
BMPR2 Variants Underlie Nonsyndromic Oligodontia
by
 ,
,
Jinglei Zheng
1,†, 
Haochen Liu
1,†,
Miao Yu
1,
Bichen Lin
2,
Kai Sun
1,
Hangbo Liu
1,
Hailan Feng
1,
Yang Liu
1,* and
Dong Han
1,* 1
Department of Prosthodontics, Peking University School and Hospital of Stomatology & National Center of Stomatology & National Clinical Research Center for Oral Diseases & National Engineering Research Center of Oral Biomaterials and Digital Medical Devices, Beijing 100081, China
2
Frist Clinical Division, Peking University School and Hospital of Stomatology & National Center of Stomatology & National Clinical Research Center for Oral Diseases & National Engineering Research Center of Oral Biomaterials and Digital Medical Devices, Beijing 100081, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2023, 24(2), 1648; https://doi.org/10.3390/ijms24021648
Submission received: 2 November 2022
/
Revised: 6 January 2023
/
Accepted: 10 January 2023
/
Published: 13 January 2023
(This article belongs to the Collection Feature Papers in “Molecular Biology”)
Abstract
:Oligodontia manifests as a congenital reduction in the number of permanent teeth. Despite the major efforts that have been made, the genetic etiology of oligodontia remains largely unknown. Bone morphogenetic protein receptor type 2 (BMPR2) variants have been associated with pulmonary arterial hypertension (PAH). However, the genetic significance of BMPR2 in oligodontia has not been previously reported. In the present study, we identified a novel heterozygous variant (c.814C > T; p.Arg272Cys) of BMPR2 in a family with nonsyndromic oligodontia by performing whole-exome sequencing. In addition, we identified two additional heterozygous variants (c.1042G > A; p.Val348Ile and c.1429A > G; p.Lys477Glu) among a cohort of 130 unrelated individuals with nonsyndromic oligodontia by performing Sanger sequencing. Functional analysis demonstrated that the activities of phospho-SMAD1/5/8 were significantly inhibited in BMPR2-knockout 293T cells transfected with variant-expressing plasmids, and were significantly lower in BMPR2 heterozygosity simulation groups than in the wild-type group, indicating that haploinsufficiency may represent the genetic mechanism. RNAscope in situ hybridization revealed that BMPR2 transcripts were highly expressed in the dental papilla and adjacent inner enamel epithelium in mice tooth germs, suggesting that BMPR2 may play important roles in tooth development. Our findings broaden the genetic spectrum of oligodontia and provide clinical and genetic evidence supporting the importance of BMPR2 in nonsyndromic oligodontia.
1. Introduction
Nonsyndromic oligodontia is an inherited developmental anomaly affecting craniofacial organs and characterized by agenesis of six or more permanent teeth (excluding the third molars) with or without abnormal tooth morphology, affecting the maxillofacial function, orofacial aesthetics, and mental health [1]. The prevalence of nonsyndromic oligodontia is estimated to be 0.25% of Asian populations in China and 0.14% of Caucasian populations in North America, Australia, and Europe [2].
Genetics is widely recognized as a major factor responsible for developmental defects in the tooth germ. To date, only seven genes (MSX1, OMIM #106600; PAX9, OMIM #604625; EDA, OMIM #313500; WNT10A, OMIM #150400; LRP6, OMIM #616724; WNT10B, OMIM #617073; and GREM2, OMIM #617275) have been confirmed as causative factors associated with nonsyndromic oligodontia [3,4,5,6,7,8,9] and there is evidence to support the candidacy of seven genes (AXIN2, EDAR, EDARADD, DKK1, BMP2, BMP4, and KDF1) that may play a role in nonsyndromic oligodontia [10]. Despite considerable efforts, the genetic etiology of nonsyndromic oligodontia remains largely unknown, suggesting the existence of unidentified related genes and molecular mechanisms. Presumably, additional components in several defined key pathways, such as WNT, TGF-β/BMP, and Hedgehog, and FGF signaling, which are involved in tooth development, may be associated with nonsyndromic oligodontia pathogenesis [11].
The bone morphogenetic protein receptor type 2 (BMPR2; OMIM *600799) gene encodes a transforming growth factor-β (TGF-β) type II receptor for bone morphogenetic proteins (BMPs). BMPR2 binds to type I receptors and BMPs to form a tetrameric complex that initiates BMP/SMAD-mediated signaling and participates in embryonic development, vasculogenesis, and bone formation [12,13]. BMPR2−/− mice consistently exhibit severe pulmonary hypertension, skeletal disorders, and embryonic lethality [14]. In humans, variation in the BMPR2 gene is the major cause of pulmonary arterial hypertension (PAH, OMIM #178600) and pulmonary venoocclusive disease 1 (OMIM #265450) [15,16]; however, no variation has been identified in individuals with oligodontia.
This study is the first to identify novel BMPR2 variants in individuals with nonsyndromic oligodontia via whole-exome sequencing (WES). Moreover, we performed in vitro functional analysis and investigated the spatiotemporal expression of BMPR2 in mouse tooth germ development stages. Our results indicate that BMPR2 can be considered a novel pathogenic gene of nonsyndromic oligodontia.
2. Results
2.1. Clinical Findings and Variant Identification
Tooth phenotypes, family diagrams, and DNA sequencing chromatograms of the four pedigrees are presented in Figure 1. The predictions of pathogenic variants and ACMG classifications of the novel BMPR2 variants are summarized in Table 1.
In a four-generation family (family #537) with inherited nonsyndromic oligodontia, the 18-year-old male proband (IV-2) had a congenital absence of six permanent teeth, two retained deciduous teeth, and one cone-shaped maxillary lateral incisor on the right side (Figure 1A–F). The mother of the proband (III-4) had a congenital absence of 12 permanent teeth (Figure 1G), while the father (III-5) had normal dentition. III-4 confirmed that her mother (II-7), grandmother (I-2), maternal aunts (II-3 and II-5), maternal uncle (II-2), and cousin (III-1) had nonsyndromic oligodontia (Figure 1H). Since no deleterious variants of known oligodontia-associated genes were found in WES, we focused on the other undetermined gene(s) in pathways related to tooth development. We identified a novel BMPR2 heterozygous missense variant: c.814C > T; p.Arg272Cys (Table 1). By performing Sanger sequencing, the variant was validated in members with available DNA samples. The sequencing results revealed that the heterozygous variant segregated with the affected individuals, but not with the unaffected family members, suggesting an autosomal dominant mode of inheritance (Figure 1I). Moreover, we performed non-invasive examinations with echocardiography and electrocardiograms on the proband and his mother, and the results showed no abnormalities in the structures and systems of their hearts and pulmonary arteries (Figure S1A–D,H–K). However, non-invasive examination has certain limitations; in the future, further invasive right heart catheterization will be necessary for the diagnosis of PAH. Bone mineral density measurement using dual-energy X-ray absorptiometry (DEXA) showed a decreasing trend in bone mineral density and an increasing risk of osteoporosis in both the proband and his mother (Figure S1E–G,L–N). Notably, the impact of loss of BMPR2 in mice was previously reported as associated with bone formation [17], which is consistent with our findings in humans.
