
Smooth Muscle-Alpha Actin R149C Pathogenic Variant Downregulates Integrin Recruitment at Cell-Matrix Adhesions and Decreases Cellular Contractility
 , , and
, , and
Abstract
:1. Introduction
2. Results
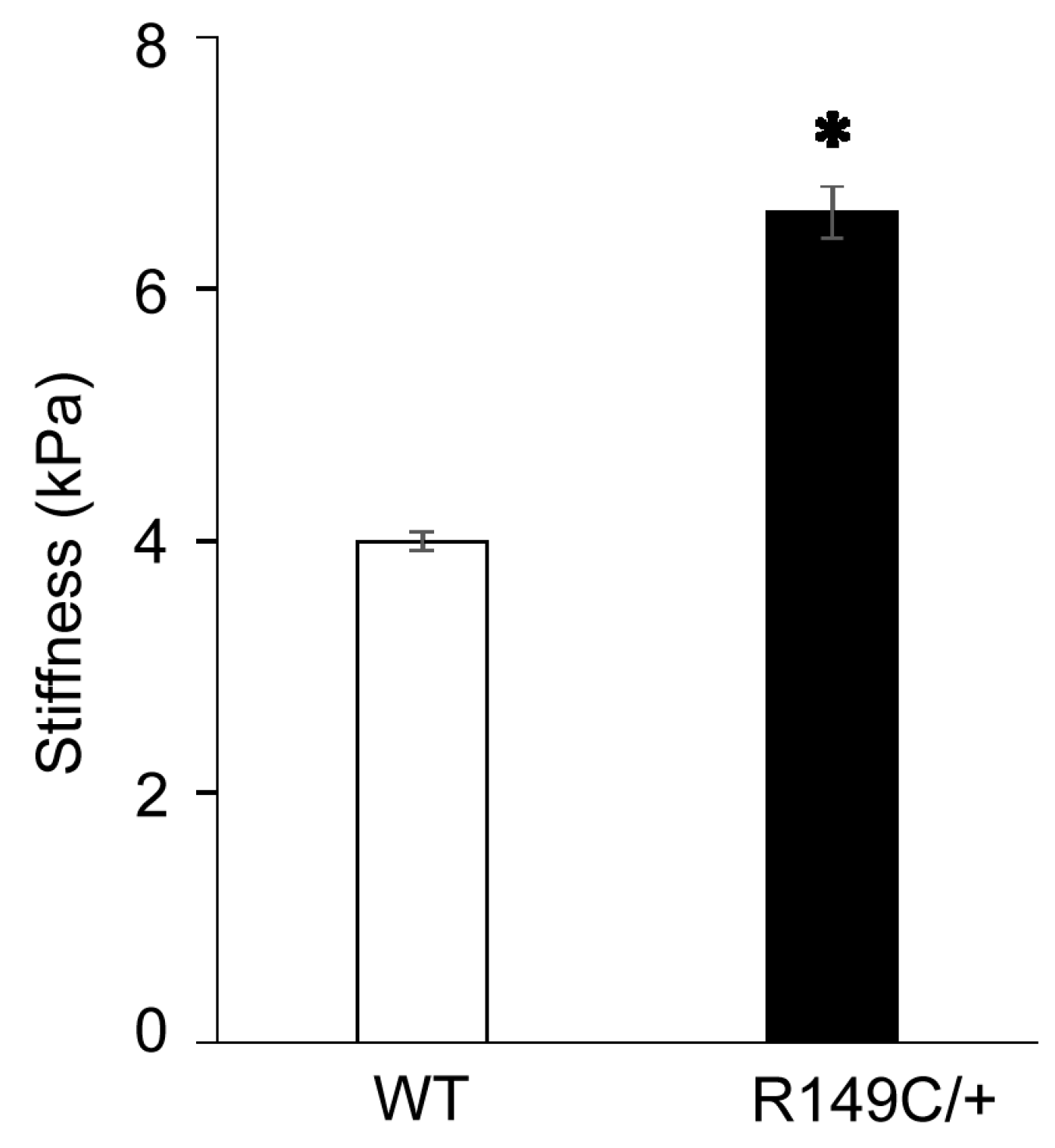
2.1. Functional Impairments in Acta2R149C/+ Aorta
2.2. Smooth Muscle Actin Isoforms Are Differentially Regulated in Acta2R149C/+ Aortic Smooth Muscle Cells
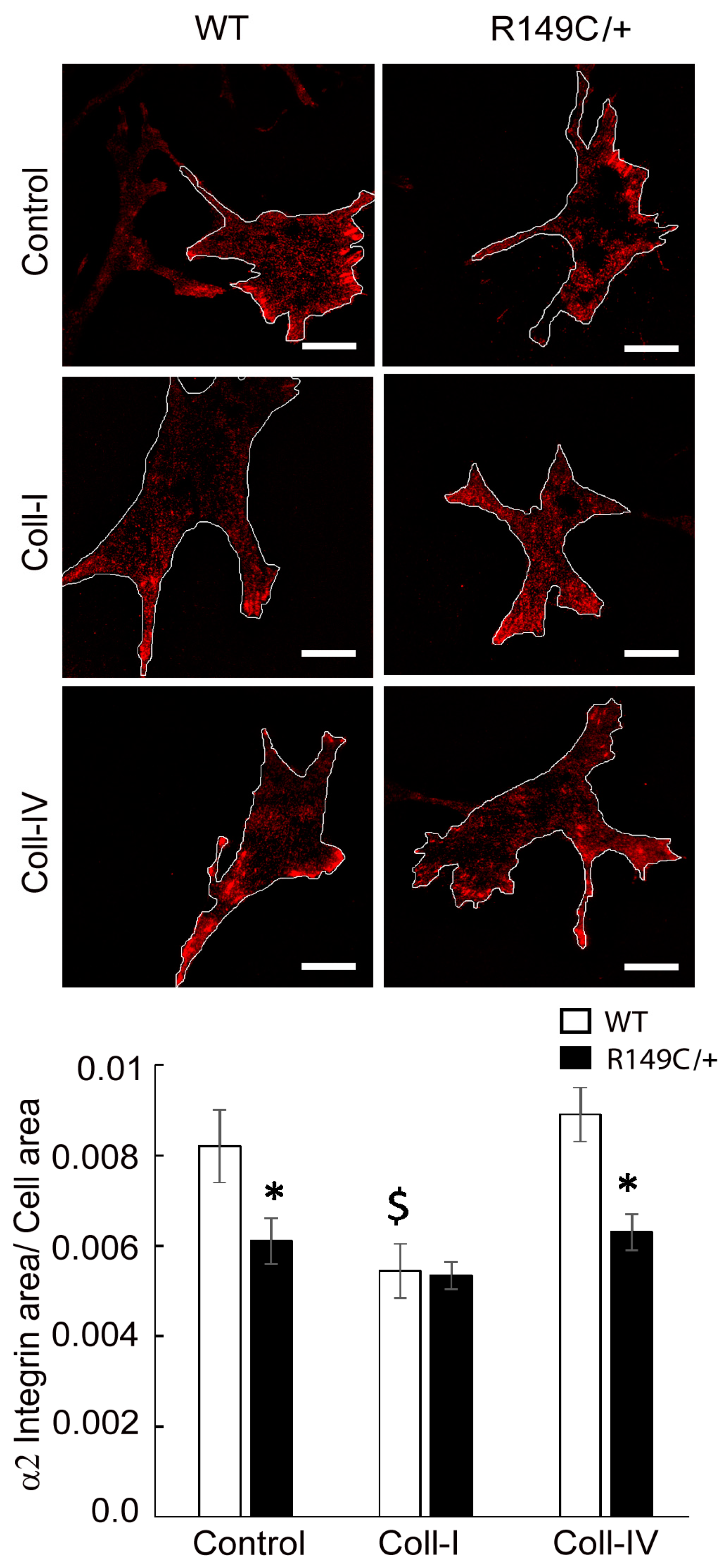
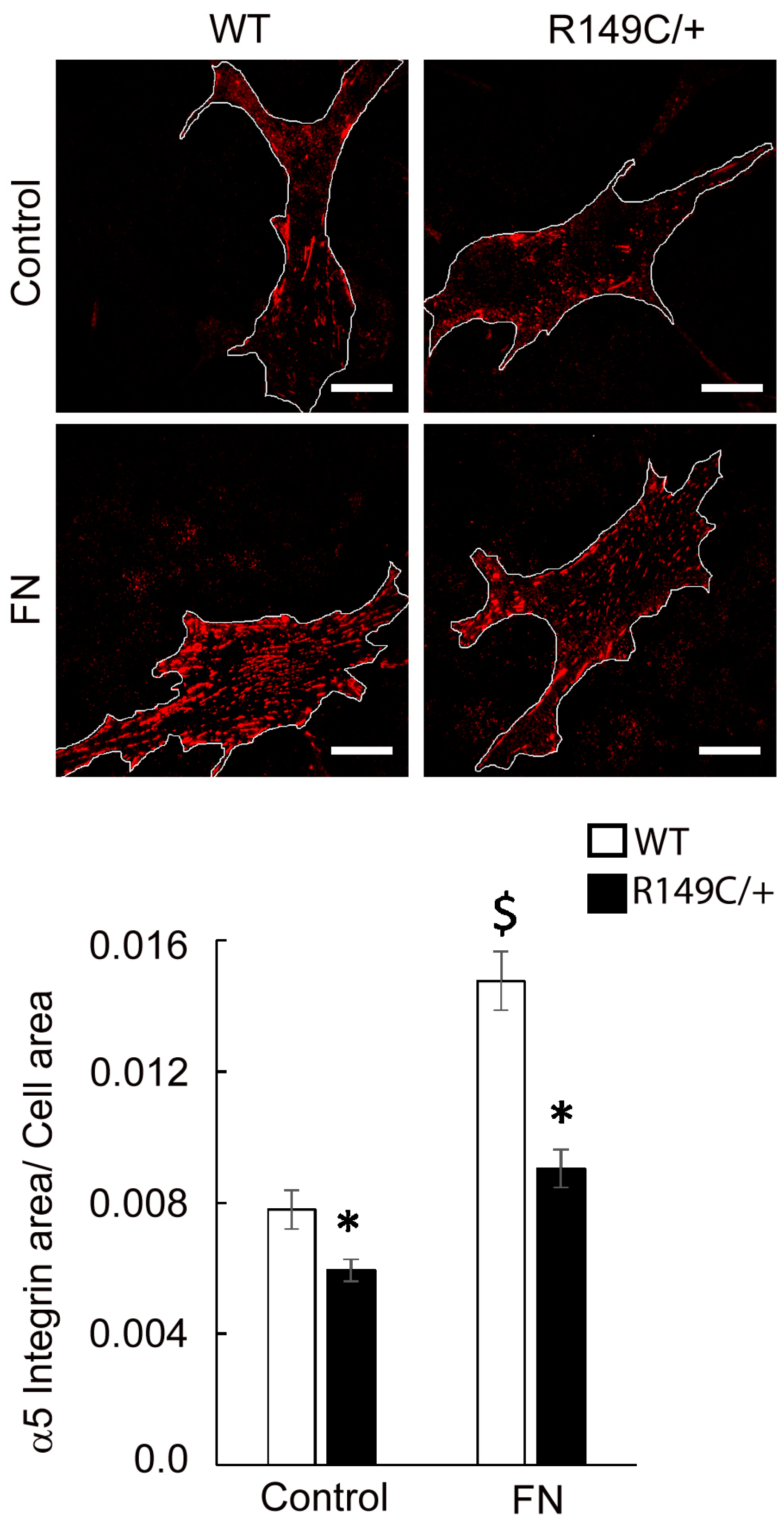
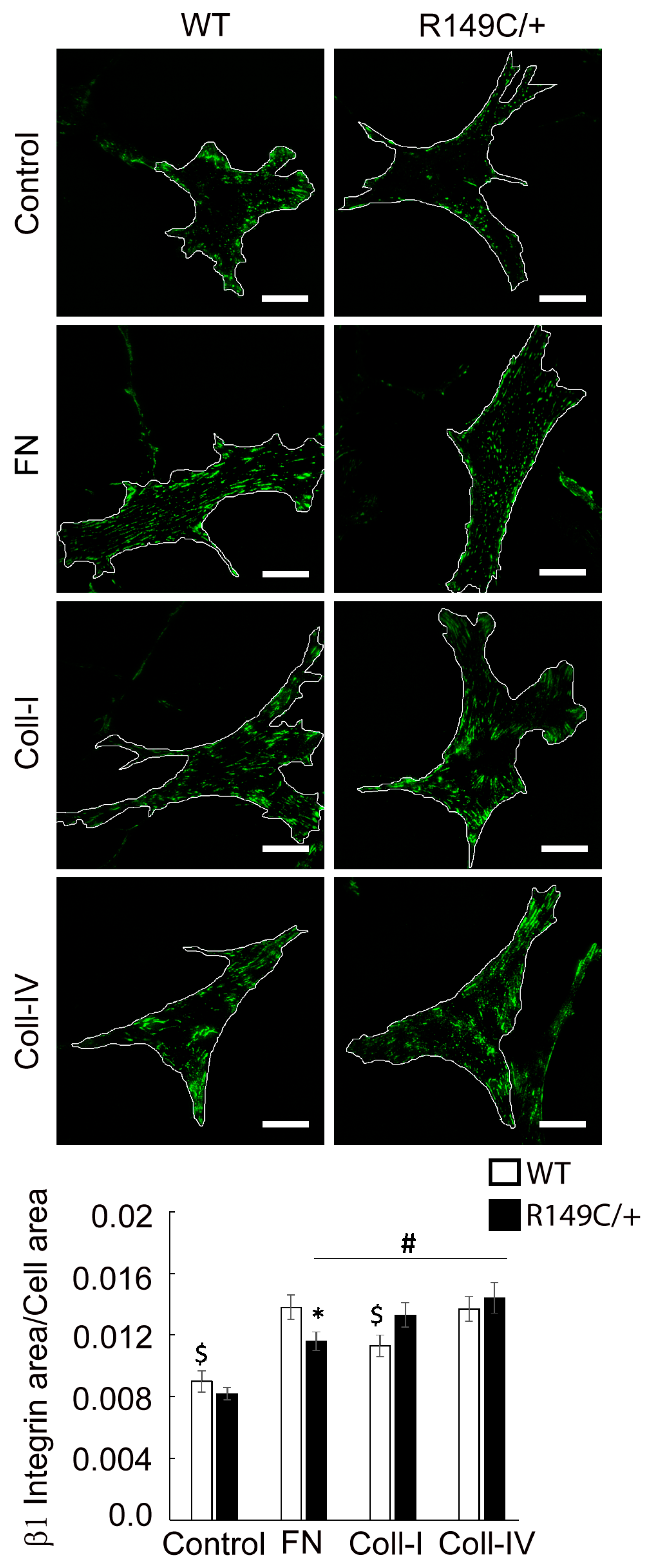
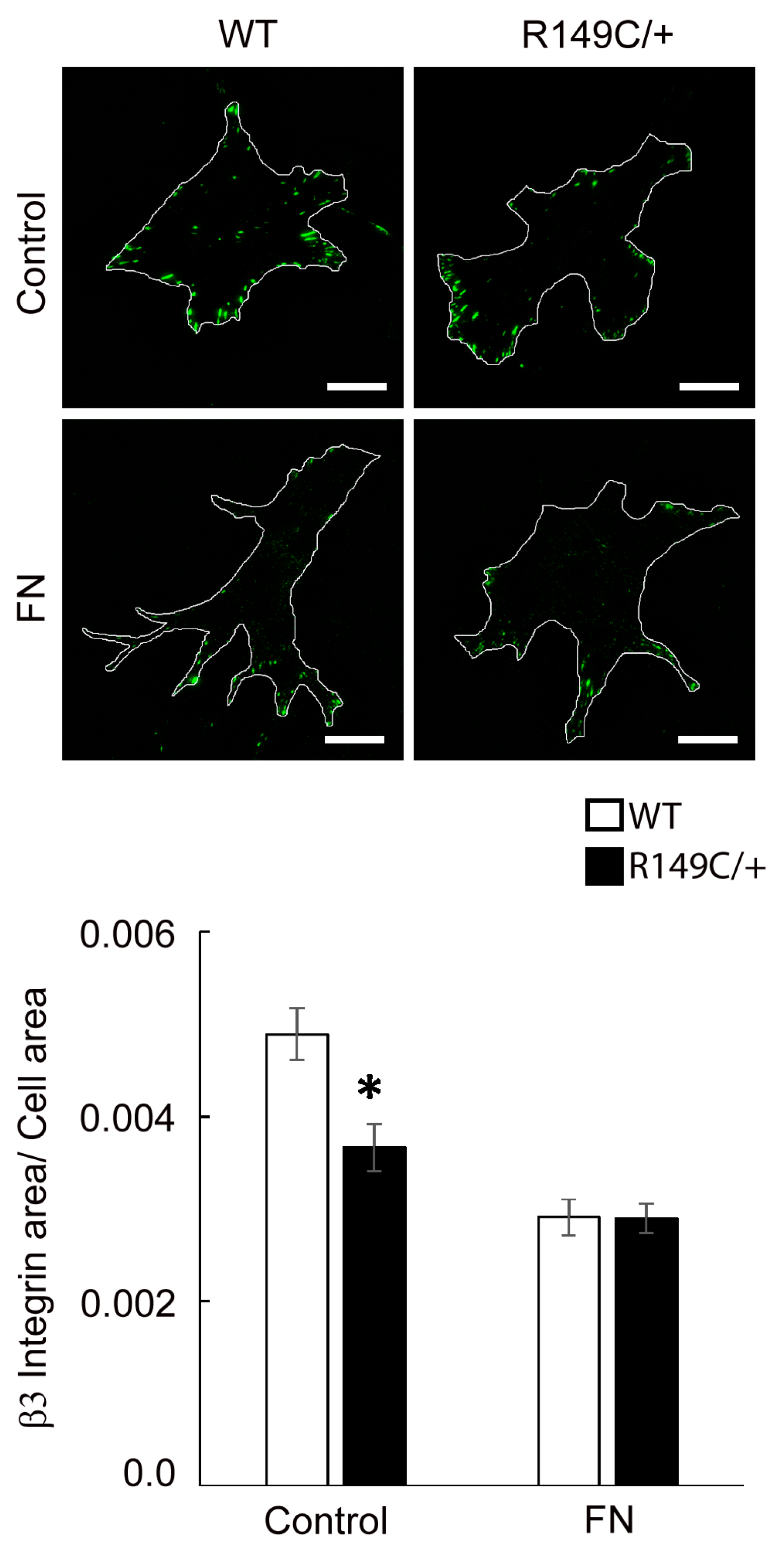
2.3. Adhesion to the Matrix Is Impaired in Acta2R149C/+ Aortic Smooth Muscle Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Aortic Ring Preparation
4.3. Stress Relaxation in Aortic Rings
4.4. Functional Assessment of Contractile Properties
4.5. Smooth Muscle Cell Culture
4.6. Quantitative RT-PCR
4.7. Immunofluorescence Staining
4.8. Smooth Muscle Cell Imaging
4.9. Fluorescence Image Analysis
4.10. Statistical Analysis
4.10.1. Aortic Ring Measurements
4.10.2. Fluorescence Measurements
4.11. Atomic Force Microscopy Measurements
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jansen, K.A.; Donato, D.M.; Balcioglu, H.E.; Schmidt, T.; Danen, E.H.; Koenderink, G.H. A guide to mechanobiology: Where biology and physics meet. Biochim. Biophys. Acta 2015, 1853, 3043–3052. [Google Scholar] [CrossRef]
