

A2BAR Antagonism Decreases the Glomerular Expression and Secretion of Chemoattractants for Monocytes and the Pro-Fibrotic M2 Macrophages Polarization during Diabetic Nephropathy
Abstract
:1. Introduction
2. Results
2.1. The In Vivo Blockade of the A2BAR Alters the Expression of Immune System-Related Genes in the Glomeruli of Diabetic Nephropathy Rats
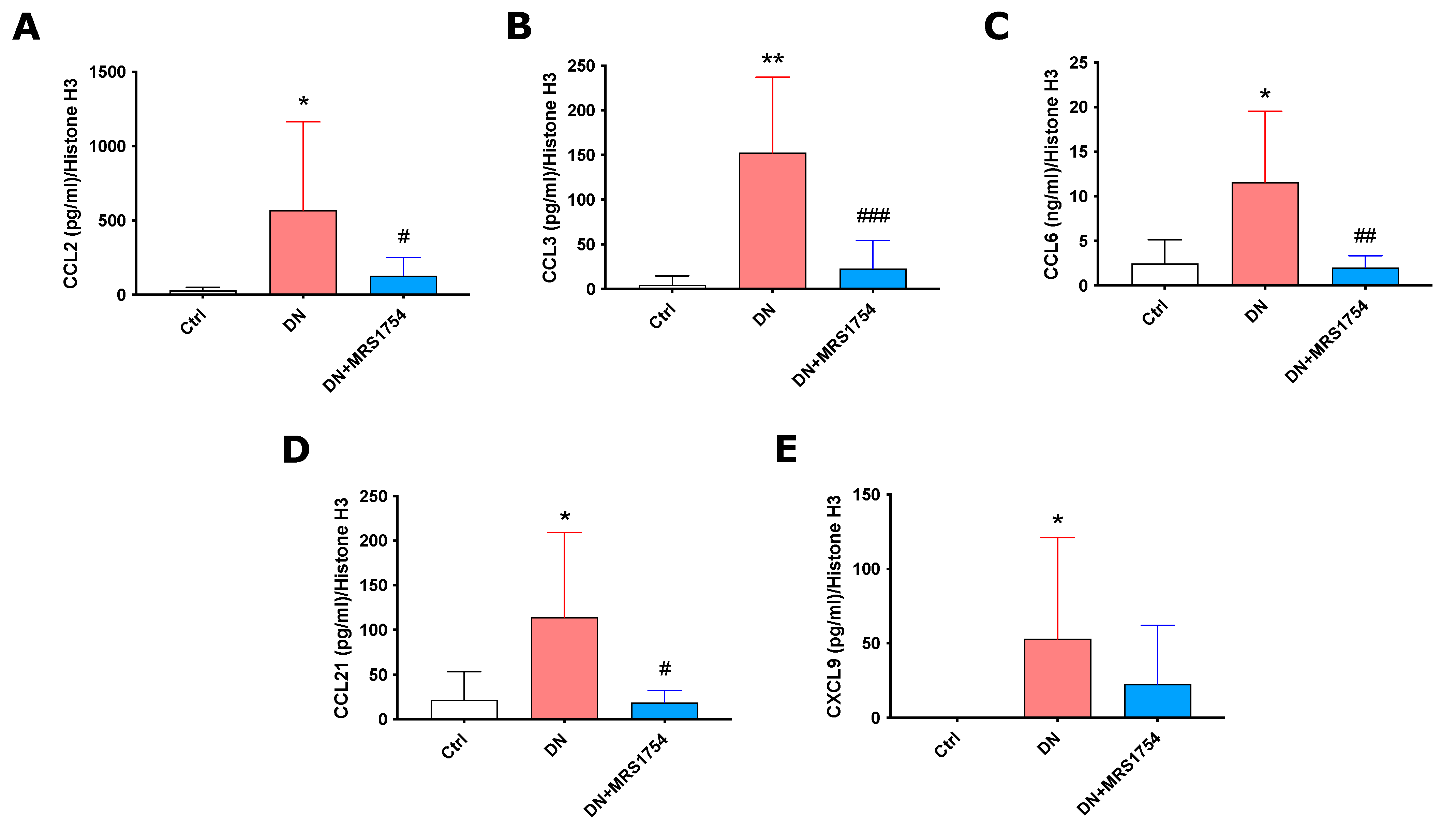
2.2. MRS1754 In Vivo Treatment Decreases the Expression and Secretion of Chemoattractants/Chemokines for Immune Cells in Rats with Diabetic Nephropathy
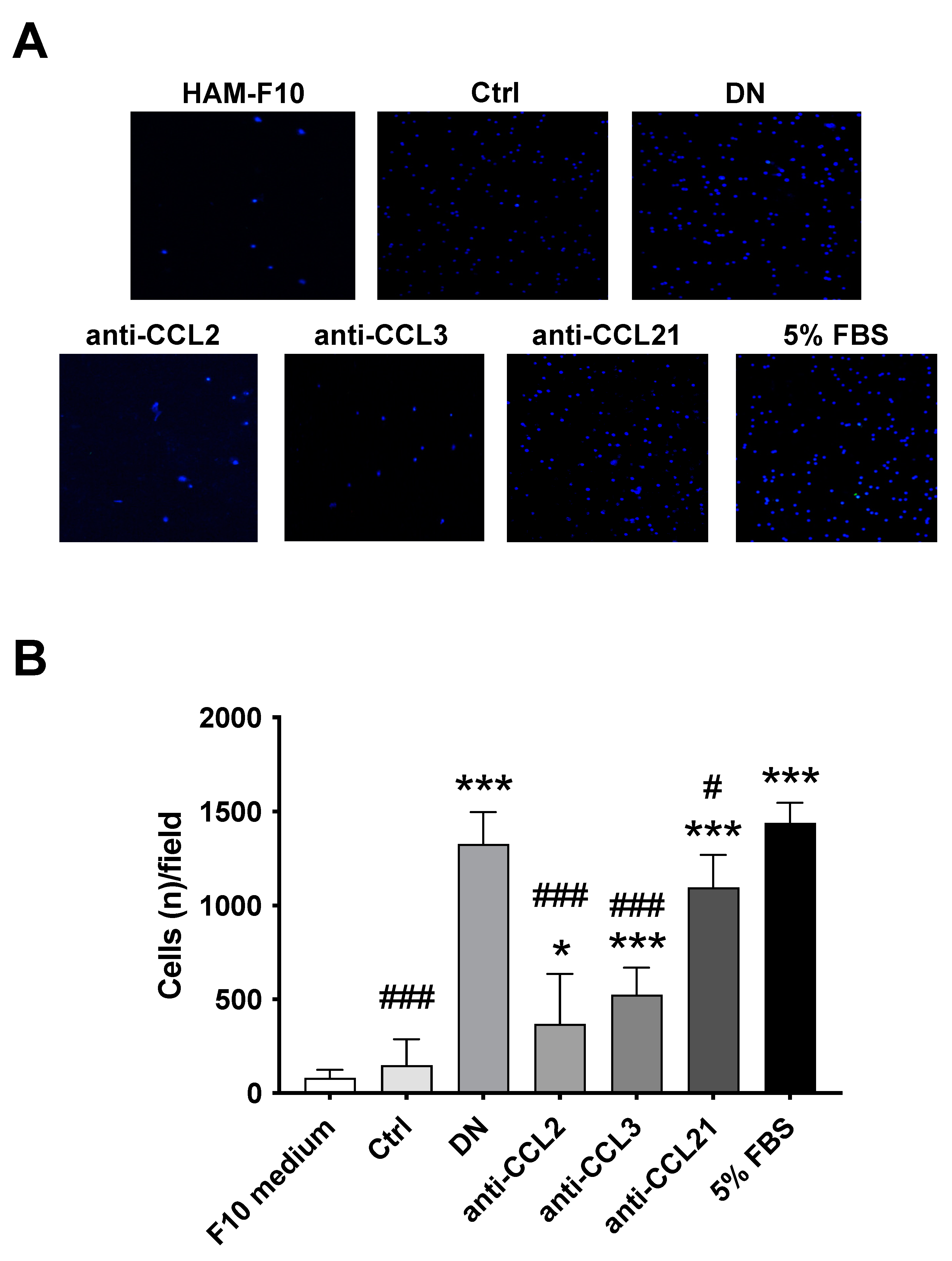
2.3. MRS1754 In Vivo Treatment Decreases the Intraglomerular Infiltration of Monocytes/Macrophages through the Reduction of Chemoattractant/Chemokine Expression/Secretion in Rats with Diabetic Nephropathy
2.4. The In Vivo Antagonism of the A2BAR Decreased M2 Macrophage Polarization in the Glomeruli of Rats with Diabetic Nephropathy
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Biopsies
4.2. Glomerulus Isolation
4.3. Transcriptomic Analysis
4.4. Immunohistochemistry and Immunohistofluorescence
4.5. ELISA
4.6. Macrophage In Vitro Cell Migration Assays
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Umanath, K.; Lewis, J.B. Update on Diabetic Nephropathy: Core Curriculum 2018. Am. J. Kidney Dis. 2018, 71, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, K.R.; Bakris, G.L.; Bilous, R.W.; Chiang, J.L.; de Boer, I.H.; Goldstein-Fuchs, J.; Hirsch, I.B.; Kalantar-Zadeh, K.; Narva, A.S.; Navaneethan, S.D.; et al. Diabetic Kidney Disease: A Report from an ADA Consensus Conference. Diabetes Care 2014, 37, 2864–2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.C.; Cooper, M.E.; Zimmet, P. Changing epidemiology of type 2 diabetes mellitus and associated chronic kidney disease. Nat. Rev. Nephrol. 2016, 12, 73–81. [Google Scholar] [CrossRef]
- Pérez-Morales, R.E.; del Pino, M.D.; Valdivielso, J.M.; Ortiz, A.; Mora-Fernández, C.; Navarro-González, J.F. Inflammation in Diabetic Kidney Disease. Nephron 2018, 143, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Qi, C.; Mao, X.; Zhang, Z.; Wu, H. Classification and Differential Diagnosis of Diabetic Nephropathy. J. Diabetes Res. 2017, 2017, 8637138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsaad, K.O.; Herzenberg, A.M. Distinguishing diabetic nephropathy from other causes of glomerulosclerosis: An update. J. Clin. Pathol. 2007, 60, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Lim, A. Diabetic nephropathy—Complications and treatment. Int. J. Nephrol. Renov. Dis. 2014, 7, 361–381. [Google Scholar] [CrossRef] [Green Version]
- Weil, E.J.; Lemley, K.V.; Mason, C.C.; Yee, B.; Jones, L.I.; Blouch, K.; Lovato, T.; Richardson, M.; Myers, B.D.; Nelson, R.G. Podocyte detachment and reduced glomerular capillary endothelial fenestration promote kidney disease in type 2 diabetic nephropathy. Kidney Int. 2012, 82, 1010–1017. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, M.; Najafian, B.; Kim, Y.; Caramori, M.L.; Mauer, M. Podocyte Detachment and Reduced Glomerular Capillary Endothelial Fenestration in Human Type 1 Diabetic Nephropathy. Diabetes 2007, 56, 2155–2160. [Google Scholar] [CrossRef] [Green Version]
- Dabla, P.K. Renal function in diabetic nephropathy. World J. Diabetes 2010, 1, 48–56. [Google Scholar] [CrossRef]
