
The Effect of Anti-Autotaxin Aptamers on the Development of Proliferative Vitreoretinopathy
 , , and
, , and 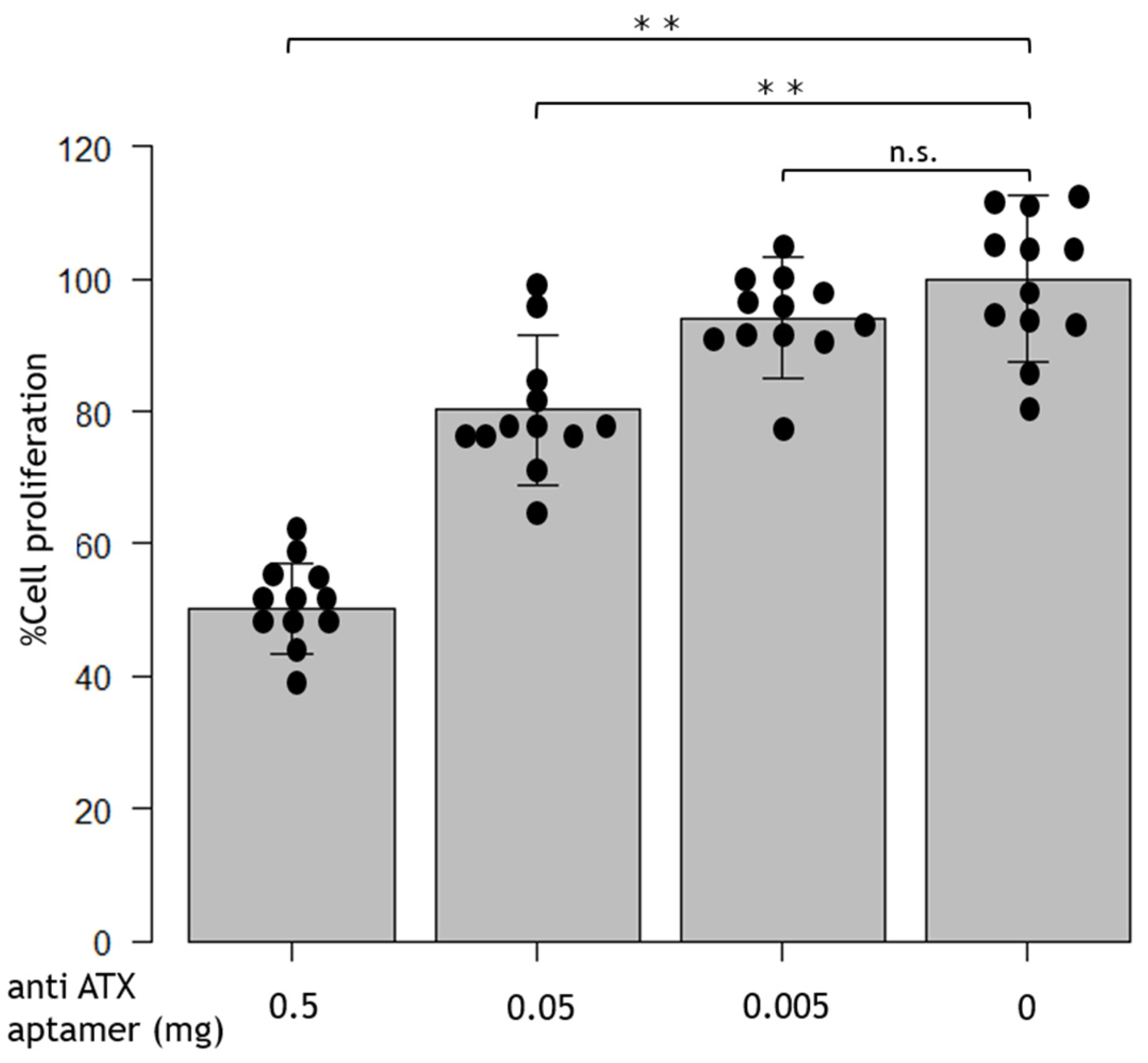{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. RBM-006 Profile
2.2. Anti-ATX Aptamer Inhibits RPE Proliferation
2.3. Anti-ATX Aptamer Inhibits RPE Migration
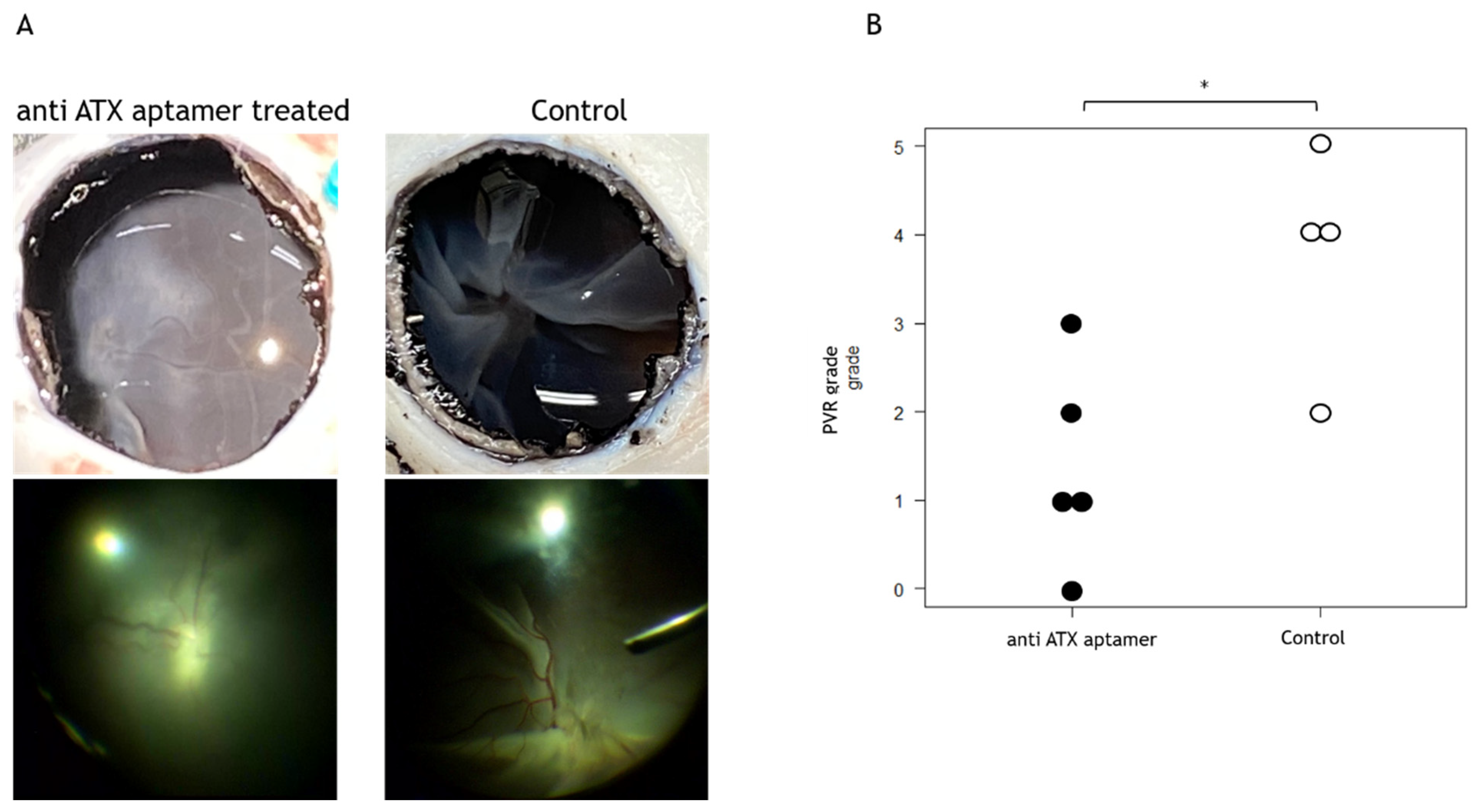
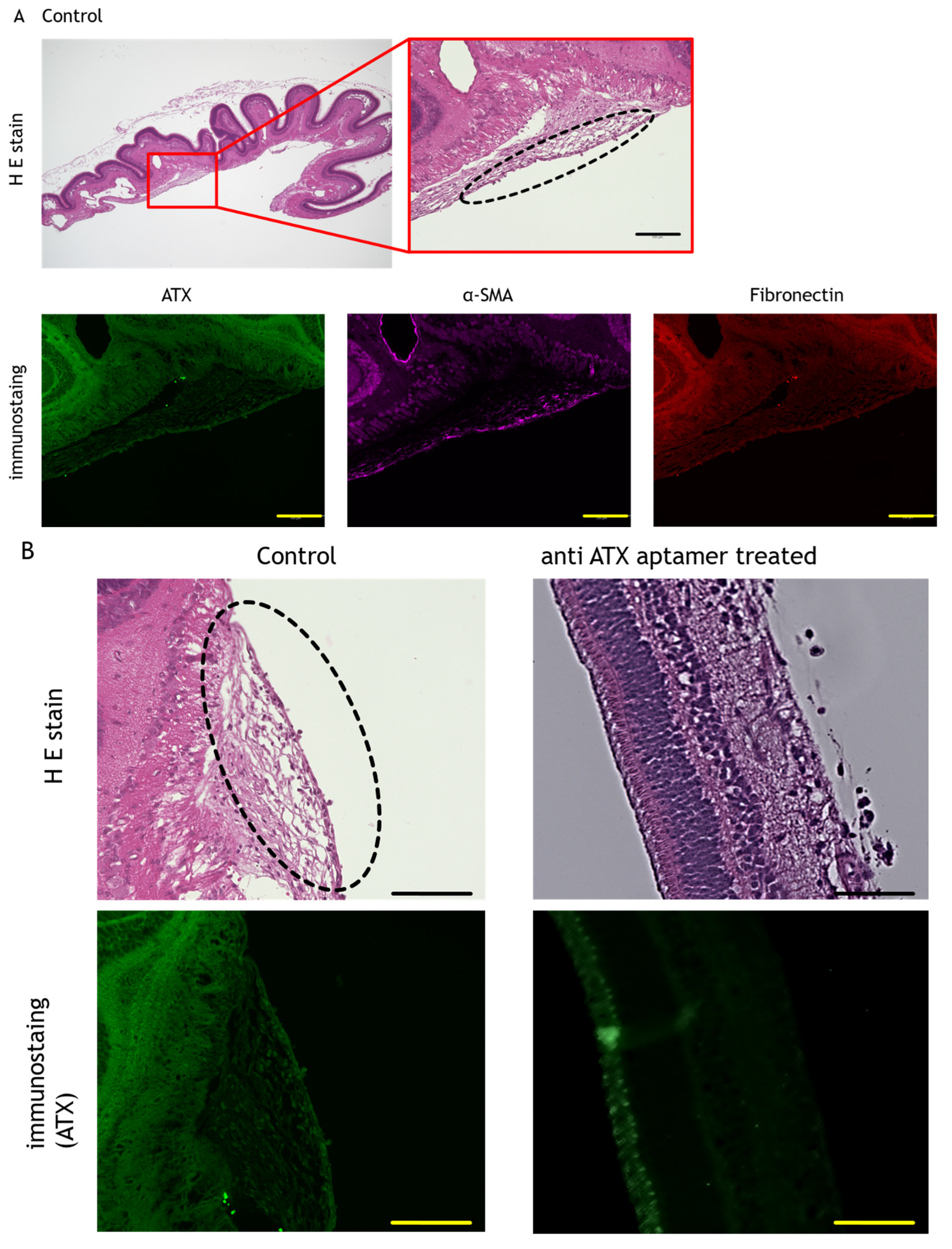
2.4. Anti-ATX Aptamer Inhibits the Progression of PVR In Vivo
3. Discussion
4. Materials and Methods
4.1. RBM-006 Profile
4.2. In Vitro RPE Cell Culture
4.3. In Vivo Swine PVR Model
4.4. Immunofluorescence and Immunohistochemistry
4.5. Cell Proliferation Assay
4.6. Cell Migration Assay
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rouberol, F.; Chiquet, C. Proliferative Vitreoretinopathy: Pathophysiology and Clinical Diagnosis. J. Fr. Ophtalmol. 2014, 37, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Pastor, J.C.; de la Rúa, E.R.; Martín, F. Proliferative Vitreoretinopathy: Risk Factors and Pathobiology. Prog. Retin. Eye Res. 2002, 21, 127–144. [Google Scholar] [CrossRef]
- Pennock, S.; Haddock, L.J.; Mukai, S.; Kazlauskas, A. Vascular Endothelial Growth Factor Acts Primarily via Platelet-Derived Growth Factor Receptor α to Promote Proliferative Vitreoretinopathy. Am. J. Pathol. 2014, 184, 3052–3068. [Google Scholar] [CrossRef] [PubMed]
- Charteris, D.G.; Sethi, C.S.; Lewis, G.P.; Fisher, S.K. Proliferative Vitreoretinopathy-Developments in Adjunctive Treatment and Retinal Pathology. Eye 2002, 16, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Sadaka, A.; Giuliari, G.P. Proliferative Vitreoretinopathy: Current and Emerging Treatments. Clin. Ophthalmol. 2012, 6, 1325–1333. [Google Scholar] [PubMed]
- Narala, R.; Nassiri, N.; Kim, C.; Mehregan, C.; Padidam, S.; Abrams, G.W. Outcomes of Repeat Pars Plana Vitrectomy After Failed Surgery for Proliferative Vitreoretinopathy. Retina 2018, 38 (Suppl. S1), S49–S59. [Google Scholar] [CrossRef] [PubMed]