To further explore the prevalence of the BMPR2 variants in unrelated individuals with oligodontia and evaluate whether BMPR2 is a candidate pathogenic gene, we screened the whole coding region of BMPR2 in a cohort of 130 individuals with undefined etiology. Two individuals were found to carry two additional novel BMPR2 variants (Figure 1J–O), indicating that the detection rate of BMPR2 in nonsyndromic oligodontia was 1.5%.
Proband #615, a 26-year-old woman, presented with twelve congenitally missing permanent teeth and three retained deciduous teeth (Figure 1J). A novel BMPR2 de novo heterozygous missense variant, c.1042G > A; p.Val348Ile, was identified in this proband, whereas it was not detected in her parents with normal dentition (Figure 1K,L, Table 1). Proband #531, a 26-year-old woman, had agenesis of fifteen permanent teeth and one retained deciduous tooth (Figure 1M). She carried another novel BMPR2 de novo heterozygous missense variant, c.1429A > G; p.Lys477Glu, while her parents showed normal dentition (Figure 1N,O, Table 1).
2.2. Bioinformatics and Conservation Analysis
The three novel variants were all located in the kinase domain of the BMPR2 protein, which was encoded by BMPR2 exons 5–11 (Figure 2A,B). According to SIFT, PolyPhen-2, MutationTaster, and the standards of ACMG pathogenic classification, the BMPR2 p.Arg272Cys variant was likely pathogenic and the BMPR2 p.Val348Ile and p.Lys477Glu variants were pathogenic (Table 1). Multi-species alignments of the BMPR2 amino acid sequence showed that Arg272, Val348, and Lys477 were highly conserved during evolution (Figure 2C).
2.3. Three-Dimensional Conformational Analysis
Homology modeling of the structures of the kinase domain was performed to analyze the conformational effects of BMPR2 variants (Figure 2D–I). The missense variant Arg272Cys led to the basic residue Arg272 being substituted with a Cys, a neutral amino acid with a sulfhydryl group, which was considerably shorter than Arg, resulting in a significant conformational change in the β-sheet near the 272nd residue (Figure 2D,D’,E,E’). The Val348Ile variant resulted in the residue Val348 being substituted with Ile, an amino acid with a longer side-chain than that of Val, which may affect the interaction of the 348th residue with the surrounding residues (Figure 2F,F’,G,G’). The Lys477Glu variant led to the basic residue Lys477 being substituted with a Glu, an acidic amino acid with a shorter side-chain than that of Lys, which may result in a conformational change in the α-helix near the 477th residue (Figure 2H,H’,I,I’).
2.4. Genetic Mechanism Explorations of BMPR2 Variants in Oligodontia
To evaluate the functional effects of the three BMPR2 variants, we constructed the BMPR2-knockout 293T cells and established a model of a heterozygous BMPR2 variant in vitro. Western blotting showed that in BMPR2-knockout 293T cells, endogenous BMPR2 expression at the protein level was reduced by approximately 90% (p < 0.0001; Figure 3A,B). Subsequently, successful expression of wild-type and all variant BMPR2-GFP fusion proteins was detected in transfected BMPR2-knockout 293T cells at the predicted weight of 141 kDa (Figure 3C). Functional analyses revealed that the expression of phospho-SMAD1/5/8 was significantly inhibited in BMPR2-knockout 293T cells transfected with Arg272Cys, Val348Ile, and Lys477Glu variant plasmids when compared to that in cells transfected with wild-type plasmids (p < 0.0001; Figure 3C,D).
To further explore the pathogenic mechanism of heterozygous BMPR2 variants associated with oligodontia, we simulated the BMPR2 heterozygosity found in oligodontia in vitro and confirmed the successful expression of heterozygotes via Western blotting (Figure 3E). Activities of phospho-SMAD1/5/8 in three heterozygous variant groups (transfected with 0.5 µg wild-type plasmid and 0.5 µg variant plasmids) were approximately 30% lower than those in the 1.0 µg wild-type plasmid-transfected group, while the activities were approximately 20% higher than those in the 0.5 µg wild-type plasmid-transfected group (p < 0.0001; Figure 3E,F), suggesting that haploinsufficiency may represent the genetic mechanism in BMPR2-associated nonsyndromic oligodontia.