- Nevola, R.; Alfano, M.; Pafundi, P.C.; Brin, C.; Gragnano, F.; Calabrò, P.; Adinolfi, L.E.; Rinaldi, L.; Sasso, F.C.; Caturano, A. Cardiorenal Impact of SGLT-2 Inhibitors: A Conceptual Revolution in The Management of Type 2 Diabetes, Heart Failure and Chronic Kidney Disease. Rev. Cardiovasc. Med. 2022, 23, 106. [Google Scholar] [CrossRef]
- Kopel, J.; Pena-Hernandez, C.; Nugent, K. Evolving spectrum of diabetic nephropathy. World J. Diabetes 2019, 10, 269–279. [Google Scholar] [CrossRef]
- Zheng, Z.; Zheng, F. Immune Cells and Inflammation in Diabetic Nephropathy. J. Diabetes Res. 2015, 2016, 1841690. [Google Scholar] [CrossRef] [Green Version]
- Tesch, G.H. Diabetic nephropathy—Is this an immune disorder? Clin. Sci. 2017, 131, 2183–2199. [Google Scholar] [CrossRef]
- Araújo, L.S.; Torquato, B.G.S.; Da Silva, C.A.; Monteiro, M.L.G.D.R.; Martins, A.L.M.D.S.; Da Silva, M.V.; Dos Reis, M.A.; Machado, J.R. Renal expression of cytokines and chemokines in diabetic nephropathy. BMC Nephrol. 2020, 21, 308. [Google Scholar] [CrossRef]
- Wada, J.; Makino, H. Inflammation and the pathogenesis of diabetic nephropathy. Clin. Sci. 2012, 124, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Araújo, L.S.; Silva, M.V.; da Silva, C.A.; Monteiro, M.L.R.; Pereira, L.H.D.M.; Rocha, L.P.; Corrêa, R.R.M.; Reis, M.A.; Machado, J.R. Cytokines and T Helper Cells in Diabetic Nephropathy Pathogenesis. J. Diabetes Mellit. 2016, 6, 230–246. [Google Scholar] [CrossRef] [Green Version]
- Navarro-González, J.F.; Mora-Fernández, C. The Role of Inflammatory Cytokines in Diabetic Nephropathy. J. Am. Soc. Nephrol. 2008, 19, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Niemir, Z.I.; Stein, H.; Ciechanowicz, A.; Olejniczak, P.; Dworacki, G.; Ritz, E.; Waldherr, R.; Czekalski, S. The in situ expression of interleukin-8 in the normal human kidney and in different morphological forms of glomerulonephritis. Am. J. Kidney Dis. 2004, 43, 983–998. [Google Scholar] [CrossRef]
- Perlman, A.S.; Chevalier, J.M.; Wilkinson, P.; Liu, H.; Parker, T.; Levine, D.M.; Sloan, B.J.; Gong, A.; Sherman, R.; Farrell, F.X. Serum Inflammatory and Immune Mediators Are Elevated in Early Stage Diabetic Nephropathy. Ann. Clin. Lab. Sci. 2015, 45, 256–263. [Google Scholar]
- Rubio-Navarro, A.; Guerrero-Hue, M.; Martín-Fernandez, B.; Cortegano, I.; Olivares-Alvaro, E.; Heras, N.D.L.; Alía, M.; de Andrés, B.; Gaspar, M.L.; Egido, J.; et al. Phenotypic Characterization of Macrophages from Rat Kidney by Flow Cytometry. J. Vis. Exp. 2016, 116, e54599. [Google Scholar] [CrossRef]
- Meng, X.-M.; Wang, S.; Huang, X.-R.; Yang, C.; Xiao, J.; Zhang, Y.; To, K.-F.; Nikolic-Paterson, D.J.; Lan, H.-Y. Inflammatory macrophages can transdifferentiate into myofibroblasts during renal fibrosis. Cell Death Dis. 2016, 7, e2495. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Meng, X.-M.; Ng, Y.-Y.; Ma, F.Y.; Zhou, S.; Zhang, Y.; Yang, C.; Huang, X.-R.; Xiao, J.; Wang, Y.-Y.; et al. TGF-β/Smad3 signalling regulates the transition of bone marrow-derived macrophages into myofibroblasts during tissue fibrosis. Oncotarget 2015, 7, 8809–8822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolic-Paterson, D.J.; Wang, S.; Lan, H.Y. Macrophages promote renal fibrosis through direct and indirect mechanisms. Kidney Int. Suppl. 2014, 4, 34–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.; Tan, R.J.; Liu, Y. Myofibroblast in Kidney Fibrosis: Origin, Activation, and Regulation; Springer: Singapore, 2019; Volume 1165. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Wu, G.; Xiong, W.; Gu, W.; Wang, C.-Y. Macrophages: Friend or foe in idiopathic pulmonary fibrosis? Respir. Res. 2018, 19, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lech, M.; Anders, H.J. Macrophages and fibrosis: How resident and infiltrating mononuclear phagocytes orchestrate all phases of tissue injury and repair. Biochim. Biophys. Acta 2013, 1832, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.M.-K.; Nikolic-Paterson, D.J.; Lan, H.-Y. Macrophages: Versatile players in renal inflammation and fibrosis. Nat. Rev. Nephrol. 2019, 15, 144–158. [Google Scholar] [CrossRef]
- Angielski, S.; Jakubowski, Z.; Pawelczyk, T.; Piec, G.; Redlak, M. Renal Handling and Metabolism of Adenosine in Diabetic Rats1. In Diabetes and the Kidney; Karger Publishers: Basel, Switzerland, 1989; Volume 73, pp. 52–58. [Google Scholar] [CrossRef]
- Xia, J.-F.; Liang, Q.-L.; Hu, P.; Wang, Y.-M.; Li, P.; Luo, G.-A. Correlations of six related purine metabolites and diabetic nephropathy in Chinese type 2 diabetic patients. Clin. Biochem. 2009, 42, 215–220. [Google Scholar] [CrossRef]
- Neméth, Z.H.; Bleich, D.; Csóka, B.; Pacher, P.; Mabley, J.G.; Himer, L.; Vizi, E.S.; Deitch, E.A.; Szabó, C.; Cronstein, B.N.; et al. Adenosine receptor activation ameliorates type 1 diabetes. FASEB J. 2007, 21, 2379–2388. [Google Scholar] [CrossRef] [Green Version]
- Oyarzún, C.; Garrido, W.; Alarcón, S.; Yáñez, A.; Sobrevia, L.; Quezada, C.; Martín, R.S. Adenosine contribution to normal renal physiology and chronic kidney disease. Mol. Asp. Med. 2017, 55, 75–89. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, A.; Toledo, C.; Oyarzún, C.; Sepúlveda, A.; Quezada, C.; Guillén-Gómez, E.; Díaz-Encarnación, M.M.; Pastor-Anglada, M.; Martín, R.S. Adenosine A2B receptor-mediated VEGF induction promotes diabetic glomerulopathy. Lab. Investig. 2013, 93, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Roa, H.; Gajardo, C.; Troncoso, E.; Fuentealba, V.; Escudero, C.; Yáñez, A.; Sobrevia, L.; Pastor-Anglada, M.; Quezada, C.; Martin, R.S. Adenosine mediates transforming growth factor-beta 1 release in kidney glomeruli of diabetic rats. FEBS Lett. 2009, 583, 3192–3198. [Google Scholar] [CrossRef] [Green Version]
- Torres, Á.; Muñoz, K.; Nahuelpán, Y.; Saez, A.-P.R.; Mendoza, P.; Jara, C.; Cappelli, C.; Suarez, R.; Oyarzún, C.; Quezada, C.; et al. Intraglomerular Monocyte/Macrophage Infiltration and Macrophage–Myofibroblast Transition during Diabetic Nephropathy Is Regulated by the A2B Adenosine Receptor. Cells 2020, 9, 1051. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Li, G.; Yang, H.; Wang, X.; Li, J.; Zhang, W. Inhibition of Adenosine A2B Receptor Ameliorates Renal Fibrosis by Regulating Macrophage Infiltration and Polarization. Circulation 2017, 136 (Suppl. 136), A17075. [Google Scholar]
- Valladares, D.; Quezada, C.; Montecinos, P.; Concha, I.; Yañez, A.; Sobrevia, L.; Martín, R.S. Adenosine A2B receptor mediates an increase on VEGF-A production in rat kidney glomeruli. Biochem. Biophys. Res. Commun. 2008, 366, 180–185. [Google Scholar] [CrossRef]
- Tesch, G.H.; Allen, T. Rodent models of streptozotocin-induced diabetic nephropathy (Methods in Renal Research). Nephrology 2007, 12, 261–266. [Google Scholar] [CrossRef]
