

Vascular Endothelial Growth Factor A (VEGFA) Regulates Hepatic Lipid and Glycogen Metabolism in Schizothorax prenanti

Abstract
:1. Introduction
2. Results
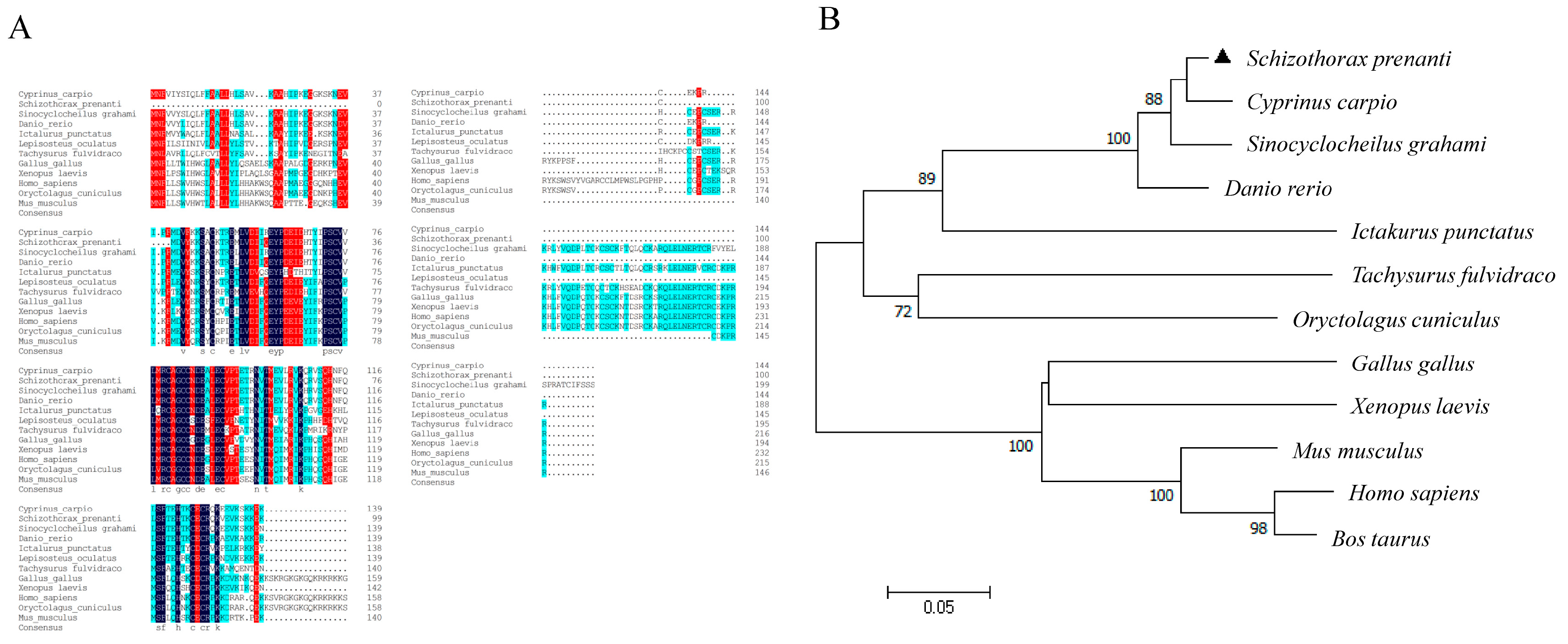
2.1. Molecular Characterization and Tissue Distribution of VEGFA Gene in S. prenanti
2.2. Production of Recombinant S. prenanti VEGFA Protein
2.3. Biochemical Analysis of Plasma in S. prenanti after Intraperitoneal Injection of VEGFA Protein
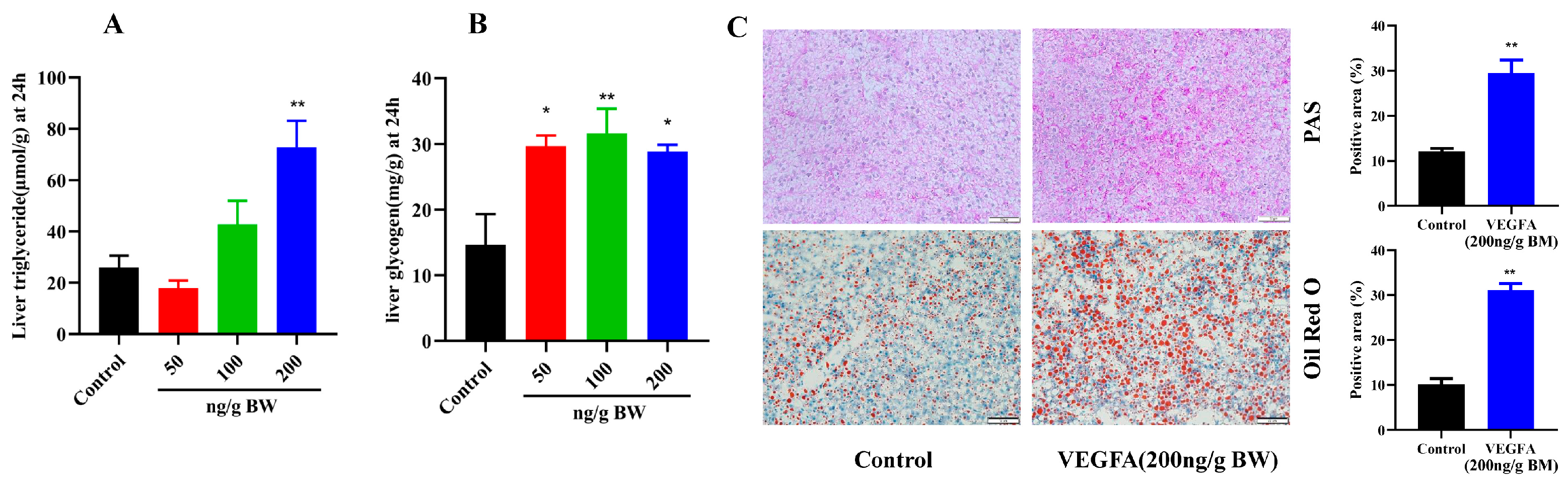
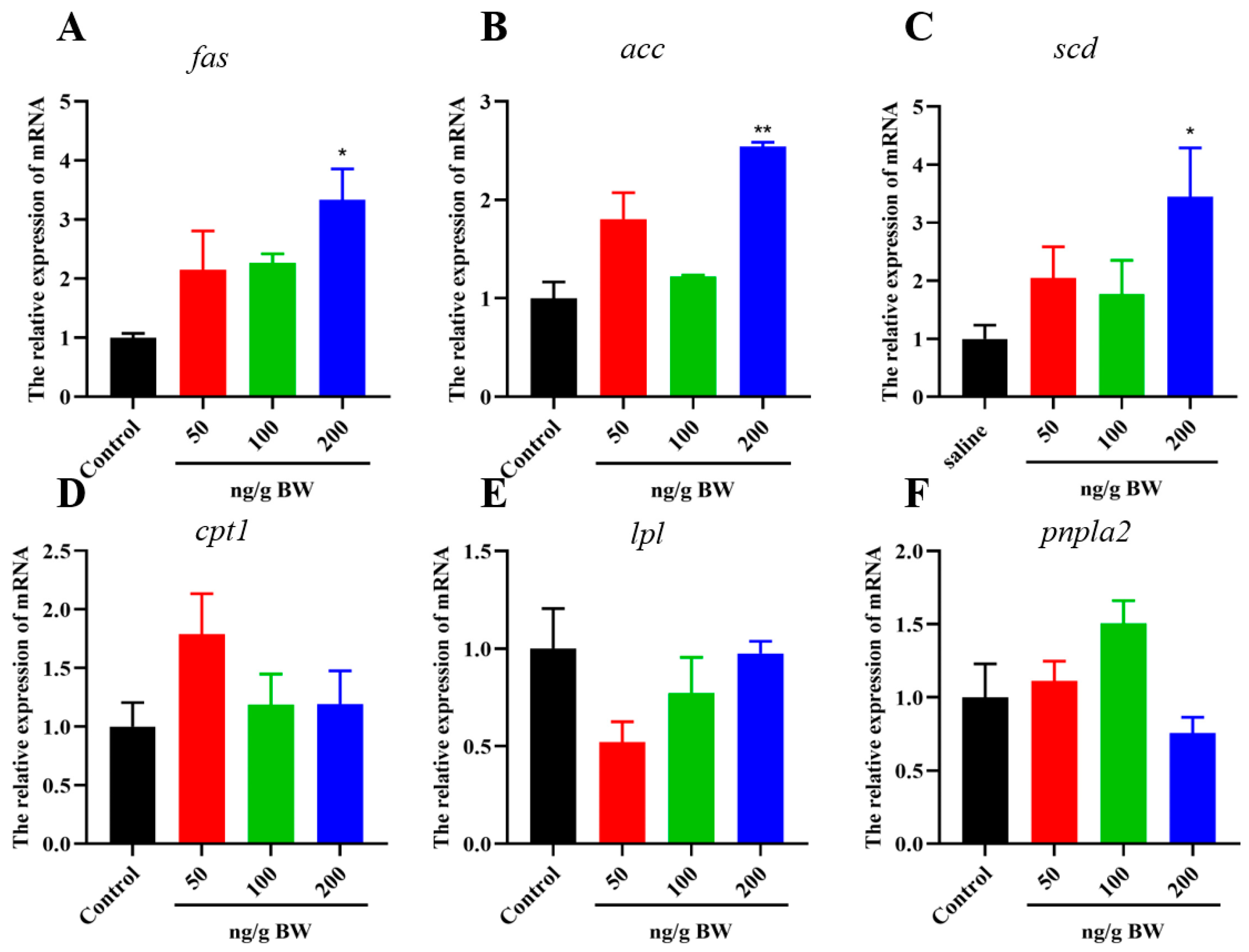
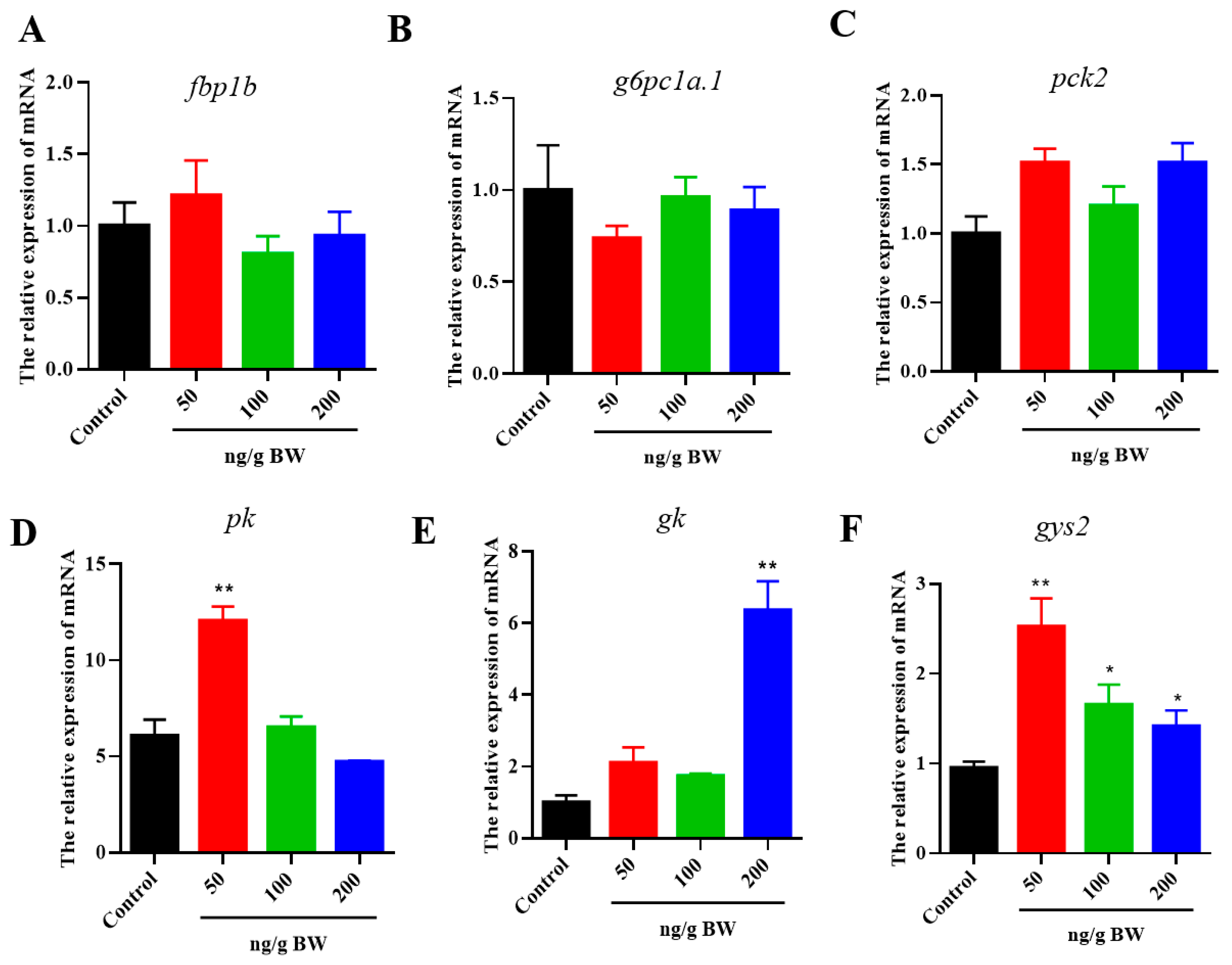
2.4. Effects of VEGFA Protein on Hepatic Lipid and Glycogen Metabolism in S. prenanti
2.5. Effects of VEGFA Protein on Lipid and Glycogen Metabolism in Hepatocytes
2.6. VEGFA Regulates Glycolipid Metabolism through AMPKα Signaling in Hepatocytes
3. Discussion
4. Materials and Methods
4.1. Animals and Intraperitoneal Injection of VEGFA Recombinant Protein
4.2. Blood Biochemical Assays
4.3. Oil Red O and PAS Staining
4.4. Triglyceride and Glycogen Content Measurement
4.5. Cloning of S. prenanti VEGFA and Sequence Analysis
4.6. Expression of VEGFA Recombinant Protein and Purification
4.7. Primary Hepatocytes Culture and Recombinant VEGFA Protein Treatment
4.8. qPCR
4.9. Western Blotting
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Q.; Du, J.; Cui, K.; Fang, W.; Zhao, Z.; Chen, Q.; Mai, K.; Ai, Q. Acetyl-CoA derived from hepatic mitochondrial fatty acid beta-oxidation aggravates inflammation by enhancing p65 acetylation. iScience 2021, 24, 103244. [Google Scholar] [CrossRef] [PubMed]
- Naiel, M.A.E.; Negm, S.S.; Ghazanfar, S.; Shukry, M.; Abdelnour, S.A. The risk assessment of high-fat diet in farmed fish and its mitigation approaches: A review. J. Anim. Physiol. Anim. Nutr. 2023, 107, 948–969. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Liao, K.; Wang, T.J.; Mai, K.S.; Xu, W.; Ai, Q.H. Dietary Lipid Levels Influence Lipid Deposition in the Liver of Large Yellow Croaker (Larimichthys crocea) by Regulating Lipoprotein Receptors, Fatty Acid Uptake and Triacylglycerol Synthesis and Catabolism at the Transcriptional Level. PLoS ONE 2015, 10, e0129937. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Castro, F.; Monroig, O.; Cao, X.; Gao, J. fat-1 transgenic zebrafish are protected from abnormal lipid deposition induced by high-vegetable oil feeding. Appl. Microbiol. Biotechnol. 2020, 104, 7355–7365. [Google Scholar] [CrossRef]
