
Exploring the Effects of Human Bone Marrow-Derived Mononuclear Cells on Angiogenesis In Vitro



Abstract
:1. Introduction
2. Results
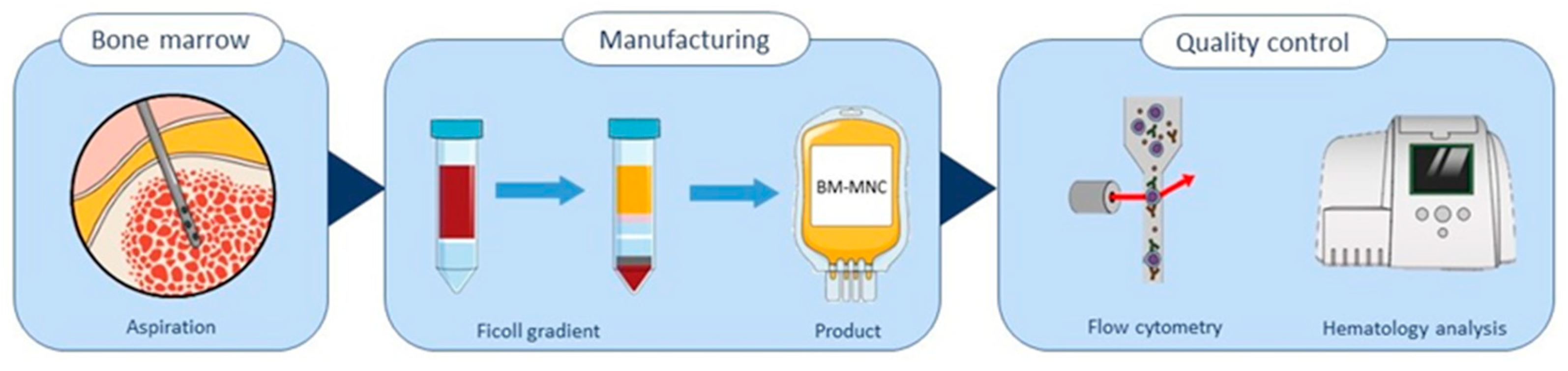
2.1. Isolation and Quality Control of Bone Marrow-Derived Cells
2.2. BM-MNCs Have No Effect on Endothelial Cell Proliferation
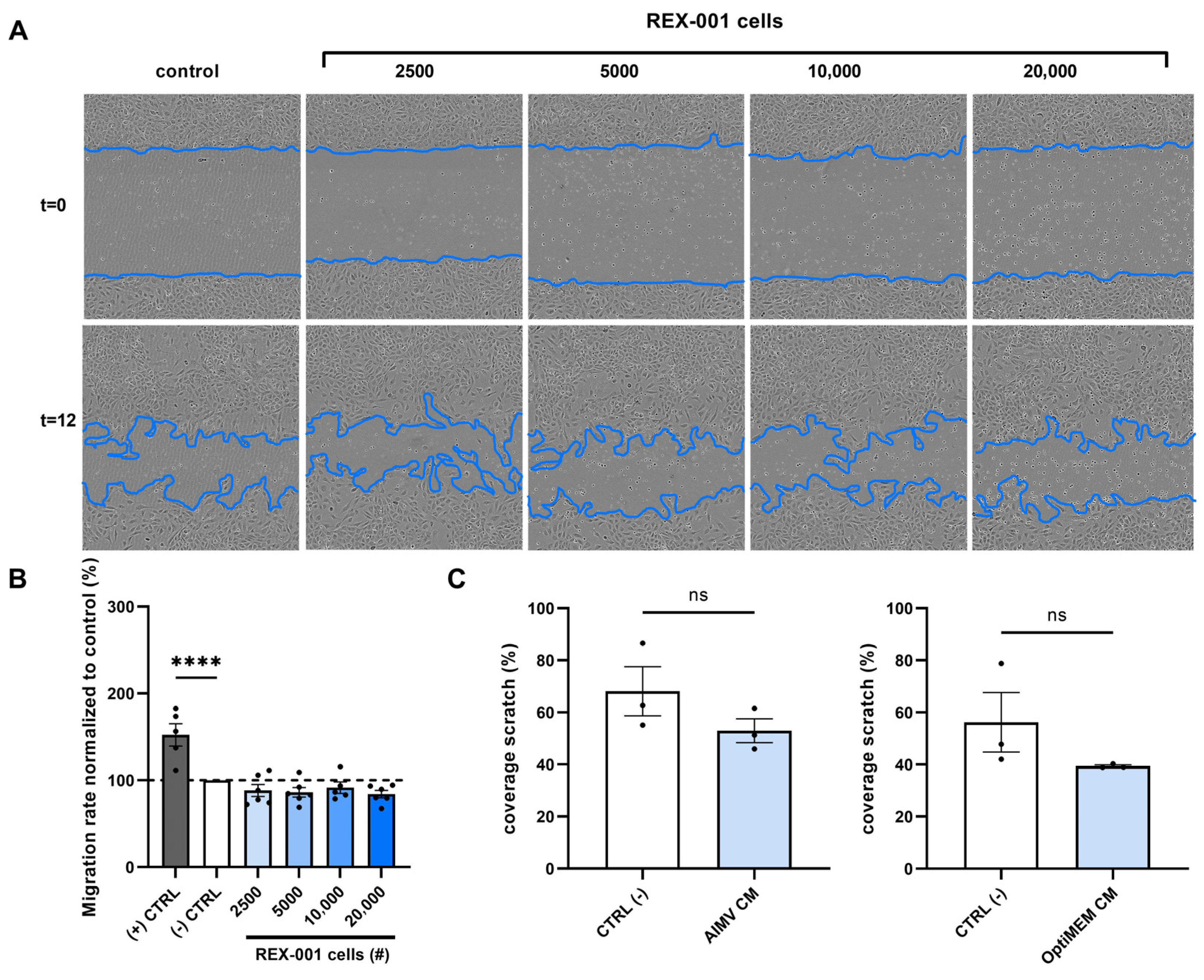
2.3. BM-MNCs Do Not Affect Endothelial Cell Migration
2.4. No Effect of BM-MNCs on Endothelial Cell Tube Formation
2.5. The Effect of BM-MNCs on Aortic Ring Sprouting
2.6. BM-MNCs Release Angiogenic Cytokines
3. Discussion
4. Materials and Methods
4.1. BM-MNC Isolates Manufacturing
4.2. Flow Cytometric Analysis
4.3. Cell Culture of Human Umbilical Vein Endothelial Cells (HUVECs)
4.4. BM-MNC Conditioned Medium
4.5. MTT Assay
4.6. Scratch Wound Healing Assay
4.7. Tube Formation Assay
4.8. Aortic Ring Assay
4.9. ELISA
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tateishi-Yuyama, E.; Matsubara, H.; Murohara, T.; Ikeda, U.; Shintani, S.; Masaki, H.; Amano, K.; Kishimoto, Y.; Yoshimoto, K.; Akashi, H.; et al. Therapeutic angiogenesis for patients with limb ischaemia by autologous transplantation of bone-marrow cells: A pilot study and a randomised controlled trial. Lancet 2002, 360, 427–435. [Google Scholar] [CrossRef]
- Yusoff, F.M.; Kajikawa, M.; Matsui, S.; Hashimoto, H.; Kishimoto, S.; Maruhashi, T.; Chowdhury, M.; Noma, K.; Nakashima, A.; Kihara, Y.; et al. Review of the Long-term Effects of Autologous Bone-Marrow Mononuclear Cell Implantation on Clinical Outcomes in Patients with Critical Limb Ischemia. Sci. Rep. 2019, 9, 7711. [Google Scholar] [CrossRef]
- Matoba, S.; Tatsumi, T.; Murohara, T.; Imaizumi, T.; Katsuda, Y.; Ito, M.; Saito, Y.; Uemura, S.; Suzuki, H.; Fukumoto, S.; et al. Long-term clinical outcome after intramuscular implantation of bone marrow mononuclear cells (Therapeutic Angiogenesis by Cell Transplantation [TACT] trial) in patients with chronic limb ischemia. Am. Heart J. 2008, 156, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Dash, N.R.; Dash, S.N.; Routray, P.; Mohapatra, S.; Mohapatra, P.C. Targeting nonhealing ulcers of lower extremity in human through autologous bone