- Gao, Z.-G.; Jacobson, K.A. A2B Adenosine Receptor and Cancer. Int. J. Mol. Sci. 2019, 20, 5139. [Google Scholar] [CrossRef] [Green Version]
- Effendi, W.I.; Nagano, T.; Kobayashi, K.; Nishimura, Y. Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases. Cells 2020, 9, 785. [Google Scholar] [CrossRef] [Green Version]
- Franco, R.; Rivas-Santisteban, R.; Navarro, G.; Reyes-Resina, I. Adenosine Receptor Antagonists to Combat Cancer and to Boost Anti-Cancer Chemotherapy and Immunotherapy. Cells 2021, 10, 2831. [Google Scholar] [CrossRef]
- Haskó, G.; Linden, J.; Cronstein, B.; Pacher, P. Adenosine receptors: Therapeutic aspects for inflammatory and immune diseases. Nat. Rev. Drug Discov. 2008, 7, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Haskó, G.; Csóka, B.; Németh, Z.H.; Vizi, E.S.; Pacher, P. A2B adenosine receptors in immunity and inflammation. Trends Immunol. 2009, 30, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buenestado, A.; Delyle, S.G.; Arnould, I.; Besnard, F.; Naline, E.; Blouquit-Laye, S.; Chapelier, A.; Bellamy, J.; Devillier, P. The role of adenosine receptors in regulating production of tumour necrosis factor-α and chemokines by human lung macrophages. Br. J. Pharmacol. 2010, 159, 1304–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, C.L.; Luster, A.D. The Chemokine System in Innate Immunity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016303. [Google Scholar] [CrossRef] [Green Version]
- Haller, H.; Bertram, A.; Nadrowitz, F.; Menne, J. Monocyte chemoattractant protein-1 and the kidney. Curr. Opin. Nephrol. Hypertens. 2016, 25, 42–49. [Google Scholar] [CrossRef]
- Cantero-Navarro, E.; Rayego-Mateos, S.; Orejudo, M.; Tejedor-Santamaria, L.; Tejera-Muñoz, A.; Sanz, A.B.; Marquez-Exposito, L.; Marchant, V.; Santos-Sanchez, L.; Egido, J.; et al. Role of Macrophages and Related Cytokines in Kidney Disease. Front. Med. 2021, 8, 688060. [Google Scholar] [CrossRef]
- Alsheikh, A.J.; Dasinger, J.H.; Abais-Battad, J.M.; Fehrenbach, D.J.; Yang, C.; Cowley, A.W., Jr.; Mattson, D.L. CCL2 mediates early renal leukocyte infiltration during salt-sensitive hypertension. Am. J. Physiol. Ren. Physiol. 2020, 318, F982–F993. [Google Scholar] [CrossRef]
- Eardley, K.; Zehnder, D.; Quinkler, M.; Lepenies, J.; Bates, R.; Savage, C.; Howie, A.; Adu, D.; Cockwell, P. The relationship between albuminuria, MCP-1/CCL2, and interstitial macrophages in chronic kidney disease. Kidney Int. 2006, 69, 1189–1197. [Google Scholar] [CrossRef] [Green Version]
- Jia, P.; Xu, S.-J.; Wang, X.; Wu, X.; Ren, T.; Zou, Z.; Zeng, Q.; Shen, B.; Ding, X. Chemokine CCL2 from proximal tubular epithelial cells contributes to sepsis-induced acute kidney injury. Am. J. Physiol. Physiol. 2022, 323, F107–F119. [Google Scholar] [CrossRef]
- de Jager, S.C.; Bot, I.; Kraaijeveld, A.O.; Korporaal, S.J.; Bot, M.; van Santbrink, P.J.; van Berkel, T.J.; Kuiper, J.; Biessen, E.A. Leukocyte-Specific CCL3 Deficiency Inhibits Atherosclerotic Lesion Development by Affecting Neutrophil Accumulation. Arter. Thromb. Vasc. Biol. 2013, 33, e75–e83. [Google Scholar] [CrossRef] [Green Version]
- Lindell, D.M.; Standiford, T.J.; Mancuso, P.; Leshen, Z.J.; Huffnagle, G.B. Macrophage Inflammatory Protein 1α/CCL3 Is Required for Clearance of an Acute Klebsiella pneumoniae Pulmonary Infection. Infect. Immun. 2001, 69, 6364–6369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipriani, B.; Borsellino, G.; Poccia, F.; Placido, R.; Tramonti, D.; Bach, S.; Battistini, L.; Brosnan, C.F. Activation of C-C beta-chemokines in human peripheral blood gammadelta T cells by isopentenyl pyrophosphate and regulation by cyto-kines. Blood 2000, 95, 39–47. [Google Scholar] [CrossRef]