- Jia, R.; Cao, L.P.; Du, J.L.; He, Q.; Gu, Z.Y.; Jeney, G.; Xu, P.; Yin, G.J. Effects of high-fat diet on antioxidative status, apoptosis and inflammation in liver of tilapia (Oreochromis niloticus) via Nrf2, TLRs and JNK pathways. Fish. Shellfish. Immunol. 2020, 104, 391–401. [Google Scholar] [CrossRef]
- Favalier, N.; Veron, V.; Marchand, M.; Surget, A.; Maunas, P.; Turonnet, N.; Panserat, S.; Marandel, L. Short-Term Effect of a Low-Protein High-Carbohydrate Diet on Mature Female and Male, and Neomale Rainbow Trout. Int. J. Mol. Sci. 2021, 22, 6149. [Google Scholar] [CrossRef]
- Morris, S.; Cholan, P.M.; Britton, W.J.; Oehlers, S.H. Glucose inhibits haemostasis and accelerates diet-induced hyperlipidaemia in zebrafish larvae. Sci. Rep. 2021, 11, 19049. [Google Scholar] [CrossRef]
- Ning, L.; Zhang, H.; Chen, X.; Zhen, J.; Chen, S.; Guang, J.; Xu, C.; Li, Y. A comparative study on the tolerance of tilapia (Oreochromis niloticus) to high carbohydrate and high lipid diets. Anim. Nutr. 2023, 13, 160–172. [Google Scholar] [CrossRef]
- Li, X.; Zheng, S.; Ma, X.; Cheng, K.; Wu, G. Effects of dietary starch and lipid levels on the protein retention and growth of largemouth bass (Micropterus salmoides). Amino Acids 2020, 52, 999–1016. [Google Scholar] [CrossRef]
- Han, H.S.; Kang, G.; Kim, J.S.; Choi, B.H.; Koo, S.H. Regulation of glucose metabolism from a liver-centric perspective. Exp. Mol. Med. 2016, 48, e218. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Toselli, C.M.; Wilkinson, B.M.; Paterson, J.; Kieffer, T.J. Vegfa/vegfr2 signaling is necessary for zebrafish islet vessel development, but is dispensable for beta-cell and alpha-cell formation. Sci. Rep. 2019, 9, 3594. [Google Scholar] [CrossRef] [PubMed]
- Jabs, N.; Franklin, I.; Brenner, M.B.; Gromada, J.; Ferrara, N.; Wollheim, C.B.; Lammert, E. Reduced insulin secretion and content in VEGF-a deficient mouse pancreatic islets. Exp. Clin. Endocrinol. Diabetes 2008, 116 (Suppl. 1), S46–S49. [Google Scholar] [CrossRef] [PubMed]
