
Are the Properties of Bone Marrow-Derived Mesenchymal Stem Cells Influenced by Overweight and Obesity?

Abstract
:1. Introduction
2. Effect on Proliferation and Colony-Forming Ability of BMSCs
3. Effect on Surface Antigen Expression of BMSCs
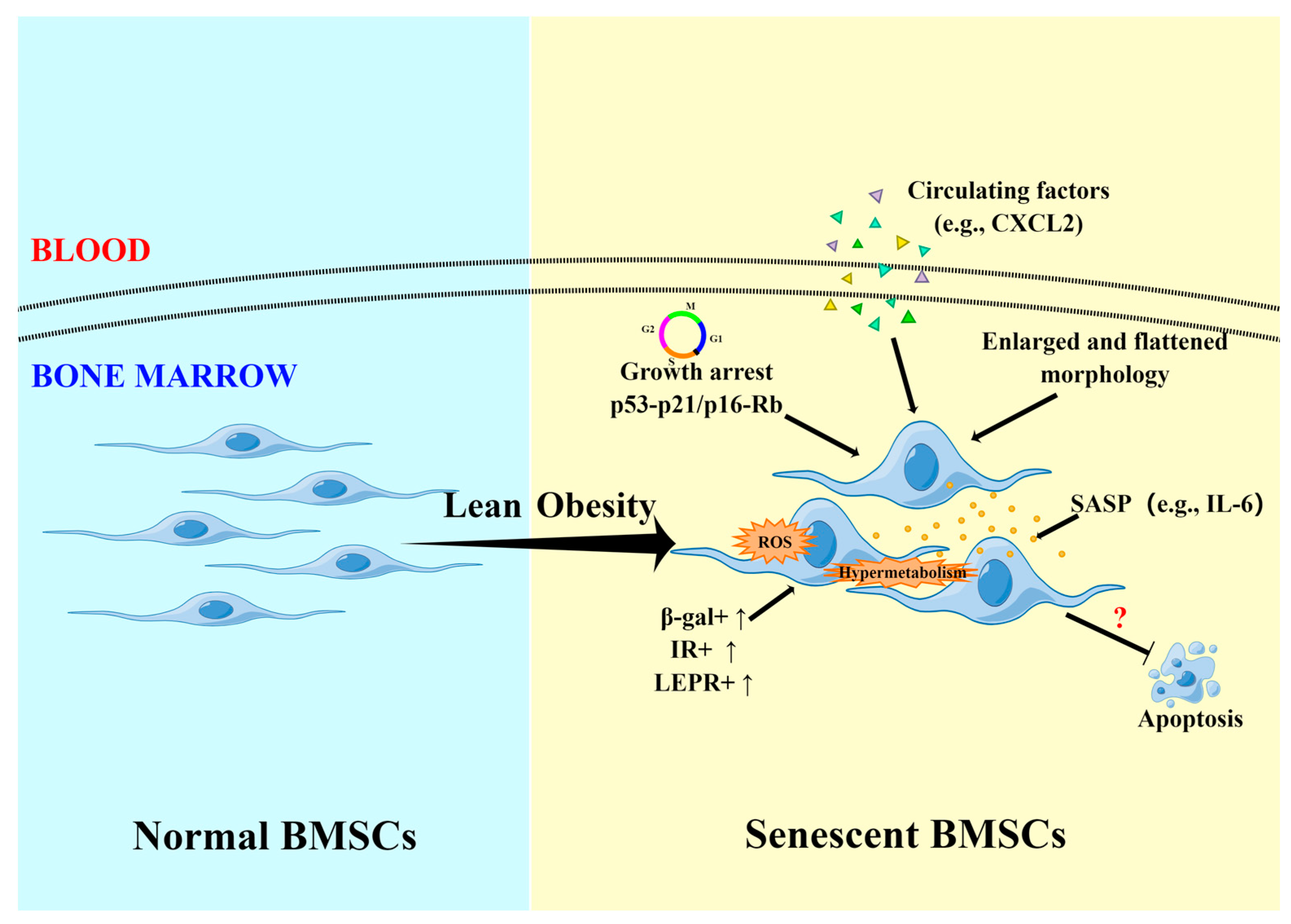
4. Effect on Senescence of BMSCs
5. Effect on Apoptosis of BMSCs
6. Effect on Trilineage Differentiation Potential of BMSCs
6.1. Osteogenic Differentiation
6.2. Adipogenic Differentiation
6.3. Chondrogenic Differentiation
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flegal, K.M.; Kit, B.K.; Orpana, H.; Graubard, B.I. Association of all-cause mortality with overweight and obesity using standard body mass index categories: A systematic review and meta-analysis. Jama 2013, 309, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Clinical Guidelines on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults--The Evidence Report. National Institutes of Health. Obes Res. 1998, 6, 464.