- Greenberger, M.J.; Strieter, R.M.; Kunkel, S.L.; Danforth, J.M.; Laichalk, L.L.; McGillicuddy, D.C.; Standiford, T.J. Neutralization of Macrophage Inflammatory Protein-2 Attenuates Neutrophil Recruitment and Bacterial Clearance in Murine Klebsiella Pneumonia. J. Infect. Dis. 1996, 173, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Danforth, J.M.; Strieter, R.M.; Kunkel, S.L.; Arenberg, D.; VanOtteren, G.M.; Standiford, T.J. Macrophage Inflammatory Protein-1α Expression in Vivo and in Vitro: The Role of Lipoteichoic Acid. Clin. Immunol. Immunopathol. 1995, 74, 77–83. [Google Scholar] [CrossRef]
- Davatelis, G.; Tekamp-Olson, P.; Wolpe, S.D.; Hermsen, K.; Luedke, C.; Gallegos, C.; Coit, D.; Merryweather, J.; Cerami, A. Cloning and characterization of a cDNA for murine macrophage inflammatory protein (MIP), a novel monokine with inflammatory and chemokinetic properties. J. Exp. Med. 1988, 167, 1939–1944. [Google Scholar] [CrossRef] [Green Version]
- Sherry, B.; Tekamp-Olson, P.; Gallegos, C.; Bauer, D.; Davatelis, G.; Wolpe, S.D.; Masiarz, F.; Coit, D.; Cerami, A. Resolution of the two components of macrophage inflammatory protein 1, and cloning and characterization of one of those components, macrophage inflammatory protein 1 beta. J. Exp. Med. 1988, 168, 2251–2259. [Google Scholar] [CrossRef]
- Orlofsky, A.; Lin, E.Y.; Prystowsky, M.B. Selective induction of the beta chemokine C10 by IL-4 in mouse macro-phages. J. Immunol. 1994, 152, 5084–5091. [Google Scholar] [CrossRef]
- Zhang, C.; Yi, W.; Li, F.; Du, X.; Wang, H.; Wu, P.; Peng, C.; Luo, M.; Hua, W.; Wong, C.C.; et al. Eosinophil-derived CCL-6 impairs hematopoietic stem cell homeostasis. Cell Res. 2018, 28, 323–335. [Google Scholar] [CrossRef]
- Yi, F.; Jaffe, R.; Prochownik, E.V. The CCL6 chemokine is differentially regulated by c-Myc and L-Myc, and promotes tumorigenesis and metastasis. Cancer Res. 2003, 63, 2923–2932. [Google Scholar]
- Quatredeniers, M.; Bienaimé, F.; Ferri, G.; Isnard, P.; Porée, E.; Billot, K.; Birgy, E.; Mazloum, M.; Ceccarelli, S.; Silbermann, F.; et al. The renal inflammatory network of nephronophthisis. Hum. Mol. Genet. 2022, 31, 2121–2136. [Google Scholar] [CrossRef]
- Jiang, Y.; Cai, C.; Zhang, P.; Luo, Y.; Guo, J.; Li, J.; Rong, R.; Zhang, Y.; Zhu, T. Transcriptional profile changes after treatment of ischemia reperfusion injury-induced kidney fibrosis with 18β-glycyrrhetinic acid. Ren. Fail. 2022, 44, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Wada, T.; Yokoyama, H.; Lipp, M.; Ueha, S.; Matsushima, K.; Kaneko, S. Secondary lymphoid tissue chemokine (SLC/CCL21)/CCR7 signaling regulates fibrocytes in renal fibrosis. Proc. Natl. Acad. Sci. USA 2006, 103, 14098–14103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.J.; Bowman, E.P.; Murphy, K.; Youngman, K.R.; Siani, M.A.; Thompson, D.; Wu, L.; Zlotnik, A.; Butcher, E.C. 6-C-kine (SLC), a Lymphocyte Adhesion-triggering Chemokine Expressed by High Endothelium, Is an Agonist for the MIP-3β Receptor CCR7. J. Cell Biol. 1998, 141, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Ogata, M.; Zhang, Y.Y.; Wang, Y.; Itakura, M.; Harada, A.; Hashimoto, S.; Matsushima, K. Chemotactic response toward chemokines and its regulation by transforming growth factor-beta1 of murine bone marrow hematopoietic progenitor cell-derived different subset of dendritic cells. Blood 1999, 93, 3225–3232. [Google Scholar] [CrossRef] [PubMed]
