
Effects of Different Sources of Culture Substrate on the Growth and Immune Performance of the Red Swamp Crayfish (Procambarus clarkii)
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Growth Performance
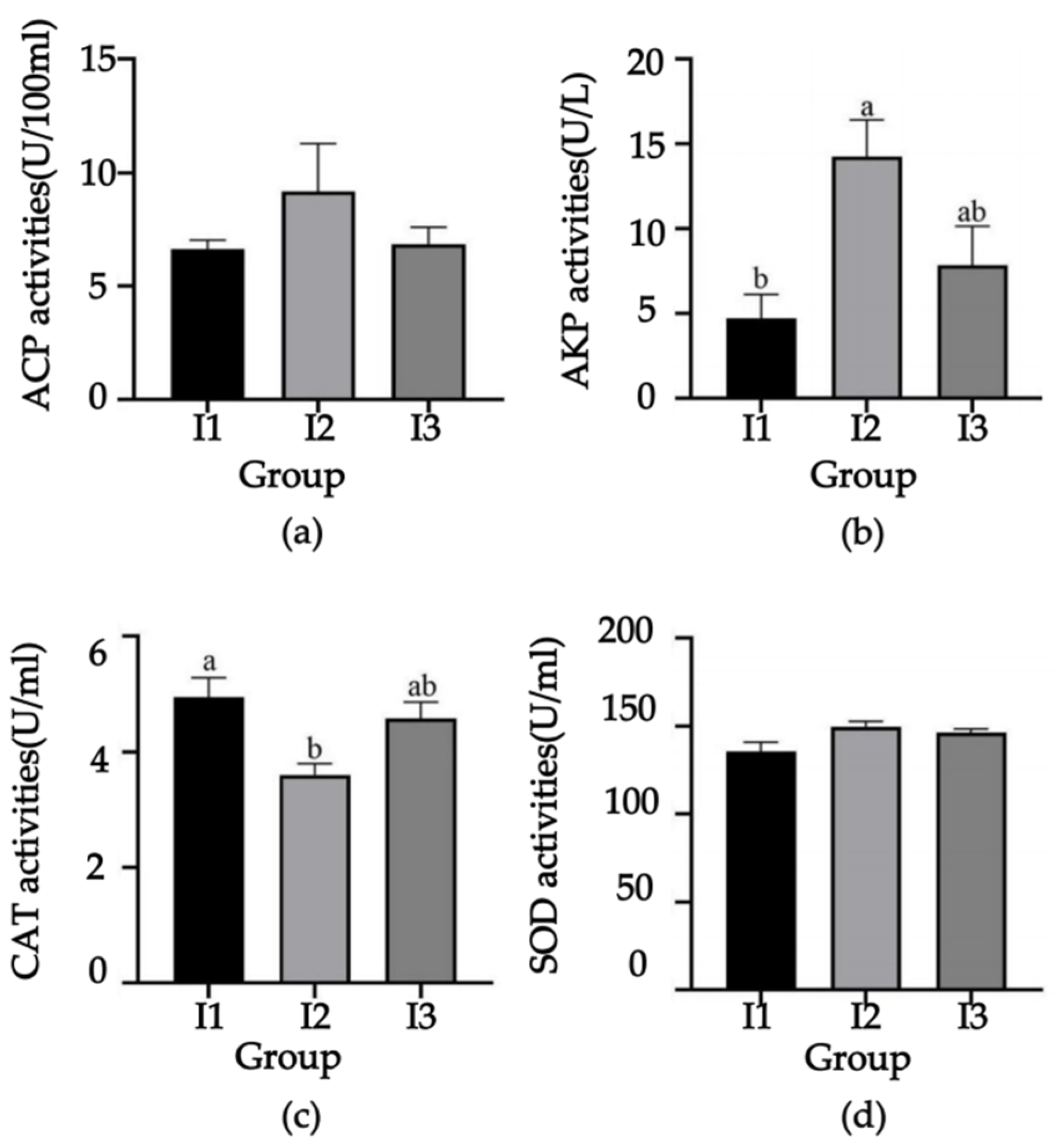
2.2. Serum Antioxidant Indicators
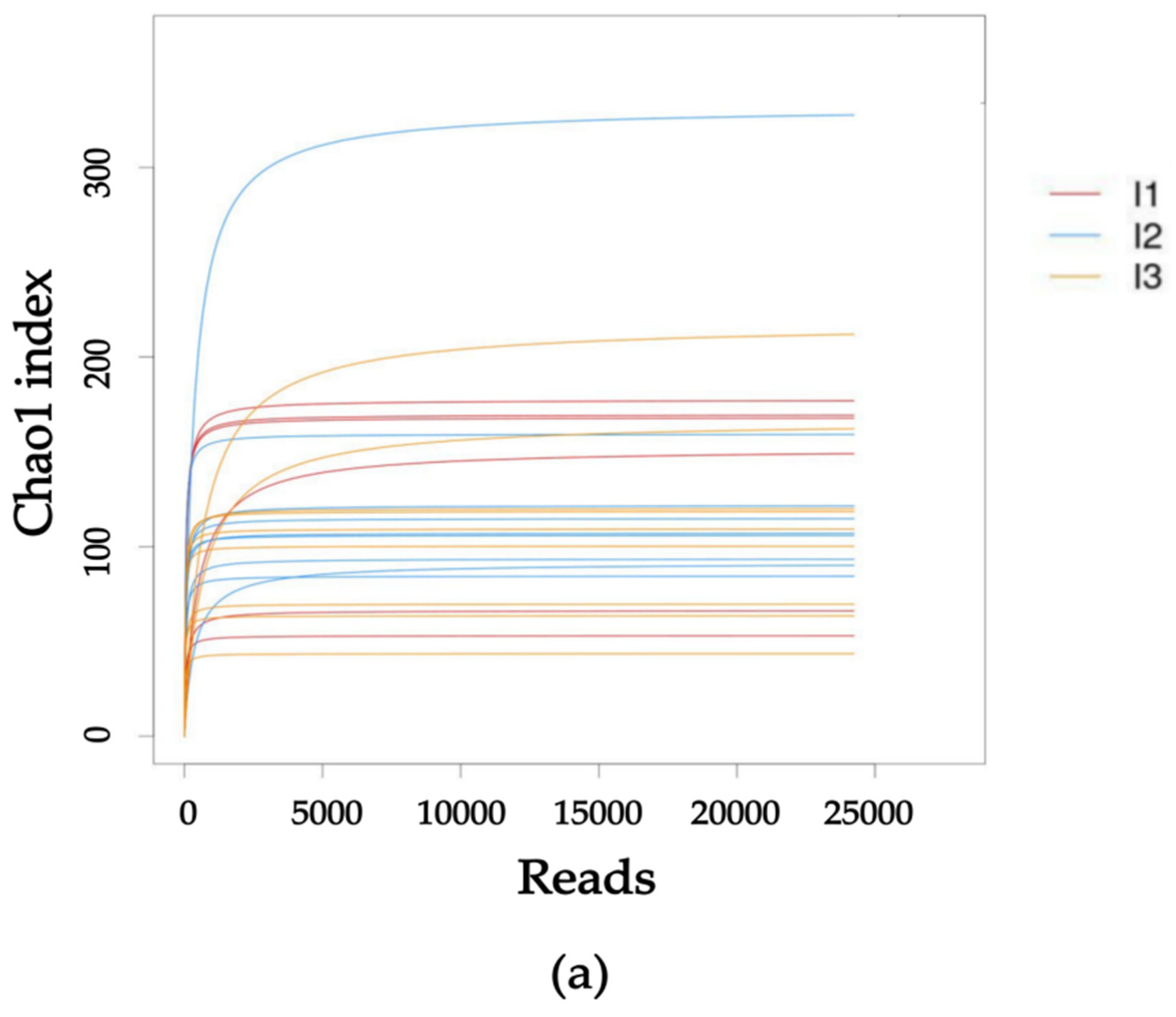
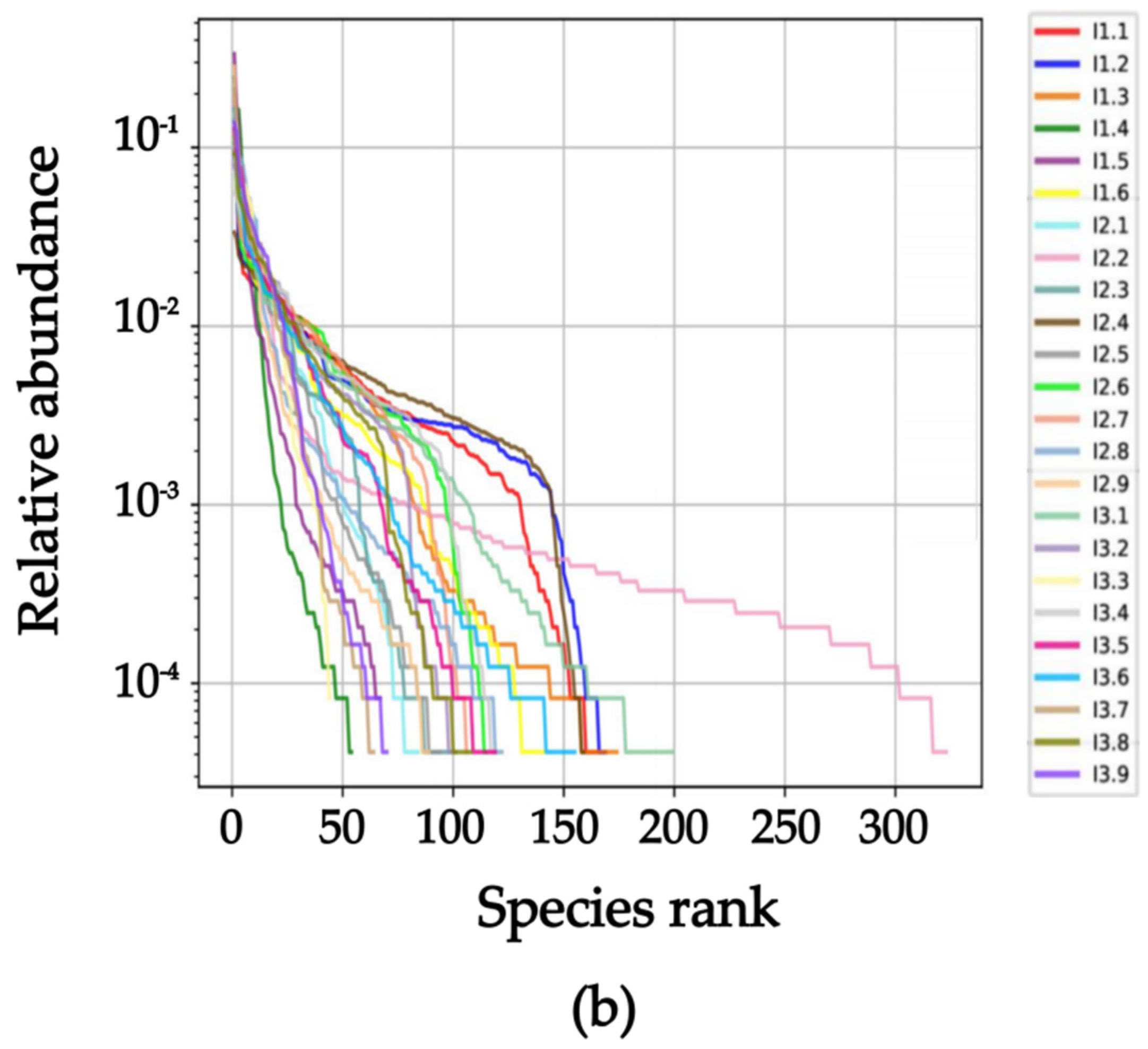
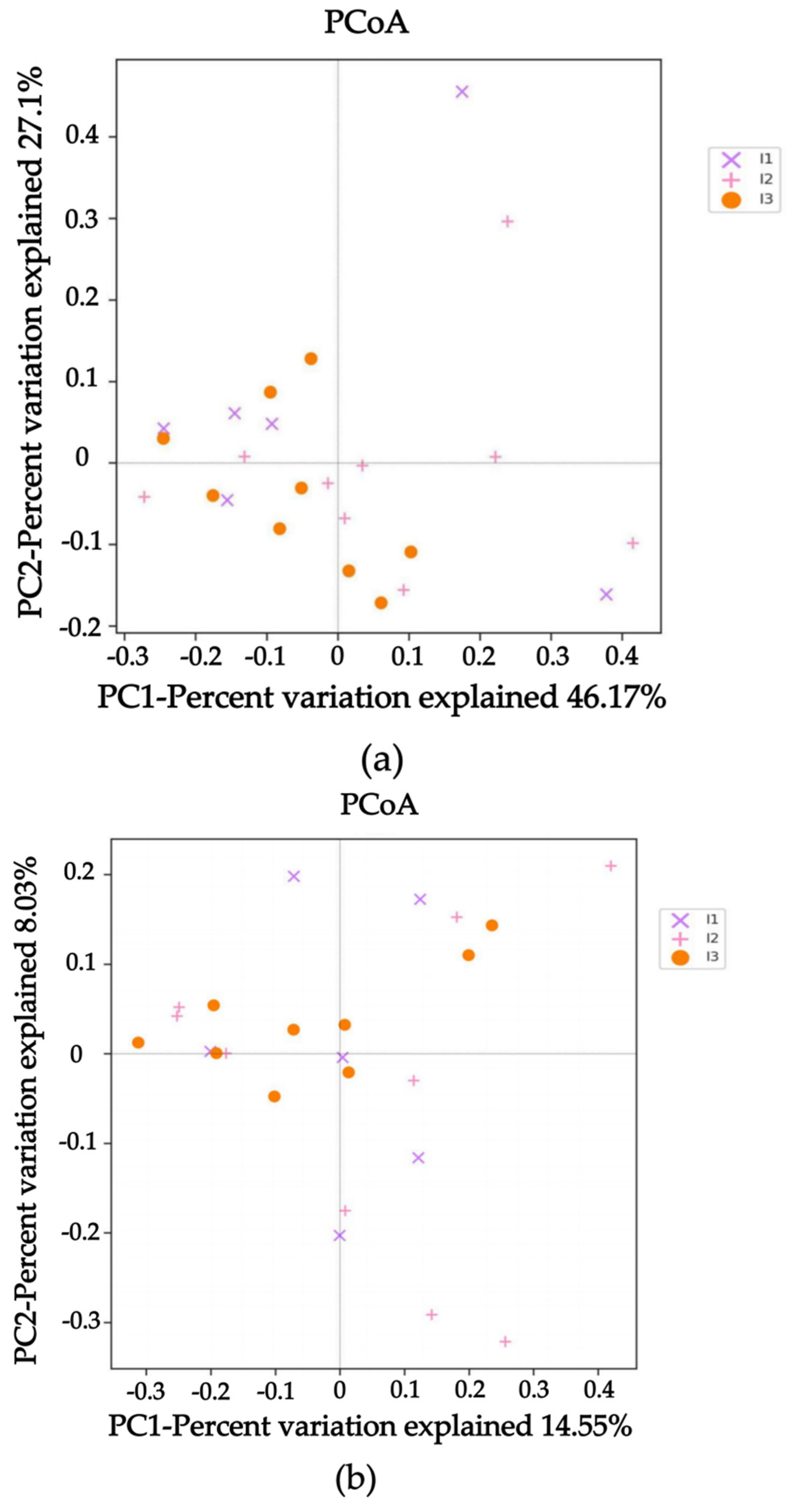
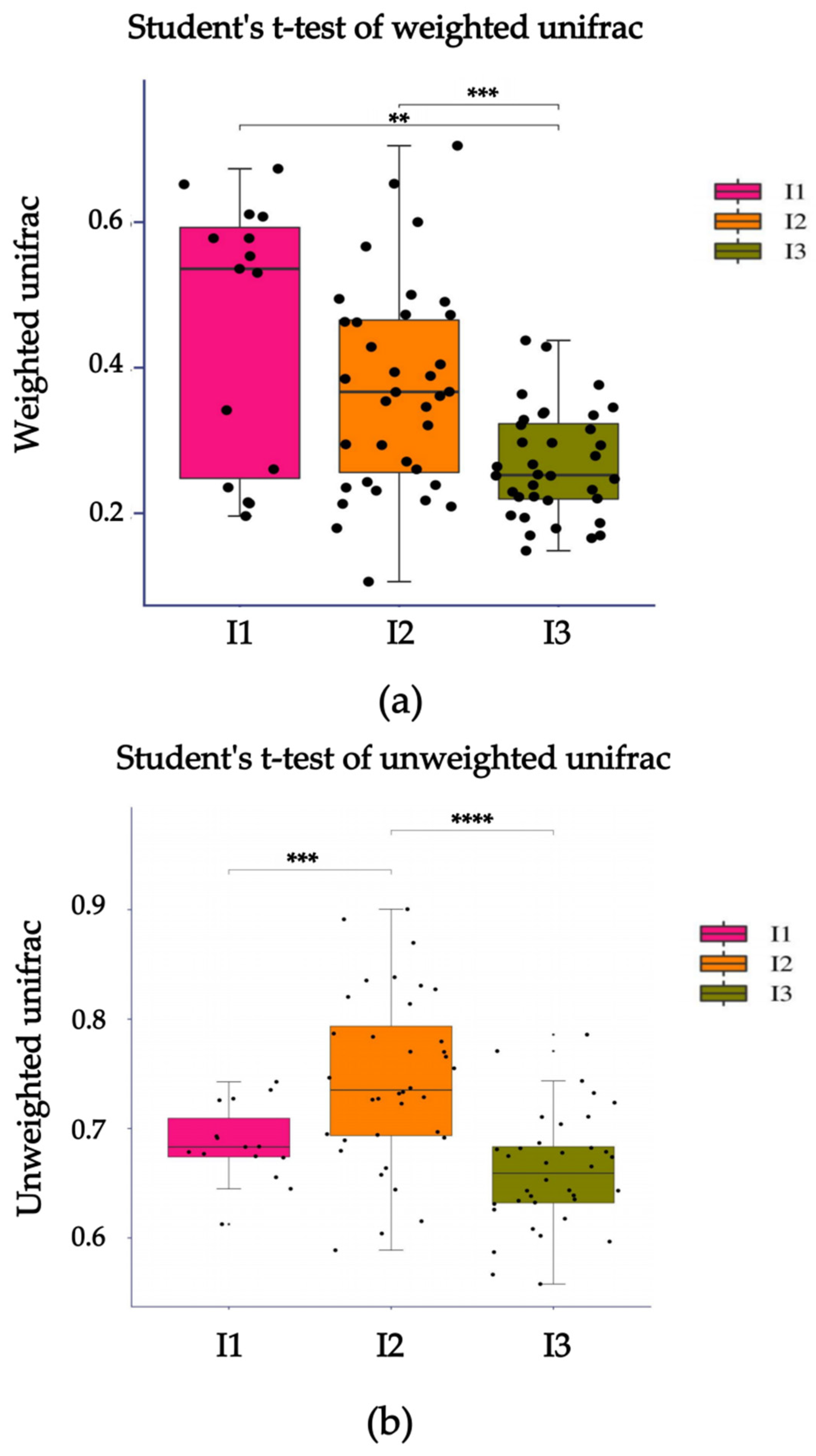
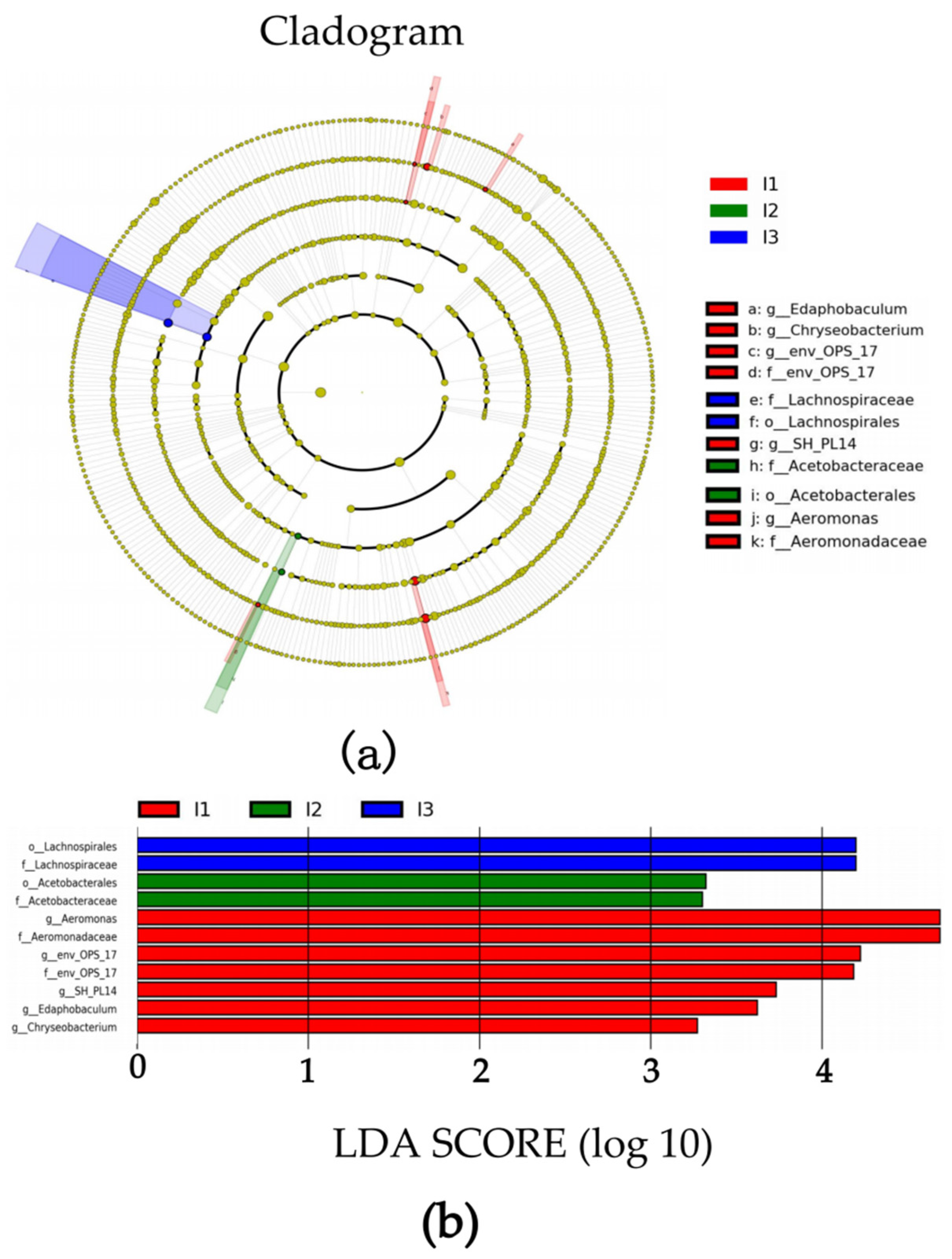
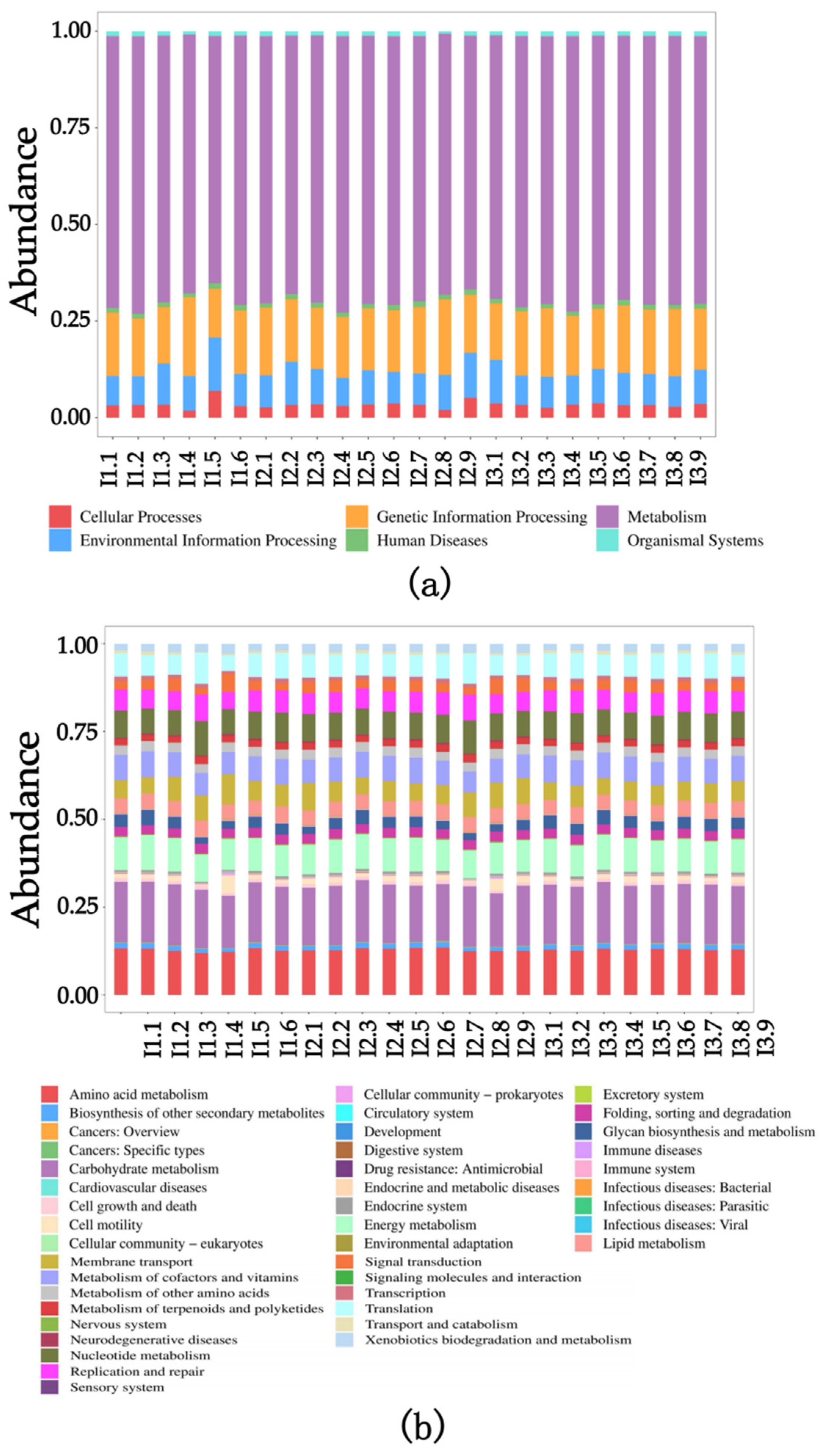
2.3. Intestinal Flora
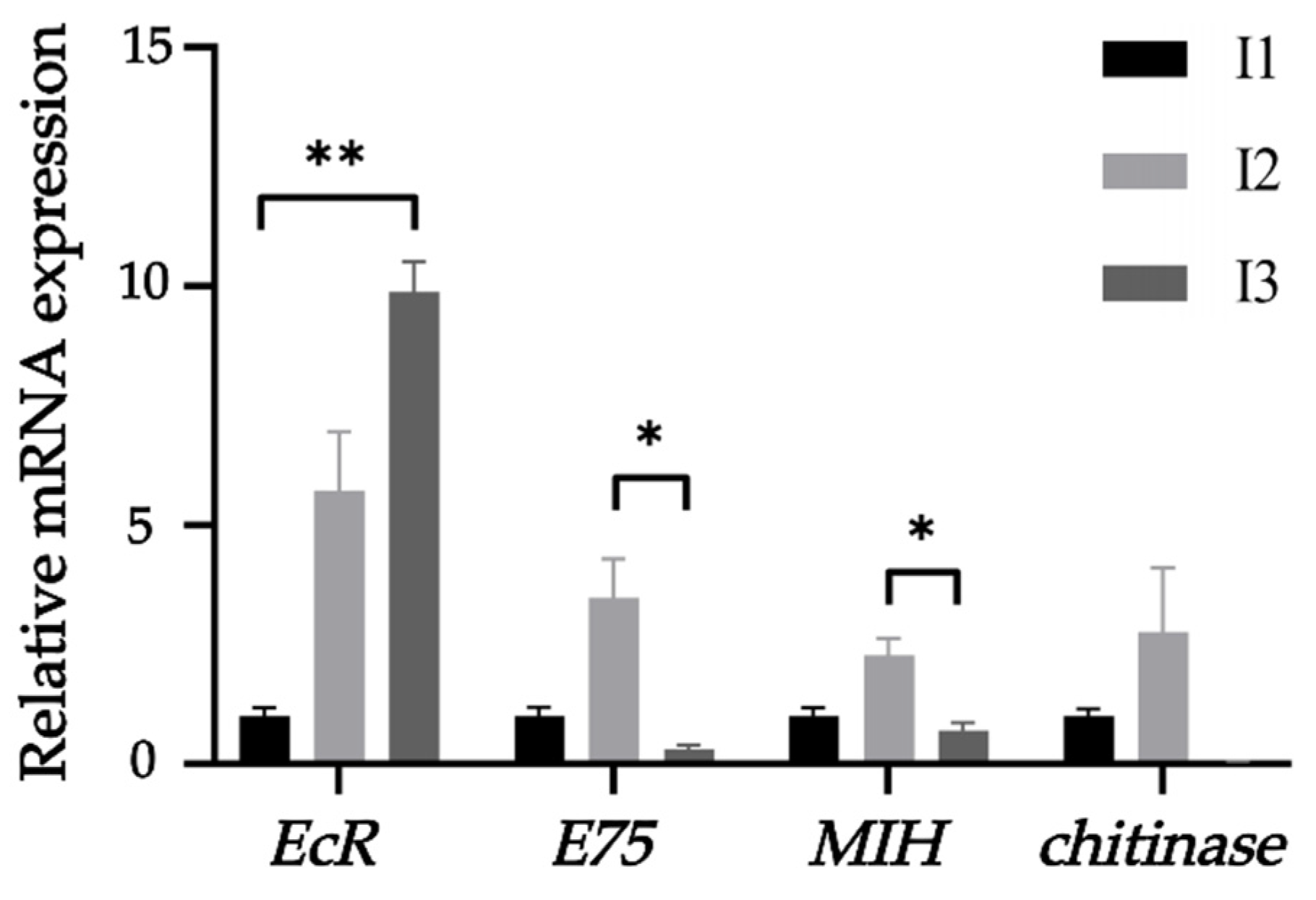
2.4. Growth-Related Gene Expression
2.5. Correlation Analysis of Growth and Immune Indexes, Growth Indicators and Growth-Related Gene Expression
3. Discussion
4. Materials and Methods
4.1. Culture Experiment
4.2. Sample Collection
4.3. Growth Index Determination
4.4. Determination of Antioxidant Indexes
4.5. Intestinal Flora Determination
4.6. Bioinformatic Analysis
4.7. Growth-Related Gene Determination
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Y.; Cao, C. Crayfish–Rice Integrated System of Production: An Agriculture Success Story in China. A Review. Agron. Sustain. Dev. 2021, 41, 68. [Google Scholar] [CrossRef]
- Altieri, M.A. Linking Ecologists and Traditional Farmers in the Search for Sustainable Agriculture. Front. Ecol. Environ. 2004, 2, 35–42. [Google Scholar] [CrossRef]
- Zhang, Z.; Deng, Q.; Cao, X.; Zhou, Y.; Song, C. Patterns of Sediment Fungal Community Dependent on Farming Practices in Aquaculture Ponds. Front. Microbiol. 2021, 12, 542064. [Google Scholar] [CrossRef]