marrow-derived mesenchymal stem cells. Rejuvenation Res. 2009, 12, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Procházka, V.; Gumulec, J.; Jalůvka, F.; Salounová, D.; Jonszta, T.; Czerný, D.; Krajča, J.; Urbanec, R.; Klement, P.; Martinek, J.; et al. Cell therapy, a new standard in management of chronic critical limb ischemia and foot ulcer. Cell Transplant. 2010, 19, 1413–1424. [Google Scholar] [CrossRef]
- Sharma, S.; Pandey, N.N.; Sinha, M.; Kumar, S.; Jagia, P.; Gulati, G.S.; Gond, K.; Mohanty, S.; Bhargava, B. Randomized, Double-Blind, Placebo-Controlled Trial to Evaluate Safety and Therapeutic Efficacy of Angiogenesis Induced by Intraarterial Autologous Bone Marrow-Derived Stem Cells in Patients with Severe Peripheral Arterial Disease. J. Vasc. Interv. Radiol. 2021, 32, 157–163. [Google Scholar] [CrossRef]
- Rigato, M.; Monami, M.; Fadini, G.P. Autologous Cell Therapy for Peripheral Arterial Disease: Systematic Review and Meta-Analysis of Randomized, Nonrandomized, and Noncontrolled Studies. Circ. Res. 2017, 120, 1326–1340. [Google Scholar] [CrossRef]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Kondo, T.; Inoue, N.; Aoki, M.; Mizuno, M.; Komori, K.; Yoshida, J.; Murohara, T. Combination of in vivo angiopoietin-1 gene transfer and autologous bone marrow cell implantation for functional therapeutic angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1465–1472. [Google Scholar] [CrossRef]
- Ruiz-Salmeron, R.; de la Cuesta-Diaz, A.; Constantino-Bermejo, M.; Pérez-Camacho, I.; Marcos-Sánchez, F.; Hmadcha, A.; Soria, B. Angiographic demonstration of neoangiogenesis after intra-arterial infusion of autologous bone marrow mononuclear cells in diabetic patients with critical limb ischemia. Cell Transplant. 2011, 20, 1629–1639. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis. Annu. Rev. Med. 2006, 57, 1–18. [Google Scholar] [CrossRef]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef]
- Li, A.; Dubey, S.; Varney, M.L.; Dave, B.J.; Singh, R.K. IL-8 directly enhanced endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated angiogenesis. J. Immunol. 2003, 170, 3369–3376. [Google Scholar] [CrossRef]