- Hiscott, P.; Sheridan, C.; Magee, R.M.; Grierson, I. Matrix and the Retinal Pigment Epithelium in Proliferative Retinal Disease. Prog. Retin. Eye Res. 1999, 18, 167–190. [Google Scholar] [CrossRef]
- Nagasaki, H.; Shinagawa, K.; Mochizuki, M. Risk Factors for Proliferative Vitreoretinopathy. Prog. Retin. Eye Res. 1998, 17, 77–98. [Google Scholar] [CrossRef]
- Li, H.; Wang, H.; Wang, F.; Gu, Q.; Xu, X. Snail Involves in the Transforming Growth Factor β1-Mediated Epithelial-Mesenchymal Transition of Retinal Pigment Epithelial Cells. PLoS ONE 2011, 6, e23322. [Google Scholar] [CrossRef]
- Tamiya, S.; Liu, L.; Kaplan, H.J. Epithelial-Mesenchymal Transition and Proliferation of Retinal Pigment Epithelial Cells Initiated upon Loss of Cell-Cell Contact. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2755–2763. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Casaroli-Marano, R.P.; Pagan, R.; Vilaró, S. Epithelial-Mesenchymal Transition in Proliferative Vitreoretinopathy: Intermediate Filament Protein Expression in Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2062–2072. [Google Scholar]
- Chen, Z.; Shao, Y.; Li, X. The Roles of Signaling Pathways in Epithelial-to-Mesenchymal Transition of PVR. Mol. Vis. 2015, 21, 706–710. [Google Scholar] [PubMed]
- Bächner, D.; Ahrens, M.; Betat, N.; Schröder, D.; Gross, G. Developmental Expression Analysis of Murine Autotaxin (ATX). Mech. Dev. 1999, 84, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Murata, J.; Clair, T.; Polymeropoulos, M.H.; Torres, R.; Manrow, R.E.; Liotta, L.A.; Stracke, M.L. Cloning, Chromosomal Localization, and Tissue Expression of Autotaxin from Human Teratocarcinoma Cells. Biochem. Biophys. Res. Commun. 1996, 218, 714–719. [Google Scholar] [CrossRef]