- Reinert, R.B.; Cai, Q.; Hong, J.Y.; Plank, J.L.; Aamodt, K.; Prasad, N.; Aramandla, R.; Dai, C.; Levy, S.E.; Pozzi, A.; et al. Vascular endothelial growth factor coordinates islet innervation via vascular scaffolding. Development 2014, 141, 1480–1491. [Google Scholar] [CrossRef]
- Sellami, N.; Lamine, L.B.; Turki, A.; Sarray, S.; Jailani, M.; Al-Ansari, A.K.; Ghorbel, M.; Mahjoub, T.; Almawi, W.Y. Association of VEGFA variants with altered VEGF secretion and type 2 diabetes: A case-control study. Cytokine 2018, 106, 29–34. [Google Scholar] [CrossRef]
- Huang, L.O.; Rauch, A.; Mazzaferro, E.; Preuss, M.; Carobbio, S.; Bayrak, C.S.; Chami, N.; Wang, Z.; Schick, U.M.; Yang, N.; et al. Genome-wide discovery of genetic loci that uncouple excess adiposity from its comorbidities. Nat. Metab. 2021, 3, 228–243. [Google Scholar] [CrossRef]
- Sung, H.K.; Doh, K.O.; Son, J.E.; Park, J.G.; Bae, Y.; Choi, S.; Nelson, S.M.L.; Cowling, R.; Nagy, K.; Michael, I.P.; et al. Adipose Vascular Endothelial Growth Factor Regulates Metabolic Homeostasis through Angiogenesis. Cell Metab. 2013, 17, 61–72. [Google Scholar] [CrossRef]
- Elias, I.; Franckhauser, S.; Bosch, F. New insights into adipose tissue VEGF-A actions in the control of obesity and insulin resistance. Adipocyte 2013, 2, 109–112. [Google Scholar] [CrossRef]
- Elias, I.; Franckhauser, S.; Ferré, T.; Vilà, L.; Tafuro, S.; Muñoz, S.; Roca, C.; Ramos, D.; Pujol, A.; Riu, E.; et al. Adipose tissue overexpression of vascular endothelial growth factor protects against diet-induced obesity and insulin resistance. Diabetes 2012, 61, 1801–1813. [Google Scholar] [CrossRef]
- Brissova, M.; Shostak, A.; Shiota, M.; Wiebe, P.O.; Poffenberger, G.; Kantz, J.; Chen, Z.; Carr, C.; Jerome, W.G.; Chen, J.; et al. Pancreatic islet production of vascular endothelial growth factor—A is essential for islet vascularization, revascularization, and function. Diabetes 2006, 55, 2974–2985. [Google Scholar] [CrossRef]
- Ji, R.; Xu, X.; Turchini, G.M.; Mai, K.; Ai, Q. Adiponectin’s roles in lipid and glucose metabolism modulation in fish: Mechanisms and perspectives. Rev. Aquac. 2021, 13, 2305–2321. [Google Scholar] [CrossRef]