- Ashtar Nakhaei, N.; Najarian, A.; Farzaei, M.H.; Norooznezhad, A.H. Endothelial dysfunction and angiogenesis: What is missing from COVID-19 and cannabidiol story? J. Cannabis. Res. 2022, 4, 21. [Google Scholar] [CrossRef]
- Herbert, S.P.; Stainier, D.Y. Molecular control of endothelial cell behaviour during blood vessel morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, regulation, and involvement in disease. Int. Immunopharmacol. 2021, 101, 107598. [Google Scholar] [CrossRef] [PubMed]
- Avraham-Davidi, I.; Yona, S.; Grunewald, M.; Landsman, L.; Cochain, C.; Silvestre, J.S.; Mizrahi, H.; Faroja, M.; Strauss-Ayali, D.; Mack, M.; et al. On-site education of VEGF-recruited monocytes improves their performance as angiogenic and arteriogenic accessory cells. J. Exp. Med. 2013, 210, 2611–2625. [Google Scholar] [CrossRef]
- Rojas-Torres, M.; Jiménez-Palomares, M.; Martín-Ramírez, J.; Beltrán-Camacho, L.; Sánchez-Gomar, I.; Eslava-Alcon, S.; Rosal-Vela, A.; Gavaldá, S.; Durán-Ruiz, M.C. REX-001, a BM-MNC Enriched Solution, Induces Revascularization of Ischemic Tissues in a Murine Model of Chronic Limb-Threatening Ischemia. Front. Cell Dev. Biol. 2020, 8, 602837. [Google Scholar] [CrossRef] [PubMed]
- Nicosia, R.F.; Ottinetti, A. Growth of microvessels in serum-free matrix culture of rat aorta. A quantitative assay of angiogenesis in vitro. Lab. Invest. 1990, 63, 115–122. [Google Scholar]
- Nowak-Sliwinska, P.; Alitalo, K.; Allen, E.; Anisimov, A.; Aplin, A.C.; Auerbach, R.; Augustin, H.G.; Bates, D.O.; van Beijnum, J.R.; Bender, R.H.F.; et al. Consensus guidelines for the use and interpretation of angiogenesis assays. Angiogenesis 2018, 21, 425–532. [Google Scholar] [CrossRef]
- Samura, M.; Hosoyama, T.; Takeuchi, Y.; Ueno, K.; Morikage, N.; Hamano, K. Therapeutic strategies for cell-based neovascularization in critical limb ischemia. J. Transl. Med. 2017, 15, 49. [Google Scholar] [CrossRef]
- Hart, C.A.; Tsui, J.; Khanna, A.; Abraham, D.J.; Baker, D.M. Stem cells of the lower limb: Their role and potential in management of critical limb ischemia. Exp. Biol. Med. 2013, 238, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Magenta, A.; Florio, M.C.; Ruggeri, M.; Furgiuele, S. Autologous cell therapy in diabetes-associated critical limb ischemia: From basic studies to clinical outcomes (Review). Int. J. Mol. Med. 2021, 48, 173. [Google Scholar] [CrossRef] [PubMed]