- Savaskan, N.E.; Rocha, L.; Kotter, M.R.; Baer, A.; Lubec, G.; van Meeteren, L.A.; Kishi, Y.; Aoki, J.; Moolenaar, W.H.; Nitsch, R.; et al. Autotaxin (NPP-2) in the Brain: Cell Type-Specific Expression and Regulation during Development and after Neurotrauma. Cell. Mol. Life Sci. 2007, 64, 230–243. [Google Scholar] [CrossRef]
- Honjo, M.; Igarashi, N.; Kurano, M.; Yatomi, Y.; Igarashi, K.; Kano, K.; Aoki, J.; Weinreb, R.N.; Aihara, M. Autotaxin-Lysophosphatidic Acid Pathway in Intraocular Pressure Regulation and Glaucoma Subtypes. Investig. Ophthalmol. Vis. Sci. 2018, 59, 693–701. [Google Scholar] [CrossRef]
- Honjo, M.; Igarashi, N.; Nishida, J.; Kurano, M.; Yatomi, Y.; Igarashi, K.; Kano, K.; Aoki, J.; Aihara, M. Role of the Autotaxin-LPA Pathway in Dexamethasone-Induced Fibrotic Responses and Extracellular Matrix Production in Human Trabecular Meshwork Cells. Investig. Ophthalmol. Vis. Sci. 2018, 59, 21–30. [Google Scholar] [CrossRef]
- Kato, K.; Ikeda, H.; Miyakawa, S.; Futakawa, S.; Nonaka, Y.; Fujiwara, M.; Okudaira, S.; Kano, K.; Aoki, J.; Morita, J.; et al. Structural Basis for Specific Inhibition of Autotaxin by a DNA Aptamer. Nat. Struct. Mol. Biol. 2016, 23, 395–401. [Google Scholar] [CrossRef]
- Zhu, G.; Chen, X. Aptamer-Based Targeted Therapy. Adv. Drug Deliv. Rev. 2018, 134, 65–78. [Google Scholar] [CrossRef]
- Nakamura, Y. Aptamers as Therapeutic Middle Molecules. Biochimie 2018, 145, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Rossi, J. Aptamers as Targeted Therapeutics: Current Potential and Challenges. Nat. Rev. Drug Discov. 2017, 16, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Ma, J.X.; Shang, Q.L.; An, J.B.; Chen, H.T. Crocetin Inhibits the Proliferation, Migration and TGF-β2-Induced Epithelial-Mesenchymal Transition of Retinal Pigment Epithelial Cells. Eur. J. Pharmacol. 2017, 815, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, H.; Liu, X.; Xu, D.; Wang, F. MicroRNA-29b Regulates TGF-β1-Mediated Epithelial-Mesenchymal Transition of Retinal Pigment Epithelial Cells by Targeting AKT2. Exp. Cell Res. 2016, 345, 115–124. [Google Scholar] [CrossRef]