- Van Raemdonck, K.; Umar, S.; Palasiewicz, K.; Volkov, S.; Volin, M.V.; Arami, S.; Chang, H.J.; Zanotti, B.; Sweiss, N.; Shahrara, S. CCL21/CCR7 signaling in macrophages promotes joint inflammation and Th17-mediated osteoclast formation in rheumatoid arthritis. Cell. Mol. Life Sci. 2019, 77, 1387–1399. [Google Scholar] [CrossRef]
- Cosway, E.J.; Ohigashi, I.; Schauble, K.; Parnell, S.M.; Jenkinson, W.E.; Luther, S.; Takahama, Y.; Anderson, G. Formation of the Intrathymic Dendritic Cell Pool Requires CCL21-Mediated Recruitment of CCR7+ Progenitors to the Thymus. J. Immunol. 2018, 201, 516–523. [Google Scholar] [CrossRef] [Green Version]
- Gunn, M.D.; Kyuwa, S.; Tam, C.; Kakiuchi, T.; Matsuzawa, A.; Williams, L.T.; Nakano, H. Mice Lacking Expression of Secondary Lymphoid Organ Chemokine Have Defects in Lymphocyte Homing and Dendritic Cell Localization. J. Exp. Med. 1999, 189, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Abe, R.; Donnelly, S.C.; Peng, T.; Bucala, R.; Metz, C.N. Peripheral Blood Fibrocytes: Differentiation Pathway and Migration to Wound Sites. J. Immunol. 2001, 166, 7556–7562. [Google Scholar] [CrossRef] [Green Version]
- Xuan, W.; Qu, Q.; Zheng, B.; Xiong, S.; Fan, G.-H. The chemotaxis of M1 and M2 macrophages is regulated by different chemokines. J. Leukoc. Biol. 2014, 97, 61–69. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Y.; Nguyen, H.; Koupenova, M.; Chauhan, A.; Makitalo, M.; Jones, M.; Hilaire, C.S.; Seldin, D.C.; Toselli, P.; et al. The A2B adenosine receptor protects against inflammation and excessive vascular adhesion. J. Clin. Investig. 2006, 116, 1913–1923. [Google Scholar] [CrossRef] [Green Version]
- Haskó, G.; Pacher, P. Regulation of Macrophage Function by Adenosine. Arter. Thromb. Vasc. Biol. 2012, 32, 865–869. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Yang, D.; Carroll, S.H.; Eltzschig, H.K.; Ravid, K. Activation of the macrophage A2b adenosine receptor regulates tumor necrosis factor–α levels following vascular injury. Exp. Hematol. 2009, 37, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Johnston-Cox, H.; Eisenstein, A.S.; Koupenova, M.; Carroll, S.; Ravid, K. The Macrophage A2b Adenosine Receptor Regulates Tissue Insulin Sensitivity. PLoS ONE 2014, 9, e98775. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Arora, S.; Dev, K.; Agarwal, B.; Das, P.; Syed, M.A. Macrophages: Their role, activation and polarization in pulmonary diseases. Immunobiology 2017, 223, 383–396. [Google Scholar] [CrossRef]
- Calle, P.; Hotter, G. Macrophage Phenotype and Fibrosis in Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 2806. [Google Scholar] [CrossRef] [Green Version]
- Landis, R.C.; Quimby, K.R.; Greenidge, A.R. M1/M2 Macrophages in Diabetic Nephropathy: Nrf2/HO-1 as Therapeutic Targets. Curr. Pharm. Des. 2018, 24, 2241–2249. [Google Scholar] [CrossRef]
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.; Ruslami, R. Type 2 Diabetes and its Impact on the Immune System. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef]