2.5. Expression Pattern of Bmpr2 during Murine Molar Development
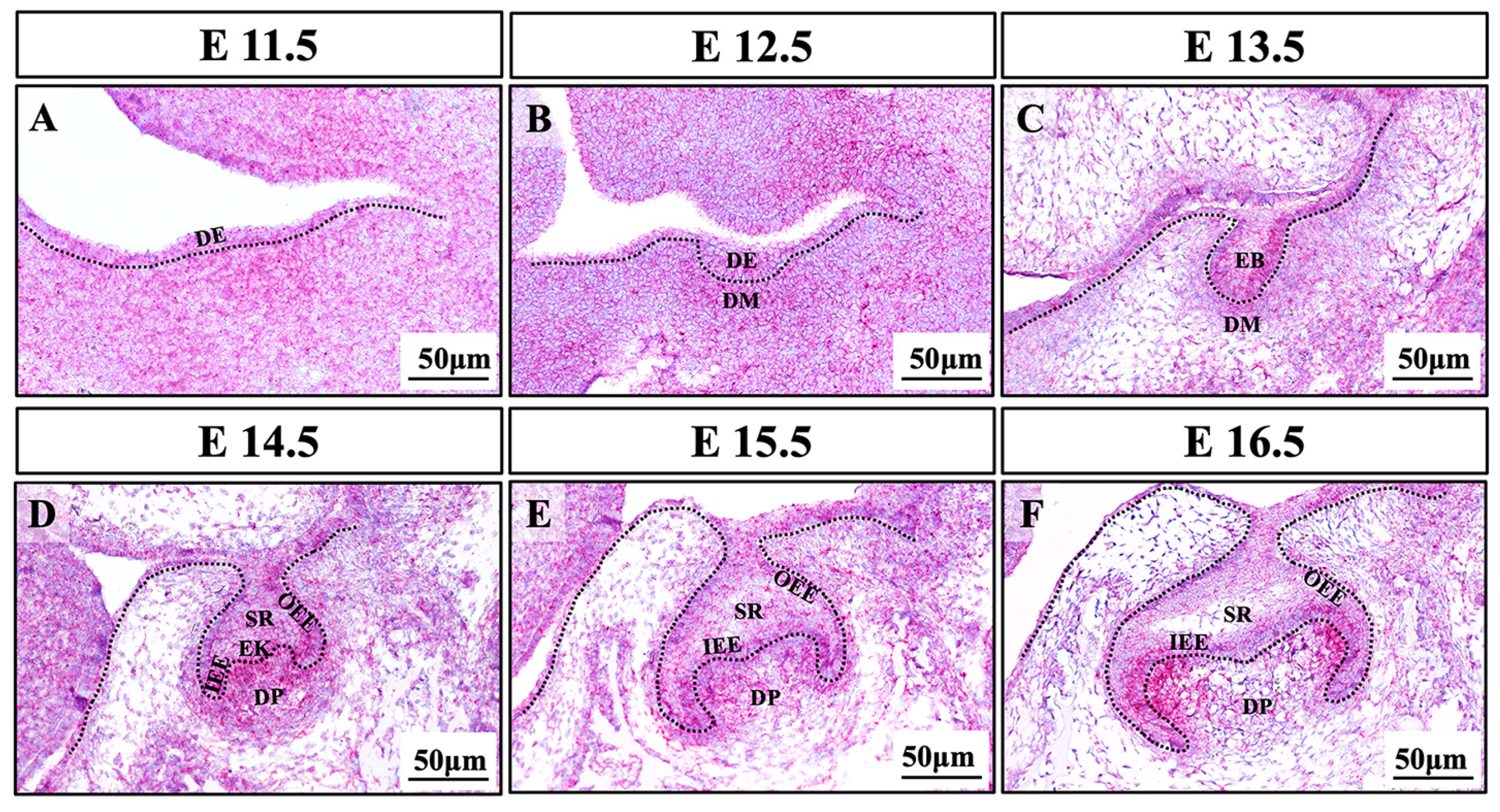
RNAscope in situ hybridization assay was used to reveal the precise spatiotemporal expression of Bmpr2 in mouse first mandibular molars at serial developmental stages. Bmpr2 was widely expressed in the dental epithelium and mesenchyme during the process of tooth development (E11.5–E16.5, Figure 4A–F). At the cap stage (E14.5), Bmpr2 expression was detected in the enamel organ, dental papilla, and dental follicle (Figure 4D). Notably, at the bell stage (E15.5–E16.5), Bmpr2 was highly expressed in the inner enamel epithelium, outer enamel epithelium, secondary enamel knots, and adjacent dental papilla cells, while it was less expressed in the stellate reticulum (Figure 4E,F).
3. Discussion
In this study, we identified three novel BMPR2 missense variants, p.Arg272Cys, p.Val348Ile, and p.Lys477Glu, in an autosomal dominant family and two isolated patients with nonsyndromic oligodontia. Our study is the first to link the genetic importance of BMPR2 to human oligodontia.
Although previous studies demonstrated the association of BMPR2 variants with PAH, all the individuals with BMPR2 variants in our study presented oligodontia with no PAH-associated symptoms. Using non-invasive examinations of echocardiography and electrocardiograms, no PAH-related abnormalities were found in proband #537 and his mother. Previous studies showed that heterozygous variations in the BMPR2 gene are found in nearly 70% of families with heritable PAH [18] and in 25% of patients with sporadic disease [19]. PAH is more common in women (female:male ratio of 1.7:1) [20]. However, the penetrance of PAH is incomplete; only approximately 10–20% of individuals with BMPR2 variants develop the disease during their lifetime, suggesting that PAH development is triggered by other genetic or environmental factors [21,22]. Therefore, BMPR2 variants can be considered as risk factors in patients, suggesting that they may be susceptible to PAH in future.
The BMPR2 protein consists of extracellular, transmembrane, kinase, and C-terminal cytoplasmic domains [12]. BMPR2 kinase domain is crucial for BMP signal transduction and it participates in BMPR2 kinase phosphorylation, type I receptor (BMPR1) activation, activated receptor complex formation, and ultimately, SMAD1/5/8 phosphorylation. In this study, all the variants—Arg272Cys, Val348Ile, and Lys477Glu—caused different conformational changes in the BMPR2 kinase domain, demonstrating that the kinase domain represents a germline variant hotspot in nonsyndromic oligodontia and plays a crucial role in the pathogenesis of nonsyndromic oligodontia.
Moreover, according to an analysis of human disease genes, approximately 50% of the mutations in membrane receptors were dominant [23]. Genetic dominance commonly results from the dominant-negative effect or haploinsufficiency. The dominant-negative effect refers to a defective subunit adversely affecting the activity of the normal one, whereas haploinsufficiency describes a situation of reduced activity by a heterozygous null allele [24]. It is sometimes difficult to determine a priori whether haploinsufficiency or the dominant-negative effect is responsible for the phenotypes, particularly in the case of intragenic point mutations. In this study, we analyzed the expression of BMPR2 by Western blotting, and demonstrated that the inactivated BMP/phospho-SMAD1/5/8 signaling was caused by the novel BMPR2 heterozygous variants, suggesting that the effects of BMPR2 are allele dose-dependent. Further results showed that compared to the wild-type group, the activities of phospho-SMAD1/5/8 in heterozygous variant groups were reduced due to a heterozygous null allele, but were no less than in a normal allele group (simulated by the 0.5 µg wild-type group). These findings indicate that a haploinsufficiency effect contributes to the genetic mechanism of BMPR2-associated nonsyndromic oligodontia, rather than the dominant-negative effect.