- Kim, K.-D.; Kim, K.-M.; Kim, K.-W.; Kang, Y.J.; Lee, S.-M. Influence of lipid level and supplemental lecithin in diet on growth, feed utilization and body composition of juvenile flounder (Paralichthys olivaceus) in suboptimal water temperatures. Aquaculture 2006, 251, 484–490. [Google Scholar] [CrossRef]
- Cao, X.-F.; Dai, Y.-J.; Liu, M.-Y.; Yuan, X.-Y.; Wang, C.-C.; Huang, Y.-Y.; Liu, W.-B.; Jiang, G.-Z. High-fat diet induces aberrant hepatic lipid secretion in blunt snout bream by activating endoplasmic reticulum stress-associated IRE1/XBP1 pathway. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, R.; Yi, J.; Guo, W.; Cheng, Q.; Zhi, L.; Lin, Y. Characterization and expression profiles of muscle transcriptome in Schizothoracine fish, Schizothorax prenanti. Gene 2019, 685, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, J.; Dong, L.; Huang, A.; Bu, G.; Meng, F.; Kong, F.; Cao, X.; Han, X.; Pan, X.J.F.; et al. Teleost-specific TLR25 identified from Schizothorax prenanti may recognize bacterial/viral components and activate NF-κB and type I IFNs signaling pathways. Fish Shellfish. Immunol. 2018, 82, 361–370. [Google Scholar] [CrossRef]
- Xin, H.; Zhong, C.; Nudleman, E.; Ferrara, N. Evidence for Pro-angiogenic Functions of VEGF-Ax. Cell 2016, 167, 275–284.e6. [Google Scholar] [CrossRef]
- Habeck, H.; Odenthal, J.; Walderich, B.; Maischein, H.; Schulte-Merker, S.; Tubingen screen, c. —Analysis of a zebrafish VEGF receptor mutant reveals specific disruption of angiogenesis. Curr. Biol. 2002, 12, 1405–1412. [Google Scholar] [CrossRef]
- Luo, W.; Liang, X.; Huang, S.; Cao, X. Molecular cloning, expression analysis and miRNA prediction of vascular endothelial growth factor A (VEGFAa and VEGFAb) in pond loach Misgurnus anguillicaudatus, an air-breathing fish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 202, 39–47. [Google Scholar] [CrossRef]
- Bahary, N.; Goishi, K.; Stuckenholz, C.; Weber, G.; LeBlanc, J.; Schafer, C.A.; Berman, S.S.; Klagsbrun, M.; Zon, L.I. Duplicate VegfA genes and orthologues of the KDR receptor tyrosine kinase family mediate vascular development in the zebrafish. Blood 2007, 110, 3627–3636. [Google Scholar] [CrossRef]