- Zhang, Z.; Deng, Q.; Wan, L.; Cao, X.; Zhou, Y.; Song, C. Bacterial Communities and Enzymatic Activities in Sediments of Long-Term Fish and Crab Aquaculture Ponds. Microorganisms 2021, 9, 501. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.X.; Zheng, W.Z.; Lin, J.Y.; Shi, Y.; Xie, W.Z.; Zhou, H.M. Effect of Metal Ions on the Activity of Green Crab (Scylla serrata) Alkaline Phosphatase. Int. J. Biochem. Cell Biol. 2000, 32, 879–885. [Google Scholar] [CrossRef]
- Dong, J.; Cheng, R.; Yang, Y.; Zhao, Y.; Wu, G.; Zhang, R.; Zhu, X.; Li, L.; Li, X. Effects of Dietary Taurine on Growth, Non-Specific Immunity, Anti-Oxidative Properties and Gut Immunity in the Chinese Mitten Crab Eriocheir sinensis. Fish Shellfish Immunol. 2018, 82, 212–219. [Google Scholar] [CrossRef]
- Ou-Yang, K.; Feng, T.; Han, Y.; Li, J.; Ma, H. Cyhalofop-Butyl and Pyribenzoxim-Induced Oxidative Stress and Transcriptome Changes in the Muscle of Crayfish (Procambarus clarkii). Sci. Total Environ. 2023, 864, 161170. [Google Scholar] [CrossRef]
- Liu, F.; Geng, C.; Qu, Y.-K.; Cheng, B.-X.; Zhang, Y.; Wang, A.-M.; Zhang, J.-H.; Liu, B.; Tian, H.-Y.; Yang, W.-P.; et al. The Feeding of Dietary Codonopsis Pilosula Polysaccharide Enhances the Immune Responses, the Expression of Immune-Related Genes and the Growth Performance of Red Swamp Crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2020, 103, 321–331. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Prognon, P. Raw Material Enzymatic Activity Determination: A Specific Case for Validation and Comparison of Analytical Methods—The Example of Superoxide Dismutase (SOD). J. Pharm. Biomed. Anal. 2006, 40, 1143–1148. [Google Scholar] [CrossRef]
- Dai, C.; Xiao, L.; Mo, A.; Yuan, Y.; Yuan, J.; Gu, Z.; Wang, J. Effect of Dietary Bacillus subtilis Supplement on Cd Toxicokinetics and Cd-Induced Immune and Antioxidant Impairment of Procambarus clarkii. Environ. Sci. Pollut. Res. Int. 2023, 30, 43914–43926. [Google Scholar] [CrossRef]