When reviewing previous research, we found that Bmpr2−/− mice died before E9.5, that is, prior to bone development [14], so skeletal or dental defects could not be assessed. Since BMPR2 variants are broadly considered to be associated with PAH in humans, studies of mouse models frequently focused on the circulatory system. In recent years, many molecules in the BMP signaling pathway have been reported to be related to tooth development. Previous studies have reported that variants of BMP4 are related to oligodontia in humans [25,26]. During mouse embryonic development, conservative Bmp signaling regulates odontogenesis via reciprocal epithelial–mesenchymal interaction. Disruption of Bmp signaling causes retardation of early tooth development [27]. Mice with K14-Cre; Bmpr1afl/fl exhibit the retardation of tooth development, loss of posterior limbs, and hair defects [28]. Moreover, conditional ablation of Bmp2 in enamel using Osx-Cre; Bmp2fl/fl transfection results in decreased enamel mineralization and thickness, which confirms the role of Bmp2 in the regulation of postnatal enamel formation [29]. Additionally, Wnt1-Cre; Bmp4fl/fl mice show an arrest in the development of mandibular molars at the bud stage [30]. However, the genetic role of Bmpr2 in tooth development and oligodontia remains unknown. Using RNAscope analysis, we found that Bmpr2 was highly expressed in the primary enamel knot, secondary enamel knot, inner and outer enamel epithelium, and adjacent mesenchymal cells. These data demonstrate that Bmpr2 may regulate the epithelial–mesenchymal interactions and cell differentiation during tooth development. These results are basically consistent with previous findings on Bmpr2 protein expression in mice [31], and the precise expression of Bmpr2 in our study complements earlier data at the mRNA level. However, the definite role of BMPR2 in the regulation of tooth development needs to be further investigated in the future.
4. Materials and Methods
4.1. Recruitment of Subjects
A four-generation family with nonsyndromic oligodontia was recruited at the Department of Prosthodontics, Peking University School of Stomatology (Beijing, China) during medical treatment of the proband. No evidence of other general abnormalities was found in this family. Additionally, a cohort of 130 unrelated individuals with nonsyndromic oligodontia from our database was enrolled for the expanded genetic screening. All individuals confirmed the absence of tooth extraction or loss. Informed consent was obtained from all the participants. This study and associated research protocols were approved by the Ethics Committee of the Peking University School and Hospital of Stomatology (PKUSSIRB-202162021).
4.2. WES
Genomic DNA was isolated from peripheral blood lymphocytes using a DNA Whole-blood Mini Kit (BioTek, Beijing, China) following the manufacturer’s instructions. WES was performed by Angen Gene Medicine Technology (Beijing, China) to identify potential pathogenic gene variations. The selection criteria for pathogenic genes were as follows: (1) genes involved in orodental development were screened; (2) short insertions or deletions and sequence nucleotide variants, including nonsense, missense, and splicing variations, with a minor allele frequency ≤ 0.01 in population frequency databases, such as the Genome Aggregation Database (gnomAD, https://gnomad.broadinstitute.org (accessed on 13 March 2021)) [32] and single nucleotide polymorphisms (dbSNP, https://www.ncbi.nlm.nih.gov/SNP/ (accessed on 13 March 2021)); (3) the potential pathogenicity was predicted via bioinformatics analysis using MutationTaster (http://www.mutationtaster.org (accessed on 13 March 2021)), Sorting Intolerant from Tolerant (SIFT, http://provean.jcvi.org (accessed on 13 March 2021)), and Polymorphism Phenotyping v2 (PolyPhen-2, http://genetics.bwh.harvard.edu/pph2/ (accessed on 13 March 2021)), and then the screened variants were classified according to the 2015 American College of Medical Genetics and Genomics guidelines [33].
4.3. Sanger Sequencing and Genetic Screening
From the WES results, we identified a novel variant of BMPR2 (RefSeq NM_001204), which was validated via Sanger sequencing and familial co-segregation. Subsequently, to further confirm the genetic role of BMPR2 in oligodontia, gene screening was performed via Sanger sequencing using samples of 130 unrelated individuals with nonsyndromic oligodontia with undefined etiology in our clinical database. Polymerase chain reaction (PCR) was performed to amplify the coding region of BMPR2 (primers and amplification conditions can be provided) using Taq PCR Master Mix (BioTek, Beijing, China). The PCR products were sequenced by Tsingke Biological Technology (Beijing, China).
4.4. Bioinformatic and Conservative Analysis
Predictions of the putative pathogenic BMPR2 variants were performed using bioinformatic tools, including SIFT, PolyPhen-2, and MutationTaster. Conservative analysis was performed using the ClustalX 2.1 program based on the multiple alignments of BMPR2 amino acid sequences (NP_001195) among different species, which were obtained from the National Center for Biotechnology Information (National Institutes of Health, Bethesda, MD, USA).
4.5. Three-Dimensional Structural Modeling
Since the identified BMPR2 variants were all distributed in the kinase domain of wild-type BMPR2 protein, an optimal template was selected for the structural homology modeling of the kinase domain (from amino acid 203 to 504) using SWISS-MODEL (https://swiss-model.expasy.org (accessed on 21 February 2021)). Changes in the three-dimensional structure of BMPR2 protein were observed using PyMOL software (Molecular Graphics System; DeLano Scientific, Palo Alto, CA, USA).
4.6. Plasmid Construction
To construct the wild-type BMPR2 plasmid, the full-length coding region (3118 bp) of the human BMPR2 gene was introduced to the green fluorescent protein (GFP)-expressing pEGFP-C1 vector between the 5′-EcoRI and 3′-XhoI sites. The wild-type plasmid pEGFP-C1-BMPR2, as well as the three variant plasmids, pEGFP-C1-Arg272Cys, pEGFP-C1-Val348Ile, and pEGFP-C1-Lys477Glu, were synthesized at the Beijing Genomic Institute (BGI, Beijing, China), and the entire sequence of the constructs was confirmed by BGI.
4.7. Cell Preparation and Western Blot Analysis
To eliminate endogenous interference, we used the clustered, regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) system to delete endogenous BMPR2 from 293T cells obtained from Tsingke Biological Technology. Subsequently, the BMPR2-knockout 293T cells (BKOTCs) were transfected with 1 μg empty vector, 1 μg wild-type plasmid, 1 μg variant plasmids, 0.5 μg wild-type plasmid, or 0.5 μg wild-type plasmid, together with 0.5 μg of each variant plasmid, using Lipofectamine 3000 (Thermo Fisher Scientific, Waltham, MA, USA).
After 48 h of transfection, recombinant GFP was observed under a fluorescence microscope (Olympus BX51; Olympus, Tokyo, Japan), and the transfected cells were lysed using protein lysis buffer (Beyotime Technology, Shanghai, China) containing phosphatase inhibitor (Solarbio, Beijing, China). Western blot analysis was performed using cell lysates, and proteins were incubated with anti-GFP (Abcam, Cambridge, United Kingdom), anti-p-SMAD1/5/8 (Cell Signaling Technology, Danvers, MA, USA), anti-SMAD1 (Cell Signaling Technology), and anti-glyceraldehyde 3-phosphate dehydrogenase antibodies (Abcam). The proteins were visualized using a chemiluminescent ECL kit (Sigma–Aldrich Corp., St. Louis, MO, USA) and semi-quantified using Image J software (National Institutes of Health). Experiments were performed in triplicate. Data were analyzed with Student’s t test using SPSS (IBM, Armonk, NY, USA). Results were presented as the mean ± standard deviation (n = 3), and p < 0.01 was considered significant.