- Daryabor, G.; Atashzar, M.R.; Kabelitz, D.; Meri, S.; Kalantar, K. The Effects of Type 2 Diabetes Mellitus on Organ Metabolism and the Immune System. Front. Immunol. 2020, 11, 1582. [Google Scholar] [CrossRef]
- Rubinstein, R.; Genaro, A.M.; Motta, A.; Cremaschi, G.; Wald, M.R. Impaired immune responses in streptozotocin-induced type I diabetes in mice. Involvement of high glucose. Clin. Exp. Immunol. 2008, 154, 235–246. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Genaro, A.M.; Wald, M.R. Differential effect of hyperglycaemia on the immune response in an experimental model of diabetes in BALB/cByJ and C57Bl/6J mice: Participation of oxidative stress. Clin. Exp. Immunol. 2013, 171, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafar, N.; Edriss, H.; Nugent, K. The Effect of Short-Term Hyperglycemia on the Innate Immune System. Am. J. Med. Sci. 2016, 351, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Lisco, G.; Giagulli, V.A.; De Pergola, G.; Guastamacchia, E.; Jirillo, E.; Triggiani, V. Hyperglycemia-induced Immune System Disorders in Diabetes Mellitus and the Concept of Hyperglycemic Memory of Innate Immune Cells: A perspective. Endocr. Metab. Immune Disord.-Drug Targets 2022, 22, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Megawati, E.R.; Meutia, N.; Lubis, L.D. The effect of hyperglycaemia on the macrophages in the cell culture. Folia Morphol. 2022, 81, 387–393. [Google Scholar] [CrossRef]
- Liu, Y.J.; Saini, A.; Cohen, D.J.; Ooi, B.S. Modulation of macrophage proliferation by hyperglycemia. Mol. Cell. Endocrinol. 1995, 114, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, S.; Lindsay, J.; Ingram, R.; Xu, H.; Chen, M. Sustained high glucose exposure sensitizes macrophage responses to cytokine stimuli but reduces their phagocytic activity. BMC Immunol. 2018, 19, 24. [Google Scholar] [CrossRef] [Green Version]
- Orliaguet, L.; Dalmas, E.; Drareni, K.; Venteclef, N.; Alzaid, F. Mechanisms of Macrophage Polarization in Insulin Signaling and Sensitivity. Front. Endocrinol. 2020, 11, 62. [Google Scholar] [CrossRef] [Green Version]
- Szondy, Z.; Garabuczi, E.; Joós, G.; Tsay, G.J.; Sarang, Z. Impaired Clearance of Apoptotic Cells in Chronic Inflammatory Diseases: Therapeutic Implications. Front. Immunol. 2014, 5, 354. [Google Scholar] [CrossRef] [Green Version]
- Khanna, S.; Biswas, S.; Shang, Y.; Collard, E.; Azad, A.; Kauh, C.; Bhasker, V.; Gordillo, G.M.; Sen, C.K.; Roy, S. Macrophage Dysfunction Impairs Resolution of Inflammation in the Wounds of Diabetic Mice. PLoS ONE 2010, 5, e9539. [Google Scholar] [CrossRef] [Green Version]
- Csóka, B.; Selmeczy, Z.; Koscsó, B.; Németh, Z.H.; Pacher, P.; Murray, P.J.; Kepka-Lenhart, D.; Morris, S.M., Jr.; Gause, W.C.; Leibovich, S.J.; et al. Adenosine promotes alternative macrophage activation via A2A and A2B receptors. FASEB J. 2011, 26, 376–386. [Google Scholar] [CrossRef] [Green Version]
- Alarcón, S.; Garrido, W.; Vega, G.; Cappelli, C.; Suárez, R.; Oyarzún, C.; Quezada, C.; Martín, R.S. Deficient Insulin-mediated Upregulation of the Equilibrative Nucleoside Transporter 2 Contributes to Chronically Increased Adenosine in Diabetic Glomerulopathy. Sci. Rep. 2017, 7, 9439. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway Name | Entities | Reactions | ||||
|---|---|---|---|---|---|---|
| Found | Ratio | p-Value | FDR | Found | Ratio | |
| Interleukin-33 signaling | 2/6 | 2.67 × 10−4 | 9.50 × 10−4 | 0.752 | 2/2 | 1.41 × 10−4 |