- Yin, G.; Jeney, G.; Racz, T.; Xu, P.; Jun, X.; Jeney, Z. Effect of Two Chinese Herbs (Astragalus radix and Scutellaria radix) on Non-Specific Immune Response of Tilapia, Oreochromis niloticus. Aquaculture 2006, 253, 39–47. [Google Scholar] [CrossRef]
- Dai, L.-S.; Sun, Y.; Sun, Y.-X.; Zhu, B.-J.; Liu, C.-L. Characterization and Function of a Cathepsin B in Red Crayfish (Procambarus clarkii) Following Lipopolysaccharide Challenge. Fish Shellfish Immunol. 2016, 56, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Priya, T.A.J.; Li, F.; Zhang, J.; Yang, C.; Xiang, J. Molecular Characterization of an Ecdysone Inducible Gene E75 of Chinese Shrimp Fenneropenaeus Chinensis and Elucidation of Its Role in Molting by RNA Interference. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 156, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Lee, C.-Y.; Watson, R.D. Crustacean Molt-Inhibiting Hormone: Structure, Function, and Cellular Mode of Action. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 152, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, G.; Kumaresan, V.; Mahesh, A.; Dhayalan, A.; Arshad, A.; Arasu, M.V.; Al-Dhabi, N.A.; Pasupuleti, M.; Arockiaraj, J. Bactericidal and Fungistatic Activity of Peptide Derived from GH18 Domain of Prawn Chitinase 3 and Its Immunological Functions during Biological Stress. Int. J. Biol. Macromol. 2018, 106, 1014–1022. [Google Scholar] [CrossRef]
- Stevens, V.; Thijs, S.; Vangronsveld, J. Diversity and Plant Growth-Promoting Potential of (Un)Culturable Bacteria in the Hedera Helix Phylloplane. BMC Microbiol. 2021, 21, 66. [Google Scholar] [CrossRef]
- Sinkko, H.; Lukkari, K.; Sihvonen, L.M.; Sivonen, K.; Leivuori, M.; Rantanen, M.; Paulin, L.; Lyra, C. Bacteria Contribute to Sediment Nutrient Release and Reflect Progressed Eutrophication-Driven Hypoxia in an Organic-Rich Continental Sea. PLoS ONE 2013, 8, e67061. [Google Scholar] [CrossRef]