- Massett, M.P.; Bywaters, B.C.; Gibbs, H.C.; Trzeciakowski, J.P.; Padgham, S.; Chen, J.; Rivera, G.; Yeh, A.T.; Milewicz, D.M.; Trache, A. Loss of smooth muscle α-actin effects on mechanosensing and cell-matrix adhesions. Exp. Biol. Med. (Maywood) 2020, 245, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Pannu, H.; Tran-Fadulu, V.; Milewicz, D.M. Genetic basis of thoracic aortic aneurysms and aortic dissections. Am. J. Med. Genet. Part C Semin. Med. Genet. 2005, 139C, 10–16. [Google Scholar] [CrossRef]
- Regalado, E.S.; Morris, S.A.; Braverman, A.C.; Hostetler, E.M.; De Backer, J.; Li, R.; Pyeritz, R.E.; Yetman, A.T.; Cervi, E.; Shalhub, S.; et al. Comparative Risks of Initial Aortic Events Associated With Genetic Thoracic Aortic Disease. J. Am. Coll. Cardiol. 2022, 80, 857–869. [Google Scholar] [CrossRef]
- Guo, D.C.; Pannu, H.; Tran-Fadulu, V.; Papke, C.L.; Yu, R.K.; Avidan, N.; Bourgeois, S.; Estrera, A.L.; Safi, H.J.; Sparks, E.; et al. Mutations in smooth muscle alpha-actin (ACTA2) lead to thoracic aortic aneurysms and dissections. Nat. Genet. 2007, 39, 1488–1493. [Google Scholar] [CrossRef]
- Chen, J.; Kaw, K.; Lu, H.; Fagnant, P.M.; Chattopadhyay, A.; Duan, X.Y.; Zhou, Z.; Ma, S.; Liu, Z.; Huang, J.; et al. Resistance of Acta2R149C/+ mice to aortic disease is associated with defective release of mutant smooth muscle α-actin from the chaperonin-containing TCP1 folding complex. J. Biol. Chem. 2021, 297, 101228. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pasapera, A.M.; Koretsky, A.P.; Waterman, C.M. Orientation-specific responses to sustained uniaxial stretching in focal adhesion growth and turnover. Proc. Natl. Acad. Sci. USA 2013, 110, E2352–E2361. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lemus, L.A.; Wu, X.; Wilson, E.; Hill, M.A.; Davis, G.E.; Davis, M.J.; Meininger, G.A. Integrins as Unique Receptors for Vascular Control. J. Vasc. Res. 2003, 40, 211–233. [Google Scholar] [CrossRef]
- Martinez-Lemus, L.A.; Crow, T.; Davis, M.J.; Meininger, G.A. alphavbeta3- and alpha5beta1-integrin blockade inhibits myogenic constriction of skeletal muscle resistance arterioles. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H322–H329. [Google Scholar] [CrossRef]
- Milewicz, D.M.; Trybus, K.M.; Guo, D.C.; Sweeney, H.L.; Regalado, E.; Kamm, K.; Stull, J.T. Altered Smooth Muscle Cell Force Generation as a Driver of Thoracic Aortic Aneurysms and Dissections. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 26–34. [Google Scholar] [CrossRef]
- Schildmeyer, L.A.; Braun, R.; Taffet, G.; Debiasi, M.; Burns, A.E.; Bradley, A.; Schwartz, R.J. Impaired vascular contractility and blood pressure homeostasis in the smooth muscle alpha-actin null mouse. FASEB J. 2000, 14, 2213–2220. [Google Scholar] [CrossRef]
- Papke, C.L.; Cao, J.; Kwartler, C.S.; Villamizar, C.; Byanova, K.L.; Lim, S.M.; Sreenivasappa, H.; Fischer, G.; Pham, J.; Rees, M.; et al. Smooth muscle hyperplasia due to loss of smooth muscle alpha-actin is driven by activation of focal adhesion kinase, altered p53 localization and increased levels of platelet-derived growth factor receptor-beta. Hum. Mol. Genet. 2013, 22, 3123–3137. [Google Scholar] [CrossRef]