4.8. Tooth Germ Preparation and RNAscope In Situ Hybridization Analysis
To detect the temporal and spatial expression pattern of Bmpr2 during early tooth development, timed-pregnant ICR mice (Department of Laboratory Animal Science, Peking University Health Science Center, Beijing, China) at the stages of embryonic (E) days 11.5 (E11.5)–E16.5 were euthanized. The embryonic heads were dissected, fixed in 4% paraformaldehyde for 24 h, dehydrated, embedded in paraffin, and serially sectioned (at a thickness of 5 μm) in the coronal plane. RNAscope Probe-Mm-BMPR2 (Advanced Cell Diagnostics, Newark, CA, USA) was designed to target the mouse Bmpr2 mRNA using an RNAscope 2.5 HD detection reagents-RED kit (Advanced Cell Diagnostics, 322360).
5. Conclusions
In conclusion, we provide the first genetic evidence of the etiological correlation between BMPR2 heterozygous variants and nonsyndromic oligodontia. The spatiotemporal expression pattern of Bmpr2 during tooth development suggests that BMPR2 may play a crucial role in tooth morphogenesis. Our findings further extend the panel of candidate genes available for the genetic screening of nonsyndromic oligodontia and provide new insights into the molecular mechanisms of nonsyndromic oligodontia.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms24021648/s1.
Author Contributions
Conceptualization, Y.L. and D.H.; methodology, J.Z., M.Y. and D.H.; software, J.Z., M.Y., H.L. (Haochen Liu) and B.L.; validation, H.L. (Haochen Liu), B.L. and Y.L.; formal analysis, J.Z. and M.Y.; investigation, J.Z. and M.Y.; resources, H.L. (Haochen Liu) and B.L.; data curation, J.Z. and D.H.; writing—original draft preparation, J.Z., M.Y., H.L. (Haochen Liu), B.L, Y.L. and D.H.; writing—review and editing, J.Z., M.Y., H.L. (Haochen Liu), B.L., Y.L. and D.H.; visualization, J.Z., M.Y., K.S. and H.L. (Hangbo Liu); supervision, H.F.; project administration, Y.L. and D.H.; funding acquisition, H.L. (Haochen Liu), M.Y., Y.L. and D.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant numbers 81970902, 82100976, 81600851, and 81600846.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Peking University School and Hospital of Stomatology (PKUS-SIRB-202162021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The variants identified in this study were submitted to the ClinVar database (submission ID SCV002578204 and SCV002578205). WES data are available from the SRA database (accession number PRJNA900358).
Acknowledgments
We are thankful to the participating families.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vastardis, H.; Karimbux, N.; Guthua, S.W.; Seidman, J.G.; Seidman, C.E. A human MSX1 homeodomain missense mutation causes selective tooth agenesis. Nat. Genet. 1996, 13, 417–421. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, H.; Camhi, H.; Seymen, F.; Koruyucu, M.; Kasimoglu, Y.; Kim, J.W.; Kim-Berman, H.; Yuson, N.M.R.; Benke, P.J.; et al. Analyses of oligodontia phenotypes and genetic etiologies. Int J. Oral Sci 2021, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- van den Boogaard, M.J.; Dorland, M.; Beemer, F.A.; van Amstel, H.K. MSX1 mutation is associated with orofacial clefting and tooth agenesis in humans. Nat. Genet. 2000, 24, 342–343. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Groppe, J.C.; Wu, J.; Ogawa, T.; Mues, G.; D’Souza, R.N.; Kapadia, H. Pathogenic mechanisms of tooth agenesis linked to paired domain mutations in human PAX9. Hum. Mol. Genet. 2009, 18, 2863–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, M.C.; Biancalana, V.; Ginisty, D.; Mandel, J.L.; Calvas, P. Mutational spectrum of the ED1 gene in X-linked hypohidrotic ectodermal dysplasia. Eur J. Hum. Genet. 2001, 9, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Bohring, A.; Stamm, T.; Spaich, C.; Haase, C.; Spree, K.; Hehr, U.; Hoffmann, M.; Ledig, S.; Sel, S.; Wieacker, P.; et al. WNT10A mutations are a frequent cause of a broad spectrum of ectodermal dysplasias with sex-biased manifestation pattern in heterozygotes. Am. J. Hum. Genet. 2009, 85, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Massink, M.P.; Creton, M.A.; Spanevello, F.; Fennis, W.M.; Cune, M.S.; Savelberg, S.M.; Nijman, I.J.; Maurice, M.M.; van den Boogaard, M.J.; van Haaften, G. Loss-of-Function Mutations in the WNT Co-receptor LRP6 Cause Autosomal-Dominant Oligodontia. Am. J. Hum. Genet. 2015, 97, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Yang, W.; Han, D.; Wang, X.; Guo, S.; Li, J.; Li, F.; Zhang, X.; Wong, S.W.; Bai, B.; et al. Mutations in WNT10B Are Identified in Individuals with Oligodontia. Am. J. Hum. Genet. 2016, 99, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantaputra, P.N.; Kaewgahya, M.; Hatsadaloi, A.; Vogel, P.; Kawasaki, K.; Ohazama, A.; Ketudat Cairns, J.R. GREMLIN 2 Mutations and Dental Anomalies. J. Dent. Res. 2015, 94, 1646–1652. [Google Scholar] [CrossRef]
- Yu, M.; Wong, S.W.; Han, D.; Cai, T. Genetic analysis: Wnt and other pathways in nonsyndromic tooth agenesis. Oral Dis. 2019, 25, 646–651. [Google Scholar] [CrossRef]
- Yin, W.; Bian, Z. The Gene Network Underlying Hypodontia. J. Dent. Res. 2015, 94, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Park, S.Y.; Chang, H.R.; Jung, E.Y.; Munkhjargal, A.; Lim, J.S.; Lee, M.S.; Kim, Y. Clinical significance linked to functional defects in bone morphogenetic protein type 2 receptor, BMPR2. BMB Rep. 2017, 50, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Puerto, M.C.; Iyengar, P.V.; Garcia de Vinuesa, A.; Ten Dijke, P.; Sanchez-Duffhues, G. Bone morphogenetic protein receptor signal transduction in human disease. J. Pathol. 2019, 247, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beppu, H.; Kawabata, M.; Hamamoto, T.; Chytil, A.; Minowa, O.; Noda, T.; Miyazono, K. BMP type II receptor is required for gastrulation and early development of mouse embryos. Dev. Biol. 2000, 221, 249–258. [Google Scholar] [CrossRef] [Green Version]