- Jia, B.; Li, Y.; Zi, X.; Gu, X.; Yuan, H.; Jeppesen, E.; Zeng, Q. Nutrient Enrichment Drives the Sediment Microbial Communities in Chinese Mitten Crab Eriocheir sinensis Culture. Environ. Res. 2023, 223, 115281. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, J.; Jin, X.; Liu, C.; Fan, C.; Guo, L.; Liang, Y.; Zheng, J.; Peng, N. Developmental, Dietary, and Geographical Impacts on Gut Microbiota of Red Swamp Crayfish (Procambarus clarkii). Microorganisms 2020, 8, 1376. [Google Scholar] [CrossRef]
- Zhu, X.; Ji, L.; Cheng, M.; Wei, H.; Wang, Z.; Ning, K. Sustainability of the Rice-Crayfish Co-Culture Aquaculture Model: Microbiome Profiles Based on Multi-Kingdom Analyses. Environ. Microbiome 2022, 17, 27. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of Diet on the Gut Microbiome and Implications for Human Health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Wang, Z.; Lam, K.-L.; Zeng, S.; Tan, B.K.; Hu, J. Role of Intestinal Microecology in the Regulation of Energy Metabolism by Dietary Polyphenols and Their Metabolites. Food Nutr. Res. 2019, 63, 1518. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Simpson, H.L.; Campbell, B.J. Review Article: Dietary Fibre-Microbiota Interactions. Aliment. Pharmacol. Ther. 2015, 42, 158–179. [Google Scholar] [CrossRef] [PubMed]
- Mazier, W.; Le Corf, K.; Martinez, C.; Tudela, H.; Kissi, D.; Kropp, C.; Coubard, C.; Soto, M.; Elustondo, F.; Rawadi, G.; et al. A New Strain of Christensenella Minuta as a Potential Biotherapy for Obesity and Associated Metabolic Diseases. Cells 2021, 10, 823. [Google Scholar] [CrossRef]