- Gong, B.; Liang, D.; Chew, T.G.; Ge, R. Characterization of the zebrafish vascular endothelial growth factor A gene: Comparison with vegf-A genes in mammals and Fugu. Biochim. Biophys. Acta 2004, 1676, 33–40. [Google Scholar] [CrossRef]
- Rissone, A.; Foglia, E.; Sangiorgio, L.; Cermenati, S.; Nicoli, S.; Cimbro, S.; Beltrame, M.; Bussolino, F.; Cotelli, F.; Arese, M. The synaptic proteins β-neurexin and neuroligin synergize with extracellular matrix-binding vascular endothelial growth factor a during zebrafish vascular development. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Fang, Y.; Gao, K.; Shen, J.; Zhong, T.P.; Li, F. Vegfa Impacts Early Myocardium Development in Zebrafish. Int. J. Mol. Sci. 2017, 18, 444. [Google Scholar] [CrossRef] [PubMed]
- Chapouly, C.; Argaw, A.T.; Horng, S.; Castro, K.; Zhang, J.Y.; Asp, L.; Loo, H.; Laitman, B.M.; Mariani, J.N.; Farber, R.S.; et al. Astrocytic TYMP and VEGFA drive blood-brain barrier opening in inflammatory central nervous system lesions. Brain 2015, 138, 1548–1567. [Google Scholar] [CrossRef] [PubMed]
- Bujaldon, E.; Cornide-Petronio, M.E.; Gulfo, J.; Rotondo, F.; Avalos de Leon, C.; Negrete-Sanchez, E.; Gracia-Sancho, J.; Novials, A.; Jimenez-Castro, M.B.; Peralta Uroz, C. Relevance of VEGFA in rat livers subjected to partial hepatectomy under ischemia-reperfusion. J. Mol. Med. 2019, 97, 1299–1314. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Zhu, D.; Fang, Y.; Chen, Y.; Yu, G.; Pan, W.; Liu, D.; Li, F.; Zhong, T.P. Vegfa signaling regulates diverse artery/vein formation in vertebrate vasculatures. J. Genet. Genom. 2017, 44, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Aghajanian, A.; Zhang, H.; Buckley, B.K.; Wittchen, E.S.; Ma, W.Y.; Faber, J.E. Decreased inspired oxygen stimulates de novo formation of coronary collaterals in adult heart. J. Mol. Cell Cardiol. 2021, 150, 1–11. [Google Scholar] [CrossRef]
- Bagchi, M.; Kim, L.A.; Boucher, J.; Walshe, T.E.; Kahn, C.R.; D’Amore, P.A. Vascular endothelial growth factor is important for brown adipose tissue development and maintenance. FASEB J. 2013, 27, 3257–3271. [Google Scholar] [CrossRef]