| Chemokine receptors bind chemokines | 4/102 | 0.005 | 0.007 | 0.752 | 4/19 | 0.001 |
| Interleukin-10 signaling | 5/175 | 0.008 | 0.01 | 0.752 | 1/15 | 0.001 |
| Interleukin receptor SHC signaling | 2/32 | 0.001 | 0.024 | 0.752 | 6/6 | 4.23 × 10−4 |
| Terminal pathway of complement | 1/9 | 4.01 × 10−4 | 0.064 | 0.752 | 4/5 | 3.52 × 10−4 |
| Peptide ligand-binding receptors | 12/515 | 0.023 | 0.089 | 0.752 | 12/83 | 0.006 |
| CASP8 activity is inhibited | 1/13 | 5.79 × 10−4 | 0.092 | 0.752 | 2/2 | 1.41 × 10−4 |
| Pathway Name | Entities | Reactions | ||||
|---|---|---|---|---|---|---|
| Found | Ratio | p-Value | FDR | Found | Ratio | |
| Neutrophil degranulation | 45/478 | 0.021 | 5.57 × 10−11 | 8.60 × 10−8 | 10/10 | 7.04 × 10−4 |
| PD-1 signaling | 6/34 | 0.002 | 6.86 × 10−4 | 0.446 | 2/5 | 3.52 × 10−4 |
| IRAK deficiency (TRL2/4) | 6/36 | 0.002 | 9.19 × 10−4 | 0.446 | 2/2 | 1.41 × 10−4 |
| Interleukin-10 signaling | 17/175 | 0.008 | 0.001 | 0.446 | 13/15 | 0.001 |
| RUNX3 Regulates Immune Response and Cell Migration | 4/19 | 8.46 × 10−4 | 0.003 | 0.799 | 2/5 | 3.52 × 10−4 |
| MyD88 deficiency (TLR2/4) | 6/46 | 0.002 | 0.003 | 0.799 | 2/2 | 1.41 × 10−4 |
| Translocation of ZAP-70 to Immunological synapse | 5/40 | 0.002 | 0.008 | 0.963 | 4/4 | 2.82 × 10−4 |
| Cross-presentation of particulate exogenous antigens (phagosomes) | 3/14 | 6.24 × 10−4 | 0.009 | 0.963 | 3/3 | 2.11 × 10−4 |
| Immunoregulatory interactions between a Lymphoid and a non- Lymphoid cell | 40/599 | 0.027 | 0.011 | 0.963 | 29/44 | 0.003 |
| Phosphorylation of CD3 and TCR zeta chains | 6/60 | 0.003 | 0.011 | 0.963 | 7/7 | 4.93 × 10−4 |
| Regulation of Complement cascade | 20/234 | 0.01 | 0.029 | 0.963 | 35/42 | 0.003 |
| Complement cascade | 21/260 | 0.012 | 0.03 | 0.963 | 52/72 | 0.005 |
| Alternative complement activation | 4/22 | 9.80 × 10−4 | 0.03 | 0.963 | 9/9 | 6.34 × 10−4 |
| Chemokine receptors bind chemokines | 10/102 | 0.005 | 0.038 | 0.963 | 9/19 | 0.001 |
| Trafficking and processing of endosomal TLR | 5/61 | 0.003 | 0.04 | 0.963 | 7/7 | 4.93 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Arévalo, Á.; Nahuelpán, Y.; Muñoz, K.; Jara, C.; Cappelli, C.; Taracha-Wiśniewska, A.; Quezada-Monrás, C.; Martín, R.S. A2BAR Antagonism Decreases the Glomerular Expression and Secretion of Chemoattractants for Monocytes and the Pro-Fibrotic M2 Macrophages Polarization during Diabetic Nephropathy. Int. J. Mol. Sci. 2023, 24, 10829. https://doi.org/10.3390/ijms241310829
Torres-Arévalo Á, Nahuelpán Y, Muñoz K, Jara C, Cappelli C, Taracha-Wiśniewska A, Quezada-Monrás C, Martín RS. A2BAR Antagonism Decreases the Glomerular Expression and Secretion of Chemoattractants for Monocytes and the Pro-Fibrotic M2 Macrophages Polarization during Diabetic Nephropathy. International Journal of Molecular Sciences. 2023; 24(13):10829. https://doi.org/10.3390/ijms241310829
Chicago/Turabian StyleTorres-Arévalo, Ángelo, Yéssica Nahuelpán, Katherin Muñoz, Claudia Jara, Claudio Cappelli, Agnieszka Taracha-Wiśniewska, Claudia Quezada-Monrás, and Rody San Martín. 2023. "A2BAR Antagonism Decreases the Glomerular Expression and Secretion of Chemoattractants for Monocytes and the Pro-Fibrotic M2 Macrophages Polarization during Diabetic Nephropathy" International Journal of Molecular Sciences 24, no. 13: 10829. https://doi.org/10.3390/ijms241310829