- Holt, C.C.; Bass, D.; Stentiford, G.D.; van der Giezen, M. Understanding the Role of the Shrimp Gut Microbiome in Health and Disease. J. Invertebr. Pathol. 2021, 186, 107387. [Google Scholar] [CrossRef]
- Chai, P.-C.; Song, X.-L.; Chen, G.-F.; Xu, H.; Huang, J. Dietary Supplementation of Probiotic Bacillus PC465 Isolated from the Gut of Fenneropenaeus chinensis Improves the Health Status and Resistance of Litopenaeus vannamei against White Spot Syndrome Virus. Fish Shellfish Immunol. 2016, 54, 602–611. [Google Scholar] [CrossRef]
- Martens, E.C.; Chiang, H.C.; Gordon, J.I. Mucosal Glycan Foraging Enhances Fitness and Transmission of a Saccharolytic Human Gut Bacterial Symbiont. Cell Host Microbe 2008, 4, 447–457. [Google Scholar] [CrossRef]
- Magnúsdóttir, S.; Heinken, A.; Kutt, L.; Ravcheev, D.A.; Bauer, E.; Noronha, A.; Greenhalgh, K.; Jäger, C.; Baginska, J.; Wilmes, P.; et al. Generation of Genome-Scale Metabolic Reconstructions for 773 Members of the Human Gut Microbiota. Nat. Biotechnol. 2017, 35, 81–89. [Google Scholar] [CrossRef]
- Tong, L.-C.; Wang, Y.; Wang, Z.-B.; Liu, W.-Y.; Sun, S.; Li, L.; Su, D.-F.; Zhang, L.-C. Propionate Ameliorates Dextran Sodium Sulfate-Induced Colitis by Improving Intestinal Barrier Function and Reducing Inflammation and Oxidative Stress. Front. Pharmacol. 2016, 7, 253. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Kholodkevich, S.; Sharov, A.; Chen, C.; Feng, Y.; Ren, N.; Sun, K. Effects of Cadmium on Intestinal Histology and Microbiota in Freshwater Crayfish (Procambarus clarkii). Chemosphere 2020, 242, 125105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, K.; Li, Z.; Chai, X.; Fu, X.; Kholodkevich, S.; Kuznetsova, T.; Chen, C.; Ren, N. Effescts of Acute Diclofenac Exposure on Intestinal Histology, Antioxidant Defense, and Microbiota in Freshwater Crayfish (Procambarus