- Weir, C.B.; Jan, A. BMI Classification Percentile and Cut off Points; StatPearls: Treasure Island, FL, USA, 2019. [Google Scholar]
- Hardesty, C.K.; Poe-Kochert, C.; Son-Hing, J.P.; Thompson, G.H. Obesity negatively affects spinal surgery in idiopathic scoliosis. Clin. Orthop. Relat. Res. 2013, 471, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; Naghavi, M.; et al. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar]
- Rask-Andersen, M.; Karlsson, T.; Ek, W.E.; Johansson, Å. Genome-wide association study of body fat distribution identifies adiposity loci and sex-specific genetic effects. Nat. Commun. 2019, 10, 339. [Google Scholar] [CrossRef] [Green Version]
- WHO. World Health Organization Obesity and Overweight Fact Sheet. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 11 November 2022).
- World Health Organization. World Health Statistics 2021: Monitoring Health for the SDGs, Sustainable Development Goals; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Bodirsky, B.L.; Dietrich, J.P.; Martinelli, E.; Stenstad, A.; Pradhan, P.; Gabrysch, S.; Mishra, A.; Weindl, I.; Le Mouël, C.; Rolinski, S.; et al. The ongoing nutrition transition thwarts long-term targets for food security, public health and environmental protection. Sci. Rep. 2020, 10, 19778. [Google Scholar] [CrossRef]
- Verma, S.; Hussain, M.E. Obesity and diabetes: An update. Diabetes Metab. Syndr. Clin. Res. Rev. 2017, 11, 73–79. [Google Scholar] [CrossRef]
- Seravalle, G.; Grassi, G. Obesity and hypertension. Pharmacol. Res. 2017, 122, 1–7. [Google Scholar] [CrossRef]
- Alpert, M.A.; Lavie, C.J.; Agrawal, H.; Aggarwal, K.B.; Kumar, S.A. Obesity and heart failure: Epidemiology, pathophysiology, clinical manifestations, and management. Transl. Res. 2014, 164, 345–356. [Google Scholar] [CrossRef]
- Kernan, W.N.; Dearborn, J.L. Obesity increases stroke risk in young adults: Opportunity for prevention. Stroke 2015, 46, 1435–1436. [Google Scholar] [CrossRef] [Green Version]
- Calle, E.E.; Kaaks, R. Overweight, obesity and cancer: Epidemiological evidence and proposed mechanisms. Nat. Rev. Cancer 2004, 4, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, G.; Lewington, S.; Sherliker, P.; Clarke, R.; Emberson, J.; Halsey, J.; Qizilbash, N.; Collins, R.; Peto, R. Body-mass index and cause-specific mortality in 900,000 adults: Collaborative analyses of 57 prospective studies. Lancet 2009, 373, 1083–1096. [Google Scholar]
- Mazini, L.; Rochette, L.; Amine, M.; Malka, G. Regenerative Capacity of Adipose Derived Stem Cells (ADSCs), Comparison with Mesenchymal Stem Cells (MSCs). Int. J. Mol. Sci. 2019, 20, 2523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Fuentes, D.E.; Fernández-Garza, L.E.; Samia-Meza, J.A.; Barrera-Barrera, S.A.; Caplan, A.I.; Barrera-Saldaña, H.A. Mesenchymal stem cells current clinical applications: A systematic review. Arch. Med. Res. 2021, 52, 93–101. [Google Scholar] [CrossRef]
- Kozlowska, U.; Krawczenko, A.; Futoma, K.; Jurek, T.; Rorat, M.; Patrzalek, D.; Klimczak, A. Similarities and differences between mesenchymal stem/progenitor cells derived from various human tissues. World J. Stem Cells 2019, 11, 347–374. [Google Scholar] [CrossRef]
- Pal, B.; Das, B. In vitro Culture of Naïve Human Bone Marrow Mesenchymal Stem Cells: A Stemness Based Approach. Front. Cell Dev. Biol. 2017, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Orlic, D.; Kajstura, J.; Chimenti, S.; Bodine, D.M.; Leri, A.; Anversa, P. Bone marrow stem cells regenerate infarcted myocardium. Pediatr. Transplant. 2003, 7, 86–88. [Google Scholar] [CrossRef]