- Sun, K.; Wernstedt Asterholm, I.; Kusminski, C.M.; Bueno, A.C.; Wang, Z.V.; Pollard, J.W.; Brekken, R.A.; Scherer, P.E. Dichotomous effects of VEGF-A on adipose tissue dysfunction. Proc. Natl. Acad. Sci. USA 2012, 109, 5874–5879. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, X.; Yang, L.; Eckel-Mahan, K.; Tong, Q.; Gu, X.; Kolonin, M.G.; Sun, K. Transient Overexpression of Vascular Endothelial Growth Factor A in Adipose Tissue Promotes Energy Expenditure via Activation of the Sympathetic Nervous System. Mol. Cell Biol. 2018, 38, e00242-18. [Google Scholar] [CrossRef]
- Sun, K.; Kusminski, C.M.; Luby-Phelps, K.; Spurgin, S.B.; An, Y.A.; Wang, Q.A.; Holland, W.L.; Scherer, P.E. Brown adipose tissue derived VEGF-A modulates cold tolerance and energy expenditure. Mol. Metab. 2014, 3, 474–483. [Google Scholar] [CrossRef]
- Wu, L.E.; Meoli, C.C.; Mangiafico, S.P.; Fazakerley, D.J.; Cogger, V.C.; Mohamad, M.; Pant, H.; Kang, M.J.; Powter, E.; Burchfield, J.G.; et al. Systemic VEGF-A neutralization ameliorates diet-induced metabolic dysfunction. Diabetes 2014, 63, 2656–2667. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, M.; Sun, K.; An, Y.A.; Gu, X.; Scherer, P.E. VEGF-A-Expressing Adipose Tissue Shows Rapid Beiging and Enhanced Survival After Transplantation and Confers IL-4-Independent Metabolic Improvements. Diabetes 2017, 66, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Giurdanella, G.; Lupo, G.; Gennuso, F.; Conti, F.; Furno, D.L.; Mannino, G.; Anfuso, C.D.; Drago, F.; Salomone, S.; Bucolo, C. Activation of the VEGF-A/ERK/PLA2 Axis Mediates Early Retinal Endothelial Cell Damage Induced by High Glucose: New Insight from an In Vitro Model of Diabetic Retinopathy. Int. J. Mol. Sci. 2020, 21, 7528. [Google Scholar] [CrossRef] [PubMed]
- Ayalasomayajula, S.P.; Kompella, U.B. Celecoxib, a selective cyclooxygenase-2 inhibitor, inhibits retinal vascular endothelial growth factor expression and vascular leakage in a streptozotocin-induced diabetic rat model. Eur. J. Pharmacol. 2003, 458, 283–289. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef]
- Esquejo, R.M.; Albuquerque, B.; Sher, A.; Blatnik, M.; Wald, K.; Peloquin, M.; Delmore, J.; Kindt, E.; Li, W.; Young, J.D.; et al. AMPK activation is sufficient to increase skeletal muscle glucose uptake and glycogen synthesis but is not required for contraction-mediated increases in glucose metabolism. Heliyon 2022, 8, e11091. [Google Scholar] [CrossRef]