- Matsuda, Y.; Nonaka, Y.; Futakawa, S.; Imai, H.; Akita, K.; Nishihata, T.; Fujiwara, M.; Ali, Y.; Bhisitkul, R.B.; Nakamura, Y. Anti-angiogenic and Anti-scarring Dual Action of an Anti-fibroblast Growth Factor 2 Aptamer in Animal Models of Retinal Disease. Mol. Ther. Nucleic Acids 2019, 17, 819–828. [Google Scholar] [CrossRef]
- Liang, C.M.; Tai, M.C.; Chang, Y.H.; Chen, Y.H.; Chen, C.L.; Lu, D.W.; Chen, J.T. Glucosamine Inhibits Epithelial-to-Mesenchymal Transition and Migration of Retinal Pigment Epithelium Cells in Culture and Morphologic Changes in a Mouse Model of Proliferative Vitreoretinopathy. Acta Ophthalmol. 2011, 89, e505–e514. [Google Scholar] [CrossRef]
- Ishikawa, K.; He, S.; Terasaki, H.; Nazari, H.; Zhang, H.; Spee, C.; Kannan, R.; Hinton, D.R. Resveratrol Inhibits Epithelial-Mesenchymal Transition of Retinal Pigment Epithelium and Development of Proliferative Vitreoretinopathy. Sci. Rep. 2015, 5, 16386. [Google Scholar] [CrossRef]
- Zheng, Y.; Bando, H.; Ikuno, Y.; Oshima, Y.; Sawa, M.; Ohji, M.; Tano, Y. Involvement of Rho-Kinase Pathway in Contractile Activity of Rabbit RPE Cells In Vivo and In Vitro. Investig. Ophthalmol. Vis. Sci. 2004, 45, 668–674. [Google Scholar] [CrossRef]
- Kita, T.; Hata, Y.; Arita, R.; Kawahara, S.; Miura, M.; Nakao, S.; Mochizuki, Y.; Enaida, H.; Goto, Y.; Shimokawa, H.; et al. Role of TGF-Beta in Proliferative Vitreoretinal Diseases and ROCK as a Therapeutic Target. Proc. Natl. Acad. Sci. USA 2008, 105, 17504–17509. [Google Scholar] [CrossRef]
- Xu, M.Y.; Porte, J.; Knox, A.J.; Weinreb, P.H.; Maher, T.M.; Violette, S.M.; McAnulty, R.J.; Sheppard, D.; Jenkins, G. Lysophosphatidic Acid Induces alphavbeta6 Integrin-Mediated TGF-Beta Activation via the LPA2 Receptor and the Small G Protein G Alpha(q). Am. J. Pathol. 2009, 174, 1264–1279. [Google Scholar] [CrossRef]