- Plotnikov, E.Y.; Khryapenkova, T.G.; Vasileva, A.K.; Marey, M.V.; Galkina, S.I.; Isaev, N.K.; Sheval, E.V.; Polyakov, V.Y.; Sukhikh, G.T.; Zorov, D.B. Cell-to-cell cross-talk between mesenchymal stem cells and cardiomyocytes in co-culture. J. Cell. Mol. Med. 2008, 12, 1622–1631. [Google Scholar] [CrossRef]
- Hu, J.; Yan, Q.; Shi, C.; Tian, Y.; Cao, P.; Yuan, W. BMSC paracrine activity attenuates interleukin-1β-induced inflammation and apoptosis in rat AF cells via inhibiting relative NF-κB signaling and the mitochondrial pathway. Am. J. Transl. Res. 2017, 9, 79–89. [Google Scholar]
- Asanuma, H.; Meldrum, D.R.; Meldrum, K.K. Therapeutic Applications of Mesenchymal Stem Cells to Repair Kidney Injury. J. Urol. 2010, 184, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Su, Y.P.; Kong, P.Y.; Zeng, D.F.; Chen, X.H.; Peng, X.G.; Zou, Z.M.; Xu, H. Human bone marrow mesenchymal stem cells expressing SDF-1 promote hematopoietic stem cell function of human mobilised peripheral blood CD34+ cells in vivo and in vitro. Int. J. Radiat. Biol. 2010, 86, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Cselenyák, A.; Pankotai, E.; Horváth, E.M.; Kiss, L.; Lacza, Z. Mesenchymal stem cells rescue cardiomyoblasts from cell death in an in vitro ischemia model via direct cell-to-cell connections. BMC Cell Biol. 2010, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sid-Otmane, C.; Perrault, L.P.; Ly, H.Q. Mesenchymal stem cell mediates cardiac repair through autocrine, paracrine and endocrine axes. J. Transl. Med. 2020, 18, 336. [Google Scholar] [CrossRef] [PubMed]
- Ulum, B.; Teker, H.T.; Sarikaya, A.; Balta, G.; Kuskonmaz, B.; Uckan-Cetinkaya, D.; Aerts-Kaya, F. Bone marrow mesenchymal stem cell donors with a high body mass index display elevated endoplasmic reticulum stress and are functionally impaired. J. Cell. Physiol. 2018, 233, 8429–8436. [Google Scholar] [CrossRef]
- Tencerova, M.; Frost, M.; Figeac, F.; Nielsen, T.K.; Ali, D.; Lauterlein, J.L.; Andersen, T.L.; Haakonsson, A.K.; Rauch, A.; Madsen, J.S.; et al. Obesity-Associated Hypermetabolism and Accelerated Senescence of Bone Marrow Stromal Stem Cells Suggest a Potential Mechanism for Bone Fragility. Cell Rep. 2019, 27, 2050–2062.e6. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.L.; Diekman, B.O.; Jain, D.; Guilak, F. Diet-induced obesity alters the differentiation potential of stem cells isolated from bone marrow, adipose tissue and infrapatellar fat pad: The effects of free fatty acids. Int. J. Obes. 2012, 37, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.J.; Gregoire, B.R.; Gao, H. High-fat diet decreases cancellous bone mass but has no effect on cortical bone mass in the tibia in mice. Bone 2009, 44, 1097–1104. [Google Scholar] [CrossRef]
- Picke, A.K.; Campbell, G.M.; Blüher, M.; Krügel, U.; Schmidt, F.N.; Tsourdi, E.; Winzer, M.; Rauner, M.; Vukicevic, V.; Busse, B.; et al. Thy-1 (CD90) promotes bone formation and protects against obesity. Sci. Transl. Med. 2018, 10, eaao6806. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.; Wu, L.; Cheng, P.; Yu, J.; Zhang, A.; Zha, J.; Liu, J.; Wang, L.; Di, W.; Hu, M.; et al. Correlation of obesity and osteoporosis: Effect of free fatty acids on bone marrow-derived mesenchymal stem cell differentiation. Exp. Ther. Med. 2010, 1, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Cortez, M.; Carmo, L.S.; Rogero, M.M.; Borelli, P.; Fock, R.A. A high-fat diet increases IL-1, IL-6, and TNF-α production by increasing NF-κB and attenuating PPAR-γ expression in bone marrow mesenchymal stem cells. Inflammation 2013, 36, 379–386. [Google Scholar] [CrossRef]