- Kucia, M.; Reca, R.; Jala, V.R.; Dawn, B.; Ratajczak, J.; Ratajczak, M.Z. Bone marrow as a home of heterogenous populations of nonhematopoietic stem cells. Leukemia 2005, 19, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Boulais, P.E.; Mizoguchi, T.; Zimmerman, S.; Nakahara, F.; Vivié, J.; Mar, J.C.; van Oudenaarden, A.; Frenette, P.S. The Majority of CD45(−) Ter119(−) CD31(−) Bone Marrow Cell Fraction Is of Hematopoietic Origin and Contains Erythroid and Lymphoid Progenitors. Immunity 2018, 49, 627–639. [Google Scholar] [CrossRef]
- Sahoo, S.; Klychko, E.; Thorne, T.; Misener, S.; Schultz, K.M.; Millay, M.; Ito, A.; Liu, T.; Kamide, C.; Agrawal, H.; et al. Exosomes from human CD34(+) stem cells mediate their proangiogenic paracrine activity. Circ. Res. 2011, 109, 724–728. [Google Scholar] [CrossRef]
- van Solingen, C.; Seghers, L.; Bijkerk, R.; Duijs, J.M.; Roeten, M.K.; van Oeveren-Rietdijk, A.M.; Baelde, H.J.; Monge, M.; Vos, J.B.; de Boer, H.C.; et al. Antagomir-mediated silencing of endothelial cell specific microRNA-126 impairs ischemia-induced angiogenesis. J. Cell Mol. Med. 2009, 13, 1577–1585. [Google Scholar] [CrossRef]
- Shi, J.; Wei, P.K. Interleukin-8: A potent promoter of angiogenesis in gastric cancer. Oncol. Lett. 2016, 11, 1043–1050. [Google Scholar] [CrossRef]
- van Hinsbergh, V.W.; Engelse, M.A.; Quax, P.H. Pericellular proteases in angiogenesis and vasculogenesis. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 716–728. [Google Scholar] [CrossRef]
- Corliss, B.A.; Azimi, M.S.; Munson, J.M.; Peirce, S.M.; Murfee, W.L. Macrophages: An Inflammatory Link Between Angiogenesis and Lymphangiogenesis. Microcirculation 2016, 23, 95–121. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar] [PubMed]
- Fields, G.B. Mechanisms of Action of Novel Drugs Targeting Angiogenesis-Promoting Matrix Metalloproteinases. Front. Immunol. 2019, 10, 1278. [Google Scholar] [CrossRef] [PubMed]