- International, P.P.H.C.; Lane, K.B.; Machado, R.D.; Pauciulo, M.W.; Thomson, J.R.; Phillips, J.A., 3rd; Loyd, J.E.; Nichols, W.C.; Trembath, R.C. Heterozygous germline mutations in BMPR2, encoding a TGF-beta receptor, cause familial primary pulmonary hypertension. Nat. Genet. 2000, 26, 81–84. [Google Scholar]
- Rigelsky, C.M.; Jennings, C.; Lehtonen, R.; Minai, O.A.; Eng, C.; Aldred, M.A. BMPR2 mutation in a patient with pulmonary arterial hypertension and suspected hereditary hemorrhagic telangiectasia. Am. J. Med. Genet. A 2008, 146, 2551–2556. [Google Scholar] [CrossRef]
- Lowery, J.W.; Intini, G.; Gamer, L.; Lotinun, S.; Salazar, V.S.; Ote, S.; Cox, K.; Baron, R.; Rosen, V. Loss of BMPR2 leads to high bone mass due to increased osteoblast activity. J. Cell Sci. 2015, 128, 1308–1315. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.D.; Aldred, M.A.; James, V.; Harrison, R.E.; Patel, B.; Schwalbe, E.C.; Gruenig, E.; Janssen, B.; Koehler, R.; Seeger, W.; et al. Mutations of the TGF-beta type II receptor BMPR2 in pulmonary arterial hypertension. Hum. Mutat. 2006, 27, 121–132. [Google Scholar] [CrossRef]
- Newman, J.H.; Trembath, R.C.; Morse, J.A.; Grunig, E.; Loyd, J.E.; Adnot, S.; Coccolo, F.; Ventura, C.; Phillips, J.A., 3rd; Knowles, J.A.; et al. Genetic basis of pulmonary arterial hypertension: Current understanding and future directions. J. Am. Coll Cardiol. 2004, 43 (Suppl S12), 33–39. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.J. Pulmonary hypertension. Jama 2012, 308, 1366–1374. [Google Scholar] [CrossRef]
- Machado, R.D.; Eickelberg, O.; Elliott, C.G.; Geraci, M.W.; Hanaoka, M.; Loyd, J.E.; Newman, J.H.; Phillips, J.A., 3rd; Soubrier, F.; Trembath, R.C.; et al. Genetics and genomics of pulmonary arterial hypertension. J. Am. Coll Cardiol. 2009, 54 (Suppl S1), 32–42. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Hong, K.H.; Kim, Y.H.; Kim, M.J.; Song, C.; Kim, M.J.; Kim, S.J.; Raizada, M.K.; Oh, S.P. SMAD1 deficiency in either endothelial or smooth muscle cells can predispose mice to pulmonary hypertension. Hypertension 2013, 61, 1044–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Sanchez, G.; Childs, B.; Valle, D. Human disease genes. Nature 2001, 409, 853–855. [Google Scholar] [CrossRef] [Green Version]
- Veitia, R.A.; Caburet, S.; Birchler, J.A. Mechanisms of Mendelian dominance. Clin. Genet. 2018, 93, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Wang, H.; Fan, Z.; Xie, C.; Liu, H.; Liu, Y.; Han, D.; Wong, S.W.; Feng, H. BMP4 mutations in tooth agenesis and low bone mass. Arch. Oral Biol 2019, 103, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lu, Y.; Mues, G.; Wang, S.; Bonds, J.; D’Souza, R. Functional evaluation of a novel tooth agenesis-associated bone morphogenetic protein 4 prodomain mutation. Eur. J. Oral Sci. 2013, 121, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, L.; Zheng, Y.; Yuan, G.; Yang, G.; He, F.; Chen, Y. BMP activity is required for tooth development from the lamina to bud stage. J. Dent. Res. 2012, 91, 690–695. [Google Scholar] [CrossRef] [Green Version]
- Andl, T.; Ahn, K.; Kairo, A.; Chu, E.Y.; Wine-Lee, L.; Reddy, S.T.; Croft, N.J.; Cebra-Thomas, J.A.; Metzger, D.; Chambon, P.; et al. Epithelial Bmpr1a regulates differentiation and proliferation in postnatal hair follicles and is essential for tooth development. Development 2004, 131, 2257–2268. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Yang, G.; Yuan, G.; Gluhak-Heinrich, J.; Yang, W.; Wang, L.; Chen, Z.; Schulze McDaniel, J.; Donly, K.J.; Harris, S.E.; et al. Abnormalities in the enamel in bmp2-deficient mice. Cells Tissues Organs 2011, 194, 216–221. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Zhou, J.; Gao, Y.; Baek, J.A.; Martin, J.F.; Lan, Y.; Jiang, R. Roles of Bmp4 during tooth morphogenesis and sequential tooth formation. Development 2013, 140, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Nadiri, A.; Kuchler-Bopp, S.; Perrin-Schmitt, F.; Lesot, H. Expression patterns of BMPRs in the developing mouse molar. Cell Tissue Res. 2006, 324, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. Off. J. Am. Coll. Med. Genet. 2015, 17, 405–424. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Clinical features and genetic screening of families with nonsyndromic oligodontia. (A–G) Digital photographs, panoramic radiographs, and missing tooth positions in proband #537 IV-2 (A–F) and his mother III-4 (G); (H,I) genogram and sequencing chromatograms revealed a novel heterozygous BMPR2 variant (c.814C > T; p.Arg272Cys) identified in members with nonsyndromic oligodontia, IV-2, III-4, II-7, II-5, and II-3, which was inherited in an autosomal dominant pattern. Subjects I-1, I-2, I-3, II-1, II-2 II-4, II-6, III-1, III-2, III-3, and IV-1 were not available for clinical evaluation or DNA analysis; (J) panoramic radiograph and schematic of proband #615 II-1; (K,L) genogram and sequencing chromatograms show a de novo heterozygous BMPR2 variant (c.1042G > A; p.Val348Ile) identified in the proband; (M) panoramic radiograph and schematic of proband #531 II-1; (N,O) genogram and sequencing chromatograms show a de novo heterozygous BMPR2 variant (c.1429A > G; p.Lys477Glu) identified in the proband. The white asterisks (*) in the panoramic radiographs and solid squares in the schematics represent the missing permanent teeth. Mand, mandibular. Max, maxillary. The yellow arrows indicate retained deciduous teeth; the red arrows indicate cone-shaped permanent teeth. The Roman numbers on the left side of each genogram represent the numbers of generations in the family. BMPR2, bone morphogenetic protein receptor type 2.