- Cluzel, C.; Saltel, F.; Lussi, J.; Paulhe, F.; Imhof, B.A.; Wehrle-Haller, B. The mechanisms and dynamics of (alpha)v(beta)3 integrin clustering in living cells. J. Cell Biol. 2005, 171, 383–392. [Google Scholar] [CrossRef]
- Regalado, E.S.; Guo, D.C.; Prakash, S.; Bensend, T.A.; Flynn, K.; Estrera, A.; Safi, H.; Liang, D.; Hyland, J.; Child, A.; et al. Aortic Disease Presentation and Outcome Associated With ACTA2 Mutations. Circ. Cardiovasc. Genet. 2015, 8, 457–464. [Google Scholar] [CrossRef]
- Guo, D.C.; Papke, C.L.; Tran-Fadulu, V.; Regalado, E.S.; Avidan, N.; Johnson, R.J.; Kim, D.H.; Pannu, H.; Willing, M.C.; Sparks, E.; et al. Mutations in smooth muscle alpha-actin (ACTA2) cause coronary artery disease, stroke, and Moyamoya disease, along with thoracic aortic disease. Am. J. Hum. Genet. 2009, 84, 617–627. [Google Scholar] [CrossRef]
- Gao, Y.Z.; Saphirstein, R.J.; Yamin, R.; Suki, B.; Morgan, K.G. Aging impairs smooth muscle-mediated regulation of aortic stiffness: A defect in shock absorption function? Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1252–H1261. [Google Scholar] [CrossRef] [PubMed]
- Leloup, A.J.A.; Van Hove, C.E.; De Moudt, S.; De Meyer, G.R.Y.; De Keulenaer, G.W.; Fransen, P. Vascular smooth muscle cell contraction and relaxation in the isolated aorta: A critical regulator of large artery compliance. Physiol. Rep. 2019, 7, e13934. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, J.B.; Mukherjee, R.; Stroud, R.E.; Jones, J.A.; Ikonomidis, J.S. Relation of murine thoracic aortic structural and cellular changes with aging to passive and active mechanical properties. J. Am. Heart Assoc. 2015, 4, e001744. [Google Scholar] [CrossRef]
- Chen, J.; Peters, A.; Papke, C.L.; Villamizar, C.; Ringuette, L.J.; Cao, J.; Wang, S.; Ma, S.; Gong, L.; Byanova, K.L.; et al. Loss of Smooth Muscle alpha-Actin Leads to NF-kappaB-Dependent Increased Sensitivity to Angiotensin II in Smooth Muscle Cells and Aortic Enlargement. Circ. Res. 2017, 120, 1903–1915. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Davis, G.E.; Meininger, G.A.; Wilson, E.; Davis, M.J. Regulation of the L-type calcium channel by alpha 5beta 1 integrin requires signaling between focal adhesion proteins. J. Biol. Chem. 2001, 276, 30285–30292. [Google Scholar] [CrossRef]
- Wu, X.; Mogford, J.E.; Platts, S.H.; Davis, G.E.; Meininger, G.A.; Davis, M.J. Modulation of calcium current in arteriolar smooth muscle by alphav beta3 and alpha5 beta1 integrin ligands. J. Cell Biol. 1998, 143, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Bellini, C.; Wang, S.; Milewicz, D.M.; Humphrey, J.D. Myh11(R247C/R247C) mutations increase thoracic aorta vulnerability to intramural damage despite a general biomechanical adaptivity. J. Biomech. 2015, 48, 113–121. [Google Scholar] [CrossRef]