- Shu, L.; Beier, E.; Sheu, T.; Zhang, H.; Zuscik, M.J.; Puzas, E.J.; Boyce, B.F.; Mooney, R.A.; Xing, L. High-fat diet causes bone loss in young mice by promoting osteoclastogenesis through alteration of the bone marrow environment. Calcif. Tissue Int. 2015, 96, 313–323. [Google Scholar] [CrossRef] [Green Version]
- da Silva, S.V.; Renovato-Martins, M.; Ribeiro-Pereira, C.; Citelli, M.; Barja-Fidalgo, C. Obesity modifies bone marrow microenvironment and directs bone marrow mesenchymal cells to adipogenesis. Obesity 2016, 24, 2522–2532. [Google Scholar] [CrossRef]
- Gautam, J.; Khedgikar, V.; Kushwaha, P.; Choudhary, D.; Nagar, G.K.; Dev, K.; Dixit, P.; Singh, D.; Maurya, R.; Trivedi, R. Formononetin, an isoflavone, activates AMP-activated protein kinase/β-catenin signalling to inhibit adipogenesis and rescues C57BL/6 mice from high-fat diet-induced obesity and bone loss. Br. J. Nutr. 2017, 117, 645–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tencerova, M.; Figeac, F.; Ditzel, N.; Taipaleenmäki, H.; Nielsen, T.K.; Kassem, M. High-Fat Diet-Induced Obesity Promotes Expansion of Bone Marrow Adipose Tissue and Impairs Skeletal Stem Cell Functions in Mice. J. Bone Miner. Res. 2018, 33, 1154–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikary, S.; Kothari, P.; Choudhary, D.; Tripathi, A.K.; Trivedi, R. Glucocorticoid aggravates bone micro-architecture deterioration and skeletal muscle atrophy in mice fed on high-fat diet. Steroids 2019, 149, 108416. [Google Scholar] [CrossRef] [PubMed]
- Ayaz-Guner, S.; Alessio, N.; Acar, M.B.; Aprile, D.; Özcan, S.; Di Bernardo, G.; Peluso, G.; Galderisi, U. A comparative study on normal and obese mice indicates that the secretome of mesenchymal stromal cells is influenced by tissue environment and physiopathological conditions. Cell Commun. Signal. 2020, 18, 118. [Google Scholar] [CrossRef]
- Li, Y.; Lu, L.; Xie, Y.; Chen, X.; Tian, L.; Liang, Y.; Li, H.; Zhang, J.; Liu, Y.; Yu, X. Interleukin-6 Knockout Inhibits Senescence of Bone Mesenchymal Stem Cells in High-Fat Diet-Induced Bone Loss. Front. Endocrinol. 2021, 11, 622950. [Google Scholar] [CrossRef]
- Alessio, N.; Acar, M.B.; Demirsoy, I.H.; Squillaro, T.; Siniscalco, D.; Bernardo, G.D.; Peluso, G.; Özcan, S.; Galderisi, U. Obesity is associated with senescence of mesenchymal stromal cells derived from bone marrow, subcutaneous and visceral fat of young mice. Aging 2020, 12, 12609–12621. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, W.; Zhao, L.; Li, Y.; Liu, Z.; Gao, H.; Bai, X.; Wang, B. Obesity regulates miR-467/HoxA10 axis on osteogenic differentiation and fracture healing by BMSC-derived exosome LncRNA H19. J. Cell Mol. Med. 2021, 25, 1712–1724. [Google Scholar] [CrossRef]
- Bi, J.; Li, Q.; Yang, Z.; Cai, L.; Lv, T.; Yang, X.; Yan, L.; Liu, X.; Wang, Q.; Fu, X.; et al. CXCL2 Impairs Functions of Bone Marrow Mesenchymal Stem Cells and Can Serve as a Serum Marker in High-Fat Diet-Fed Rats. Front. Cell Dev. Biol. 2021, 9, 687942. [Google Scholar] [CrossRef]