- Geng, H.; Lan, R.; Singha, P.K.; Gilchrist, A.; Weinreb, P.H.; Violette, S.M.; Weinberg, J.M.; Saikumar, P.; Venkatachalam, M.A. Lysophosphatidic Acid Increases Proximal Tubule Cell Secretion of Profibrotic Cytokines PDGF-B and CTGF Through LPA2- and Gαq-Mediated Rho and αvβ6 Integrin-Dependent Activation of TGF-β. Am. J. Pathol. 2012, 181, 1236–1249. [Google Scholar] [CrossRef] [PubMed]
- Zuehlke, J.; Ebenau, A.; Krueger, B.; Goppelt-Struebe, M. Vectorial Secretion of CTGF as a Cell-Type Specific Response to LPA and TGF-β in Human Tubular Epithelial Cells. Cell Commun. Signal. 2012, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Chun, J.; Duffield, J.S.; Wada, T.; Luster, A.D.; Tager, A.M. LPA1-Induced Cytoskeleton Reorganization Drives Fibrosis Through CTGF-Dependent Fibroblast Proliferation. FASEB J. 2013, 27, 1830–1846. [Google Scholar] [CrossRef] [PubMed]
- Tager, A.M.; LaCamera, P.; Shea, B.S.; Campanella, G.S.; Selman, M.; Zhao, Z.; Polosukhin, V.; Wain, J.; Karimi-Shah, B.A.; Kim, N.D.; et al. The Lysophosphatidic Acid Receptor LPA1 Links Pulmonary Fibrosis to Lung Injury by Mediating Fibroblast Recruitment and Vascular Leak. Nat. Med. 2008, 14, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Moll, S.; Chaykovska, L.; Meier, M.; Budd, D.C.; Formentini, I.; Pomposiello, S.; Prunotto, M. Targeting the Epithelial Cells in Fibrosis: A New Concept for an Old Disease. Drug Discov. Today 2013, 18, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Thoreson, W.B.; Khandalavala, B.N.; Manahan, R.G.; Polyak, I.A.; Liu, J.L.; Chacko, D.M. Lysophosphatidic Acid Stimulates Proliferation of Human Retinal Pigment Epithelial Cells. Curr. Eye Res. 1997, 16, 698–702. [Google Scholar] [CrossRef]
- Konno, T.; Kotani, T.; Setiawan, J.; Nishigaito, Y.; Sawada, N.; Imada, S.; Saito, Y.; Murata, Y.; Matozaki, T. Role of Lysophosphatidic Acid in Proliferation and Differentiation of Intestinal Epithelial Cells. PLoS ONE 2019, 14, e0215255. [Google Scholar] [CrossRef]