- Cao, J.; Gong, L.; Guo, D.C.; Mietzsch, U.; Kuang, S.Q.; Kwartler, C.S.; Safi, H.; Estrera, A.; Gambello, M.J.; Milewicz, D.M. Thoracic aortic disease in tuberous sclerosis complex: Molecular pathogenesis and potential therapies in Tsc2+/− mice. Hum. Mol. Genet. 2010, 19, 1908–1920. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Martinez-Lemus, L.A.; Trache, A.; Trzeciakowski, J.P.; Davis, G.E.; Pohl, U.; Meininger, G.A. Mechanical properties of the interaction between fibronectin and alpha5beta1-integrin on vascular smooth muscle cells studied using atomic force microscopy. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2526–H2535. [Google Scholar] [CrossRef]
- Seawright, J.W.; Sreenivasappa, H.; Gibbs, H.C.; Padgham, S.; Shin, S.Y.; Chaponnier, C.; Yeh, A.T.; Trzeciakowski, J.P.; Woodman, C.R.; Trache, A. Vascular Smooth Muscle Contractile Function Declines With Age in Skeletal Muscle Feed Arteries. Front. Physiol. 2018, 9, 856. [Google Scholar] [CrossRef] [PubMed]
- Arnoldi, R.; Hiltbrunner, A.; Dugina, V.; Tille, J.C.; Chaponnier, C. Smooth muscle actin isoforms: A tug of war between contraction and compliance. Eur. J. Cell Biol. 2013, 92, 187–200. [Google Scholar] [CrossRef]
- Chaponnier, C.; Gabbiani, G. Monoclonal antibodies against muscle actin isoforms: Epitope identification and analysis of isoform expression by immunoblot and immunostaining in normal and regenerating skeletal muscle. F1000Res 2016, 5, 416. [Google Scholar] [CrossRef]
- Trache, A.; Meininger, G.A. Total internal reflection fluorescence (TIRF) microscopy. Curr. Protoc. Microbiol. 2008, 10, 2A.2.1–2A.2.22. [Google Scholar] [CrossRef]
