska, K.; Zawilska, J.B. Orexins promote survival of rat cortical neurons. Neurosci. Lett. 2012, 506, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.D.; Wildenberg, M.E.; van den Brink, G.R. Mechanism of Action of Anti-TNF Therapy in Inflammatory Bowel Disease. J. Crohns Colitis 2016, 10, 989–997. [Google Scholar] [CrossRef]
- Peake, S.T.; Bernardo, D.; Mann, E.R.; Al-Hassi, H.O.; Knight, S.C.; Hart, A.L. Infliximab induces a dysregulated tissue-homing profile on human T-lymphocytes in-vitro: A novel mechanism for paradoxical inflammation? J. Crohns Colitis 2013, 7, 765–767. [Google Scholar] [CrossRef]
- Horiuchi, T.; Mitoma, H.; Harashima, S.; Tsukamoto, H.; Shimoda, T. Transmembrane TNF-alpha: Structure, function and interaction with anti-TNF agents. Rheumatology 2010, 49, 1215–1228. [Google Scholar] [CrossRef]
- ten Hove, T.; van Montfrans, C.; Peppelenbosch, M.P.; van Deventer, S.J. Infliximab treatment induces apoptosis of lamina propria T lymphocytes in Crohn’s disease. Gut 2002, 50, 206–211. [Google Scholar] [CrossRef]
- van den Brande, J.M.; Peppelenbosch, M.P.; Hommes, D.W. Synergistic effect of methotrexate and infliximab on activated lymphocyte apoptosis. Inflamm. Bowel Dis. 2007, 13, 118–119. [Google Scholar] [CrossRef] [PubMed]
- Vos, A.C.; Wildenberg, M.E.; Arijs, I.; Duijvestein, M.; Verhaar, A.P.; de Hertogh, G.; Vermeire, S.; Rutgeerts, P.; van den Brink, G.R.; Hommes, D.W. Regulatory macrophages induced by infliximab are involved in healing in vivo and in vitro. Inflamm. Bowel Dis. 2012, 18, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Assas, M.B.; Levison, S.E.; Little, M.; England, H.; Battrick, L.; Bagnall, J.; McLaughlin, J.T.; Paszek, P.; Else, K.J.; Pennock, J.L. Anti-inflammatory effects of infliximab in mice are independent of tumour necrosis factor α neutralization. Clin. Exp. Immunol. 2017, 187, 225–233. [Google Scholar] [CrossRef]
- Kalliolias, G.D.; Ivashkiv, L.B. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Radu, A.F.; Bungau, S.G. Nanomedical approaches in the realm of rheumatoid arthritis. Ageing Res. Rev. 2023, 87, 101927. [Google Scholar] [CrossRef]
- Couvineau, A.; Nicole, P.; Gratio, V.; Voisin, T. The Orexin receptors: Structural and anti-tumoral properties. Front. Endocrinol. 2022, 13, 931970. [Google Scholar] [CrossRef]
- Putula, J.; Pihlajamaa, T.; Kukkonen, J.P. Calcium affects OX1 orexin (hypocretin) receptor responses by modifying both orexin binding and the signal transduction machinery. Br. J. Pharmacol. 2014, 171, 5816–5828. [Google Scholar] [CrossRef]
- Lee, J.W.J.; Plichta, D.; Hogstrom, L.; Borren, N.Z.; Lau, H.; Gregory, S.M.; Tan, W.; Khalili, H.; Clish, C.; Vlamakis, H.; et al. Multi-omics reveal microbial determinants impacting responses to biologic therapies in inflammatory bowel disease. Cell Host Microbe 2021, 29, 1294–1304.e4. [Google Scholar] [CrossRef]
- Pavel, F.M.; Vesa, C.M.; Gheorghe, G.; Diaconu, C.C.; Stoicescu, M.; Munteanu, M.A.; Babes, E.E.; Tit, D.M.; Toma, M.M.; Bungau, S. Highlighting the Relevance of Gut Microbiota Manipulation in Inflammatory Bowel Disease. Diagnostics 2021, 11, 1090. [Google Scholar] [CrossRef]