Figure 1.
Clinical features and genetic screening of families with nonsyndromic oligodontia. (A–G) Digital photographs, panoramic radiographs, and missing tooth positions in proband #537 IV-2 (A–F) and his mother III-4 (G); (H,I) genogram and sequencing chromatograms revealed a novel heterozygous BMPR2 variant (c.814C > T; p.Arg272Cys) identified in members with nonsyndromic oligodontia, IV-2, III-4, II-7, II-5, and II-3, which was inherited in an autosomal dominant pattern. Subjects I-1, I-2, I-3, II-1, II-2 II-4, II-6, III-1, III-2, III-3, and IV-1 were not available for clinical evaluation or DNA analysis; (J) panoramic radiograph and schematic of proband #615 II-1; (K,L) genogram and sequencing chromatograms show a de novo heterozygous BMPR2 variant (c.1042G > A; p.Val348Ile) identified in the proband; (M) panoramic radiograph and schematic of proband #531 II-1; (N,O) genogram and sequencing chromatograms show a de novo heterozygous BMPR2 variant (c.1429A > G; p.Lys477Glu) identified in the proband. The white asterisks (*) in the panoramic radiographs and solid squares in the schematics represent the missing permanent teeth. Mand, mandibular. Max, maxillary. The yellow arrows indicate retained deciduous teeth; the red arrows indicate cone-shaped permanent teeth. The Roman numbers on the left side of each genogram represent the numbers of generations in the family. BMPR2, bone morphogenetic protein receptor type 2.

Figure 2.
Location, conservation analysis, and tertiary structure analysis of BMPR2 variants. (A,B) Locations of three identified variants (marked in red) in schematic diagram of BMPR2. Correlations between exons and protein domains are shown in different colors. ECD, extracellular domain; TM, transmembrane domain; KD, kinase domain; CTD, cytoplasmic tail domain; (C) conservation analysis of the three variation sites among different species; (D–I) structural changes in three BMPR2 variants (E,G,I) compared with the wild-type BMPR2 (D,F,H) in the kinase domain. Dashed boxes denote the locations of Arg272 (D), Cys272 (E), Val348 (F), Ile348 (G), Lys477 (H), and Glu477 (I) residues; (D’–I’) higher magnifications of the boxed areas. aa, amino acid. BMPR2, bone morphogenetic protein receptor type 2.
Figure 2.
Location, conservation analysis, and tertiary structure analysis of BMPR2 variants. (A,B) Locations of three identified variants (marked in red) in schematic diagram of BMPR2. Correlations between exons and protein domains are shown in different colors. ECD, extracellular domain; TM, transmembrane domain; KD, kinase domain; CTD, cytoplasmic tail domain; (C) conservation analysis of the three variation sites among different species; (D–I) structural changes in three BMPR2 variants (E,G,I) compared with the wild-type BMPR2 (D,F,H) in the kinase domain. Dashed boxes denote the locations of Arg272 (D), Cys272 (E), Val348 (F), Ile348 (G), Lys477 (H), and Glu477 (I) residues; (D’–I’) higher magnifications of the boxed areas. aa, amino acid. BMPR2, bone morphogenetic protein receptor type 2.
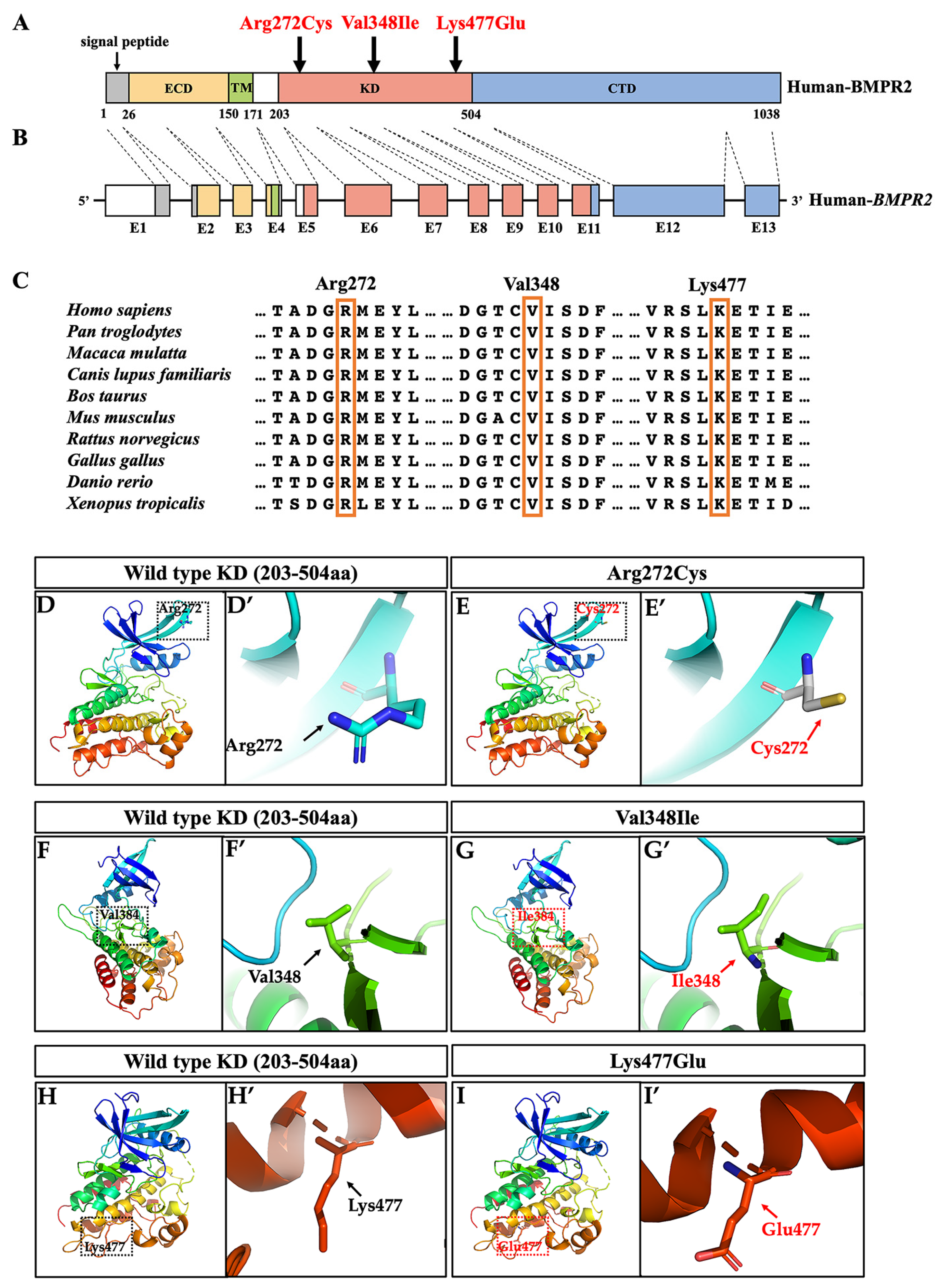
Figure 3.