- Zhang, C.; Zhong, T.; Li, Y.; Li, X.; Yuan, X.; Liu, L.; Wu, W.; Wu, J.; Wu, Y.; Liang, R.; et al. The hepatic AMPK-TET1-SIRT1 axis regulates glucose homeostasis. Elife 2021, 10, e70672. [Google Scholar] [CrossRef]
- Rindone, G.M.; Dasso, M.E.; Centola, C.L.; Pellizzari, E.H.; Camberos, M.D.C.; Toneatto, J.; Galardo, M.N.; Meroni, S.B.; Riera, M.F. Sertoli cell adaptation to glucose deprivation: Potential role of AMPK in the regulation of lipid metabolism. J. Cell Biochem. 2023, 124, 716–730. [Google Scholar] [CrossRef]
- Wang, L.; Liu, X.; Zhan, S.; Guo, J.; Yang, S.; Zhong, T.; Li, L.; Zhang, H.; Wang, Y. Inhibition of GSK3beta Reduces Ectopic Lipid Accumulation and Induces Autophagy by the AMPK Pathway in Goat Muscle Satellite Cells. Cells 2019, 8, 1378. [Google Scholar] [CrossRef]
- Hemre, G.I.; Mommsen, T.P.; Krogdahl, Å. Carbohydrates in fish nutrition: Effects on growth, glucose metabolism and hepatic enzymes. Aquac. Nutr. 2002, 8, 175–194. [Google Scholar] [CrossRef]
- Alexander, C.; Sahu, N.; Pal, A.; Akhtar, M. Haemato-immunological and stress responses of Labeo rohita (Hamilton) fingerlings: Effect of rearing temperature and dietary gelatinized carbohydrate. J. Anim. Physiol. Anim. Nutr. 2011, 95, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, P.; Ni, J.; Mo, Q.; Luo, W.; Du, Z.; Jiang, J.; Yang, S.; Zhao, L.; Gong, Q. Molecular and functional characterization of the retinol-binding protein 4 (RBP4) in hepatocytes of Schizothorax prenanti in response to palmitic acid. Fish. Physiol. Biochem. 2022, 48, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Zhou, T.; Wang, Y.; Wang, L. Cold Exposure Regulates Hepatic Glycogen and Lipid Metabolism in Newborn Goats. Int. J. Mol. Sci. 2023, 24, 14330. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, P.; Mo, Q.; Luo, W.; Du, Z.; Jiang, J.; Yang, S.; Zhao, L.; Gong, Q.; Wang, Y. Comprehensive analysis of full-length transcriptomes of Schizothorax prenanti by single-molecule long-read sequencing. Genomics 2021, 114, 456–464. [Google Scholar] [CrossRef]