- Umazume, K.; Liu, L.; Scott, P.A.; de Castro, J.P.; McDonald, K.; Kaplan, H.J.; Tamiya, S. Inhibition of PVR with a Tyrosine Kinase Inhibitor, Dasatinib, in the Swine. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1150–1159. [Google Scholar] [CrossRef]
- Abu El-Asrar, A.M.; Missotten, L.; Geboes, K. Expression of Autotaxin and Acylglycerol Kinase in Proliferative Vitreoretinal Epiretinal Membranes. Acta Ophthalmol. 2012, 90, e84–e89. [Google Scholar] [CrossRef]
- Tamiya, S.; Kaplan, H.J. Role of Epithelial-Mesenchymal Transition in Proliferative Vitreoretinopathy. Exp. Eye Res. 2016, 142, 26–31. [Google Scholar] [CrossRef]
- Tosi, G.M.; Marigliani, D.; Romeo, N.; Toti, P. Disease Pathways in Proliferative Vitreoretinopathy: An Ongoing Challenge. J. Cell. Physiol. 2014, 229, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, I.; Martin, R.; Ussa, F.; Fernandez-Bueno, I. The Parameters of the Porcine Eyeball. Graefes Arch. Clin. Exp. Ophthalmol. 2011, 249, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Bertschinger, D.R.; Beknazar, E.; Simonutti, M.; Safran, A.B.; Sahel, J.A.; Rosolen, S.G.; Picaud, S.; Salzmann, J. A Review of In Vivo Animal Studies in Retinal Prosthesis Research. Graefes Arch. Clin. Exp. Ophthalmol. 2008, 246, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Spratt, J.R.; Mattison, L.M.; Iaizzo, P.A.; Meyer, C.; Brown, R.Z.; Iles, T.; Panoskaltsis-Mortari, A.; Loor, G. Lung transplant after prolonged ex vivo lung perfusion: Predictors of allograft function in swine. Transpl. Int. 2018, 31, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, P.G.; Bittle, G.J.; Williams, K.; Pasrija, C.; Xu, K.; Wei, X.; Wu, Z.J.; Griffith, B.P. Ex vivo lung evaluation of prearrest heparinization in donation after cardiac death. Ann. Surg. 2013, 257, 534–541. [Google Scholar] [CrossRef]