- Trache, A.; Lim, S.M. Integrated microscopy for real-time imaging of mechanotransduction studies in live cells. J. Biomed. Opt. 2009, 14, 034024. [Google Scholar] [CrossRef]
- Lim, S.M.; Kreipe, B.A.; Trzeciakowski, J.; Dangott, L.; Trache, A. Extracellular matrix effect on RhoA signaling modulation in vascular smooth muscle cells. Exp. Cell Res. 2010, 316, 2833–2848. [Google Scholar] [CrossRef]
- Ballestrem, C.; Hinz, B.; Imhof, B.A.; Wehrle-Haller, B. Marching at the front and dragging behind: Differential alphaVbeta3-integrin turnover regulates focal adhesion behavior. J. Cell Biol. 2001, 155, 1319–1332. [Google Scholar] [CrossRef] [PubMed]
- Trache, A.; Meininger, G.A. Atomic force microscopy (AFM). Curr. Protoc. Microbiol. 2008, 8, 2C.2.1–2C.2.17. [Google Scholar] [CrossRef] [PubMed]
- Trzeciakowski, J.; Meininger, G. NForceR: Nanoscale Force Reader and AFM Data Analysis Package (Copyrighted), 2004.
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-Plus; Springer: Berlin/Heidelberg, Germany, 1994. [Google Scholar]
- Trache, A.; Trzeciakowski, J.P.; Gardiner, L.; Sun, Z.; Muthuchamy, M.; Guo, M.; Yuan, S.Y.; Meininger, G.A. Histamine effects on endothelial cell fibronectin interaction studied by atomic force microscopy. Biophys. J. 2005, 89, 2888–2898. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ -> 3′) | Reverse (5′ -> 3′) |
|---|---|---|
| Acta2 | GTCCCAGACATCAGGGAGTAA | TCGGATACTTCAGCGTCAGGA |
| Actg2 | CCGCCCTAGACATCAGGGT | TCTTCTGGTGCTACTCGAAGC |