- Yang, L.; Zhi, S.; Yang, G.; Qin, C.; Zhao, W.; Niu, M.; Zhang, W.; Tang, W.; Yan, X.; Zhang, Y. Molecular identification of FNDC5 and effect of irisin on the glucose metabolism in common carp (Cyprinus carpio L.). Gen. Comp. Endocrinol. 2021, 301, 113647. [Google Scholar] [CrossRef]
- Chhetri, G.; Kalita, P.; Tripathi, T. An efficient protocol to enhance recombinant protein expression using ethanol in Escherichia coli. MethodsX 2015, 2, 385–391. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D.L. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, X.; Chen, Q.; Li, Q.; Li, J.; Cui, K.; Zhang, Y.; Kong, A.; Zhang, Y.; Wan, M.; Mai, K.; et al. Effects of High Levels of Dietary Linseed Oil on the Growth Performance, Antioxidant Capacity, Hepatic Lipid Metabolism, and Expression of Inflammatory Genes in Large Yellow Croaker (Larimichthys crocea). Front. Physiol. 2021, 12, 631850. [Google Scholar] [CrossRef]
- Ni, J.; Zhu, P.; Mo, Q.; Luo, W.; Du, Z.; Jiang, J.; Yang, S.; Zhao, L.; Gong, Q.; Wang, Y. Dynamic Transcriptomic Profiling during Liver Development in Schizothorax Prenanti. Front. Physiol. 2022, 13, 928858. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasma Parameters | Control | 50 ng/g BW | 100 ng/g BW | 200 ng/g BW |
|---|---|---|---|---|
| Glucose (mmol/L) | 7.54 ± 0.74 | 7.32 ± 0.83 | 15.46 ± 2.84 * | 6.75 ± 0.59 |
| TG (mmol/L) | 2.94 ± 0.14 | 3.32 ± 0.07 | 3.67 ± 0.16 * | 4.03 ± 0.27 * |
| TC (mmol/L) | 7.02 ± 0.94 | 8.04 ± 1.27 | 8.84 ± 1.16 * | 9.45 ± 1.07 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Ni, J.; Wang, A.; Zhang, R.; Wang, L. Vascular Endothelial Growth Factor A (VEGFA) Regulates Hepatic Lipid and Glycogen Metabolism in Schizothorax prenanti. Int. J. Mol. Sci. 2023, 24, 15171. https://doi.org/10.3390/ijms242015171
Wang Y, Ni J, Wang A, Zhang R, Wang L. Vascular Endothelial Growth Factor A (VEGFA) Regulates Hepatic Lipid and Glycogen Metabolism in Schizothorax prenanti. International Journal of Molecular Sciences. 2023; 24(20):15171. https://doi.org/10.3390/ijms242015171
Chicago/Turabian StyleWang, Yan, Jiahui Ni, Aiyu Wang, Run Zhang, and Linjie Wang. 2023. "Vascular Endothelial Growth Factor A (VEGFA) Regulates Hepatic Lipid and Glycogen Metabolism in Schizothorax prenanti" International Journal of Molecular Sciences 24, no. 20: 15171. https://doi.org/10.3390/ijms242015171