- Kavurma, M.M.; Bursill, C.; Stanley, C.P.; Passam, F.; Cartland, S.P.; Patel, S.; Loa, J.; Figtree, G.A.; Golledge, J.; Aitken, S.; et al. Endothelial cell dysfunction: Implications for the pathogenesis of peripheral artery disease. Front. Cardiovasc. Med. 2022, 9, 1054576. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Wang, S.; Zuo, A.; Zhang, J.; Wen, W.; Jiang, W.; Chen, H.; Liang, D.; Sun, J.; Wang, M. HIF-1α/JMJD1A signaling regulates inflammation and oxidative stress following hyperglycemia and hypoxia-induced vascular cell injury. Cell. Mol. Biol. Lett. 2021, 26, 40. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process Performance Indicator | BM-MNCs Relative to Bone Marrow (%) | Acceptance Criteria (%) |
|---|---|---|
| Leukocyte recovery | 13.05 *–22.28 | >15% |
| Erythrocyte depletion | 99.91–100 | >96% |
| Thrombocyte depletion | 95.79–100 | >60% |
| Cell population | BM-MNCs (%) | |
| Viability | 99.50–99.90 | >80% |
| Leukocytes (CD45+) 1 | 63.63–86.87 | - |
| B lymphocytes (CD19+) | 1.08–4.63 | - |
| T lymphocytes (CD3+) | 4.84–9.74 | - |
| CD4+ T lymphocytes | 2.52–4.68 | - |
| CD8+ T lymphocytes | 1.36–3.42 | - |
| Granulocytes (CD16+, CD14−) | 68.23–84.40 | >30% |
| Monocytes (CD14+) | 2.76–10.50 | - |
| CD34+ leukocytes | 1.34–3.59 | >0.1% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peeters, J.A.H.M.; Peters, H.A.B.; Videler, A.J.; Hamming, J.F.; Schepers, A.; Quax, P.H.A. Exploring the Effects of Human Bone Marrow-Derived Mononuclear Cells on Angiogenesis In Vitro. Int. J. Mol. Sci. 2023, 24, 13822. https://doi.org/10.3390/ijms241813822
Peeters JAHM, Peters HAB, Videler AJ, Hamming JF, Schepers A, Quax PHA. Exploring the Effects of Human Bone Marrow-Derived Mononuclear Cells on Angiogenesis In Vitro. International Journal of Molecular Sciences. 2023; 24(18):13822. https://doi.org/10.3390/ijms241813822
Chicago/Turabian StylePeeters, Judith A. H. M., Hendrika A. B. Peters, Anique J. Videler, Jaap F. Hamming, Abbey Schepers, and Paul H. A. Quax. 2023. "Exploring the Effects of Human Bone Marrow-Derived Mononuclear Cells on Angiogenesis In Vitro" International Journal of Molecular Sciences 24, no. 18: 13822. https://doi.org/10.3390/ijms241813822