clarkii). Chemosphere 2021, 263, 128130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Z.; Tian, X.; Xu, P.; Sun, K.; Ren, N. Acute Toxic Effects of Microcystin-LR on Crayfish (Procambarus clarkii): Insights from Antioxidant System, Histopathology and Intestinal Flora. Environ. Sci. Pollut. Res. Int. 2023, 30, 56608–56619. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Li, H.; Ma, H.; Zhang, H.; Jin, L. Sensitive Acid Phosphatase Assay Based on Light-Activated Specific Oxidase Mimic Activity. Talanta 2023, 255, 124236. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, X.; Song, X.; Teng, P.; Ji, H.; Peng, L.; Qiu, Y.; Guo, D.; Jiang, S. Effect of Maduramicin on Crayfish (Procambius clarkii): Hematological Parameters, Oxidative Stress, Histopathological Changes and Stress Response. Ecotoxicol. Environ. Saf. 2021, 211, 111896. [Google Scholar] [CrossRef]
- Han, M.; Gao, T.; Liu, G.; Zhu, C.; Zhang, T.; Sun, M.; Li, J.; Ji, F.; Si, Q.; Jiang, Q. The Effect of a Polystyrene Nanoplastic on the Intestinal Microbes and Oxidative Stress Defense of the Freshwater Crayfish, Procambarus clarkii. Sci. Total Environ. 2022, 833, 155722. [Google Scholar] [CrossRef]
- Liu, F.; Qu, Y.-K.; Geng, C.; Wang, A.-M.; Zhang, J.-H.; Chen, K.-J.; Liu, B.; Tian, H.-Y.; Yang, W.-P.; Yu, Y.-B. Effects of Hesperidin on the Growth Performance, Antioxidant Capacity, Immune Responses and Disease Resistance of Red Swamp Crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2020, 99, 154–166. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-End Assembler for Illumina Sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality Control and Preprocessing of Metagenomic Datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Guo, F.; Wu, W.; Zhang, T. Characterization of Tetracycline Resistant Bacterial Community in Saline Activated Sludge Using Batch Stress Incubation with High-Throughput Sequencing Analysis. Water Res. 2013, 47, 4207–4216. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive Functional Profiling of Microbial Communities Using 16S RRNA Marker Gene Sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | I1 | I2 | I3 |
|---|---|---|---|
| Initial weight/g | 8.81 (8.13, 11.39) | 9.45 (8.73, 9.94) | 10.15 (9.58, 10.67) |
| Final weight/g | 14.27 (14.23, 14.33) | 15.71(14.53, 15.53) | 13.17 (12.13, 13.62) |
| WGR (%) | 63.03 a (51.00, 75.03) | 66.09 a (46.18, 77.89) | 29.14 b (18.92, 42.17) |
| Initial length/cm | 7.24 (7.06, 7.41) | 7.39 (7.12, 7.62) | 7.51 (7.36, 7.75) |
| Final length/cm | 8.37 (8.31, 8.41) | 8.59 (8.40, 8.89) | 8.26 (8.07, 8.41) |
| BLGR (%) | 15.68 a (12.15, 19.12) | 16.08 a (10.76, 19.65) | 9.86 b (6.71, 14.27) |
| SGR (%) | 0.97 a (0.82, 1.12) | 1.02 a (0.76, 1.15) | 0.51 b (0.35, 0.70) |
| SR (%) | 65.00 a (60.00, 80.00) | 73.33 a (60.00, 80.00) | 80.00 a (70.00, 90.00) |
| Sample | Chao1 | Shannon | Simpson |
|---|---|---|---|
| I1 | 133.67 ± 23.48 | 5.08 ± 0.57 | 0.93 ± 0.03 |
| I2 | 134.88 ± 24.73 | 5.16 ± 0.28 | 0.95 ± 0.01 |
| I3 | 112.13 ± 17.16 | 5.13 ± 0.22 | 0.95 ± 0.01 |
| Index | WGR | BLGR | SGR | SR | ACP | AKP | CAT | SOD |
|---|---|---|---|---|---|---|---|---|
| WGR | 1 | |||||||
| BLGR | 1.00 * | 1 | ||||||
| SGR | 1.00 ** | 1.00 * | 1 | |||||
| SR | −0.99 | −0.98 | −0.99 | 1 | ||||
| ACP | 0.50 | 0.48 | 0.51 | −0.64 | 1 | |||
| AKP | 0.27 | 0.25 | 0.28 | −0.42 | 0.97 | 1 | ||
| CAT | −0.33 | −0.31 | −0.34 | 0.48 | −0.98 | −1.00 * | 1 | |
| SOD | −0.23 | −0.24 | −0.21 | 0.06 | 0.73 | 0.88 | −0.85 | 1 |
| Index | WGR | BLGR | SGR | SR | EcR | E75 | MIH | Chitinase |
|---|---|---|---|---|---|---|---|---|