| Itga2 | TACAGACGTGCTCCTGGTAGGT | CCGAGCATTTCCAGTGCCTTCT |
| Itga5 | GTGTGAGGAACTGGTCGCCTAT | CCGTTCTCTGGTCCAACCGATA |
| Itgb1 | CTCCAGAAGGTGGCTTTGATGC | GTGAAACCCAGCATCCGTGGAA |
| Itgb3 | GTGAGTGCGATGACTTCTCCTG | CAGGTGTCAGTGCGTGTAGTAC |
| 18S | GTAACCCGTTGAACCCCATT | CCATCCAATCGGTAGTAGCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojha, K.R.; Kim, H.; Padgham, S.; Hopkins, L.; Zamen, R.J.; Chattopadhyay, A.; Han, G.; Milewicz, D.M.; Massett, M.P.; Trache, A. Smooth Muscle-Alpha Actin R149C Pathogenic Variant Downregulates Integrin Recruitment at Cell-Matrix Adhesions and Decreases Cellular Contractility. Int. J. Mol. Sci. 2023, 24, 9616. https://doi.org/10.3390/ijms24119616
Ojha KR, Kim H, Padgham S, Hopkins L, Zamen RJ, Chattopadhyay A, Han G, Milewicz DM, Massett MP, Trache A. Smooth Muscle-Alpha Actin R149C Pathogenic Variant Downregulates Integrin Recruitment at Cell-Matrix Adhesions and Decreases Cellular Contractility. International Journal of Molecular Sciences. 2023; 24(11):9616. https://doi.org/10.3390/ijms24119616
Chicago/Turabian StyleOjha, Krishna R., Hyoseon Kim, Samuel Padgham, Laura Hopkins, Robert J. Zamen, Abhijnan Chattopadhyay, Gang Han, Dianna M. Milewicz, Michael P. Massett, and Andreea Trache. 2023. "Smooth Muscle-Alpha Actin R149C Pathogenic Variant Downregulates Integrin Recruitment at Cell-Matrix Adhesions and Decreases Cellular Contractility" International Journal of Molecular Sciences 24, no. 11: 9616. https://doi.org/10.3390/ijms24119616