Genetic mechanisms of BMPR2 variants causing oligodontia. (A,B) Western blot analysis of endogenous BMPR2 gene knockout in 293T cells; (C) expression of BMPR2, p-SMAD1/5/8, and SMAD1 in wild-type and variant groups (Arg272Cys, Val348Ile, and Lys477Glu). An empty vector was transfected into cells as a negative control; (D) quantification of p-SMAD1/5/8 level via densitometry analysis using image J software; (E) expression of BMPR2, p-SMAD1/5/8, and SMAD1 in wild-type, 0.5μg wild-type, and three heterozygous variant groups; (F) quantification of p-SMAD1/5/8 levels by densitometry analysis using image J software. The asterisks denote the difference with statistical significance (*** p < 0.0001) compared to the wild-type group. The results are depicted as the mean ± standard deviation. BMPR2, bone morphogenetic protein receptor type 2.
Figure 3.
Genetic mechanisms of BMPR2 variants causing oligodontia. (A,B) Western blot analysis of endogenous BMPR2 gene knockout in 293T cells; (C) expression of BMPR2, p-SMAD1/5/8, and SMAD1 in wild-type and variant groups (Arg272Cys, Val348Ile, and Lys477Glu). An empty vector was transfected into cells as a negative control; (D) quantification of p-SMAD1/5/8 level via densitometry analysis using image J software; (E) expression of BMPR2, p-SMAD1/5/8, and SMAD1 in wild-type, 0.5μg wild-type, and three heterozygous variant groups; (F) quantification of p-SMAD1/5/8 levels by densitometry analysis using image J software. The asterisks denote the difference with statistical significance (*** p < 0.0001) compared to the wild-type group. The results are depicted as the mean ± standard deviation. BMPR2, bone morphogenetic protein receptor type 2.

Figure 4.
Expression patterns of Bmpr2 during early tooth development in mouse. (A,B) Expression patterns of Bmpr2 detected using RNAscope assay during the dental epithelium thickening stage (E11.5 and E12.5), (C) the bud stage (E13.5), (D) the cap stage (E14.5), and (E,F) the bell stage (E16.5 and E15.5). DE, dental epithelium; DM, dental mesenchyme; DP, dental papilla; DF, dental follicle; EB, epithelial bud; EK, enamel knot; IEE, inner enamel epithelium; OEE, outer enamel epithelium; SR, stellate reticulum; Bmpr2, bone morphogenetic protein receptor type 2. Scale bars: 50 μm.
Figure 4.
Expression patterns of Bmpr2 during early tooth development in mouse. (A,B) Expression patterns of Bmpr2 detected using RNAscope assay during the dental epithelium thickening stage (E11.5 and E12.5), (C) the bud stage (E13.5), (D) the cap stage (E14.5), and (E,F) the bell stage (E16.5 and E15.5). DE, dental epithelium; DM, dental mesenchyme; DP, dental papilla; DF, dental follicle; EB, epithelial bud; EK, enamel knot; IEE, inner enamel epithelium; OEE, outer enamel epithelium; SR, stellate reticulum; Bmpr2, bone morphogenetic protein receptor type 2. Scale bars: 50 μm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Harmful prediction of the variants in BMPR2.
| Patient | Exon | Nucleotide Change | Protein Change | Variation Type | dbSNP | gnomAD (MAF) | SIFT | PolyPhen-2 | Mutation Taster | ACMG Classification (Evidence of Pathogenicity) |
|---|---|---|---|---|---|---|---|---|---|---|
| #537 | 6 | c.814C > T | p.Arg272Cys | Missense | rs773655445 | 0.00003186 | 0.008 (damaging) | 0.999 (probably damaging) | Disease causing | Likely pathogenic (PS3 + PM2 + PP1 + PP3) |
| #615 | 8 | c.1042G > A | p.Val348Ile | Missense | rs201067849 | 0.0005339 | 0.023 (damaging) | 0.715 (possibly damaging) | Disease causing | Pathogenic (PS1 + PS2 + PS3 + PP3) |
| #531 | 11 | c.1429A > G | p.Lys477Glu | Missense | — 1 | — 1 | 0.060 (tolerated) | 1.000 (probably damaging) | Disease causing | Pathogenic (PS2 + PS3 + PM2 + PP3) |
1 Variant was not found in dbSNP or gnomAD; ACMG: American College of Medical Genetics; PS: pathogenic criterion is weighted as strong; PM: pathogenic criterion is weighted as moderate; PP: pathogenic criterion is weighted as supporting.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zheng, J.; Liu, H.; Yu, M.; Lin, B.; Sun, K.; Liu, H.; Feng, H.; Liu, Y.; Han, D. BMPR2 Variants Underlie Nonsyndromic Oligodontia. Int. J. Mol. Sci. 2023, 24, 1648. https://doi.org/10.3390/ijms24021648
AMA Style
Zheng J, Liu H, Yu M, Lin B, Sun K, Liu H, Feng H, Liu Y, Han D. BMPR2 Variants Underlie Nonsyndromic Oligodontia. International Journal of Molecular Sciences. 2023; 24(2):1648. https://doi.org/10.3390/ijms24021648
Chicago/Turabian StyleZheng, Jinglei, Haochen Liu, Miao Yu, Bichen Lin, Kai Sun, Hangbo Liu, Hailan Feng, Yang Liu, and Dong Han. 2023. "BMPR2 Variants Underlie Nonsyndromic Oligodontia" International Journal of Molecular Sciences 24, no. 2: 1648. https://doi.org/10.3390/ijms24021648
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.