- Park, H.S.; Kim, J.E.; You, H.J.; Gu, J.; Yoo, B.; Lee, S.; Lee, H.J.; Hwang, H.Y.; Hwang, Y.; Kim, H.K.; et al. Beneficial effect of a nitric oxide donor in an ex vivo model of pig-to-human pulmonary xenotransplantation. Xenotransplantation 2015, 22, 391–398. [Google Scholar] [CrossRef]
- Ikeda, H.; Miyakawa, S. Aptamer Inhibiting Biological Activity of Autotaxin by Binding with Autotaxin, and Use Thereof. Patent Application No. WO/2015/147290. 2015. Available online: https://patentscope2.wipo.int/search/en/detail.jsf?docId=WO2015147290 (accessed on 10 July 2021).
- Sonoda, S.; Spee, C.; Barron, E.; Ryan, S.J.; Kannan, R.; Hinton, D.R. A Protocol for the Culture and Differentiation of Highly Polarized Human Retinal Pigment Epithelial Cells. Nat. Protoc. 2009, 4, 662–673. [Google Scholar] [CrossRef]
- Umazume, K.; Barak, Y.; McDonald, K.; Liu, L.; Kaplan, H.J.; Tamiya, S. Proliferative Vitreoretinopathy in the Swine-A New Model. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4910–4916. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanazaki, H.; Yokota, H.; Yamagami, S.; Nakamura, Y.; Nagaoka, T. The Effect of Anti-Autotaxin Aptamers on the Development of Proliferative Vitreoretinopathy. Int. J. Mol. Sci. 2023, 24, 15926. https://doi.org/10.3390/ijms242115926
Hanazaki H, Yokota H, Yamagami S, Nakamura Y, Nagaoka T. The Effect of Anti-Autotaxin Aptamers on the Development of Proliferative Vitreoretinopathy. International Journal of Molecular Sciences. 2023; 24(21):15926. https://doi.org/10.3390/ijms242115926
Chicago/Turabian StyleHanazaki, Hirotsugu, Harumasa Yokota, Satoru Yamagami, Yoshikazu Nakamura, and Taiji Nagaoka. 2023. "The Effect of Anti-Autotaxin Aptamers on the Development of Proliferative Vitreoretinopathy" International Journal of Molecular Sciences 24, no. 21: 15926. https://doi.org/10.3390/ijms242115926