- Blais, A.; Drouin, G.; Chaumontet, C.; Voisin, T.; Couvelard, A.; Even, P.C.; Couvineau, A. Impact of Orexin-A Treatment on Food Intake, Energy Metabolism and Body Weight in Mice. PLoS ONE 2017, 12, e0169908. [Google Scholar] [CrossRef] [PubMed]
- Lan, A.; Andriamihaja, M.; Blouin, J.M.; Liu, X.; Descatoire, V.; Desclee de Maredsous, C.; Davila, A.M.; Walker, F.; Tome, D.; Blachier, F. High-protein diet differently modifies intestinal goblet cell characteristics and mucosal cytokine expression in ileum and colon. J. Nutr. Biochem. 2015, 6, 91–98. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | CT | DSS | IFX | OxA |
|---|---|---|---|---|
| Inflammatory markers | ||||
| Tnf-a | 1.09 ± 0.16 a | 3.55 ± 0.30 b | 1.86 ± 0.28 a | 1.93 ± 0.36 a |
| Il-6 | 1.03 ± 0.12 a | 11.17 ± 0.70 b | 5.21 ± 1.11 c | 7.53 ± 0.65 c |
| Il-10 | 1.07 ± 0.07 a | 11.44 ± 1.26 b | 4.81 ± 0.63 c | 3.59 ± 0.57 a,c |
| Il-13 | 1.08 ± 0.08 a | 0.46 ± 0.05 b | 1.17 ± 0.08 a | 0.91 ± 0.13 a |
| Tight-junction protein | ||||
| Tjp1 | 1.01 ± 0.16 a,b | 0.60 ± 0.05 a | 1.04 ± 0.07 a,b | 1.15 ± 0.22 b |
| Ocln | 1.03 ± 0.07 a | 0.48 ± 0.04 b | 1.02 ± 0.17 a | 1.06 ± 0.23 a |
| Cldn2 | 1.06 ± 0.15 | 1.51 ± 0.30 | 1.83 ± 0.16 | 1.35 ± 0.32 |
| Goblet cell markers and mucins | ||||
| Muc2 | 0.99 ± 0.09 a | 0.37 ± 0.04 b | 1.02 ± 0.15 a | 0.87 ± 0.12 a |
| Klf4 | 1.00 ± 0.06 a | 0.42 ± 0.06 b | 0.74 ± 0.09 a | 0.74 ± 0.08 a |
| Epithelial-repair-modulating factors | ||||
| Igf1 | 1.09 ± 0.18 a | 4.59 ± 1.05 b | 2.10 ± 0.34 a | 2.15 ± 0.25 a |
| Il-15 | 1.02 ± 0.10 a | 0.39 ± 0.05 b | 1.05 ± 0.21 a | 1.24 ± 0.22 a |
| Il-22 | 1.02 ± 0.24 a | 3.05 ± 0.36 b | 2.81 ± 0.43 b | 2.66 ± 0.59 a,b |
| Ingredients (g/kg of Diet) | P14 |
|---|---|
| Acid casein (Armor Protéines®, ref. 139860) | 112 |
| Whey protein (Armor Protéines®, Protarmor 80, ref. 139805) | 28 |
| Corn starch | 622.4 |
| Sucrose | 100.3 |
| Soybean oil | 40 |
| Alpha cellulose | 50 |
| AIN 93M mineral mix | 35 |
| AIN 93M Vitamins | 10 |
| Choline | 2.3 |
| Metabolizable energy, kJ/g | 14.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blais, A.; Lan, A.; Blachier, F.; Benamouzig, R.; Jouet, P.; Couvineau, A. Efficiency of Orexin-A for Inflammatory Flare and Mucosal Healing in Experimental Colitis: Comparison with the Anti-TNF Alpha Infliximab. Int. J. Mol. Sci. 2023, 24, 9554. https://doi.org/10.3390/ijms24119554
Blais A, Lan A, Blachier F, Benamouzig R, Jouet P, Couvineau A. Efficiency of Orexin-A for Inflammatory Flare and Mucosal Healing in Experimental Colitis: Comparison with the Anti-TNF Alpha Infliximab. International Journal of Molecular Sciences. 2023; 24(11):9554. https://doi.org/10.3390/ijms24119554
Chicago/Turabian StyleBlais, Anne, Annaïg Lan, François Blachier, Robert Benamouzig, Pauline Jouet, and Alain Couvineau. 2023. "Efficiency of Orexin-A for Inflammatory Flare and Mucosal Healing in Experimental Colitis: Comparison with the Anti-TNF Alpha Infliximab" International Journal of Molecular Sciences 24, no. 11: 9554. https://doi.org/10.3390/ijms24119554