| WGR | 1 | |||||||
| BLGR | 1.00 * | 1 | ||||||
| SGR | 1.00 ** | 1.00 * | 1 | |||||
| SR | −0.99 | −0.98 | −0.99 | 1 | ||||
| EcR | −0.81 | −0.82 | −0.80 | 0.69 | 1 | |||
| E75 | 0.72 | 0.71 | 0.73 | −0.83 | −0.17 | 1 | ||
| MIH | 0.70 | 0.69 | 0.71 | −0.81 | −0.14 | 1.00 * | 1 | |
| Chitinase | 0.82 | 0.81 | 0.83 | −0.90 | −0.32 | 0.99 | 0.98 | 1 |
| Primer | Sequence (5′-3′) | Usage |
|---|---|---|
| AK | F: TCCTCGACGTAATCCAGTCC | expression of AK |
| R: CGAAGTCCTTGTTGGGATGT | ||
| EcR | F: GCTCGGACGCAGAGATTCAA | expression of EcR |
| R: GAAAGTTTTCGCCGCCGATG | ||
| E75 | F: TGTCTACGACGCCATTAGGC | expression of E75 |
| R: CGAATCTGCGATGTCCACCT | ||
| MIH | F: CTCCCAAGATCACAGCGTCA | expression of MIH |
| R: CAGTTCAAGGTCGAGTCCCA | ||
| Chitinase | F: GTACGATCTGCGAGGCAACT | expression of chitinase |
| R: CAACACCAGTTTGTCAGCGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, R.; Zhang, C.; Tang, Y.; Zhu, J.; Yang, N.; Su, S. Effects of Different Sources of Culture Substrate on the Growth and Immune Performance of the Red Swamp Crayfish (Procambarus clarkii). Int. J. Mol. Sci. 2023, 24, 14098. https://doi.org/10.3390/ijms241814098
Wan R, Zhang C, Tang Y, Zhu J, Yang N, Su S. Effects of Different Sources of Culture Substrate on the Growth and Immune Performance of the Red Swamp Crayfish (Procambarus clarkii). International Journal of Molecular Sciences. 2023; 24(18):14098. https://doi.org/10.3390/ijms241814098
Chicago/Turabian StyleWan, Rong, Chengfeng Zhang, Yongkai Tang, Jian Zhu, Nan Yang, and Shengyan Su. 2023. "Effects of Different Sources of Culture Substrate on the Growth and Immune Performance of the Red Swamp Crayfish (Procambarus clarkii)" International Journal of Molecular Sciences 24, no. 18: 14098. https://doi.org/10.3390/ijms241814098