- de Oliveira, G.P.; de Andrade, D.C.; Nascimento, A.L.R.; Cortez, E.; de Carvalho, S.N.; Stumbo, A.C.; Garcia-Souza, É.; Moura, A.S.; Carvalho, L.; Thole, A.A. Insulin-like growth factor-1 short-period therapy stimulates bone marrow cells in obese swiss mice. Cell Tissue Res. 2021, 384, 721–734. [Google Scholar] [CrossRef]
- Li, J.; Zhang, T.; Huang, C.; Xu, M.; Xie, W.; Pei, Q.; Xie, X.; Wang, B.; Li, X. Chemerin located in bone marrow promotes osteogenic differentiation and bone formation via Akt/Gsk3β/β- catenin axis in mice. J. Cell. Physiol. 2021, 236, 6042–6054. [Google Scholar] [CrossRef] [PubMed]
- Ali, D.; Figeac, F.; Caci, A.; Ditzel, N.; Schmal, C.; Kerckhofs, G.; Havelund, J.; Faergeman, N.; Rauch, A.; Tencerova, M.; et al. High-fat diet-induced obesity augments the deleterious effects of estrogen deficiency on bone: Evidence from ovariectomized mice. Aging Cell 2022, 21, e13726. [Google Scholar] [CrossRef] [PubMed]
- Benova, A.; Ferencakova, M.; Bardova, K.; Funda, J.; Prochazka, J.; Spoutil, F.; Cajka, T.; Dzubanova, M.; Balcaen, T.; Kerckhofs, G.; et al. Novel thiazolidinedione analog reduces a negative impact on bone and mesenchymal stem cell properties in obese mice compared to classical thiazolidinediones. Mol. Metab. 2022, 65, 101598. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Han, D.; Liu, T.; Huang, C.; Hu, Z.; Tan, X.; Wu, S. Asiatic acid improves high-fat-diet-induced osteoporosis in mice via regulating SIRT1/FOXO1 signaling and inhibiting oxidative stress. Histol. Histopathol. 2022, 37, 18446. [Google Scholar]
- Li, H.; Gou, Y.; Tian, F.; Lian, Q.; Hu, Y.; Zhang, L. The combined anti-osteoporotic effects of simvastatin and exercise in ovariectomized mice fed a high-fat diet. Exp. Gerontol. 2022, 164, 111794. [Google Scholar] [CrossRef]
- McCann, R.M.; Marsh, D.R.; Horner, A.; Clarke, S.A. Body mass index is more predictive of progenitor number in bone marrow stromal cell population than age in men: Expanding the predictors of the progenitor compartment. Tissue Eng. Part A 2010, 16, 889–896. [Google Scholar] [CrossRef]
- Di Bernardo, G.; Messina, G.; Capasso, S.; Del Gaudio, S.; Cipollaro, M.; Peluso, G.; Casale, F.; Monda, M.; Galderisi, U. Sera of overweight people promote in vitro adipocyte differentiation of bone marrow stromal cells. Stem Cell Res. Ther. 2014, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.T.; Phuong, T.N.T.; Tien, N.L.B.; Tran, D.K.; Thanh, V.V.; Quang, T.L.; Truong, D.T.; Pham, V.H.; Ngoc, V.T.N.; Chu-Dinh, T.; et al. An Update on the Progress of Isolation, Culture, Storage, and Clinical Application of Human Bone Marrow Mesenchymal Stem/Stromal Cells. Int. J. Mol. Sci. 2020, 21, 708. [Google Scholar] [CrossRef] [Green Version]
- Pochampally, R. Colony forming unit assays for MSCs. Methods Mol. Biol. 2008, 449, 83–91. [Google Scholar]
- Chaput, C.D.; Shar, A.; Jupiter, D.; Hubert, Z.; Clough, B.; Krause, U.; Gregory, C.A. How stem cell composition in bone marrow aspirate relates to clinical outcomes when used for cervical spine fusion. PLoS ONE 2018, 13, e0203714. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Contartese, D.; Giavaresi, G.; Sicuro, L.; Barbanti Brodano, G.; Gasbarrini, A.; Fini, M. A Rationale for the Use of Clotted Vertebral Bone Marrow to Aid Tissue Regeneration Following Spinal Surgery. Sci. Rep. 2020, 10, 4115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Guo, J.; Liu, F.; Wang, M.; Li, C.; Jia, L.; Zhai, L.; Wei, W.; Bai, Y. Depot-specific inflammation with decreased expression of ATM2 in white adipose tissues induced by high-margarine/lard intake. PLoS ONE 2017, 12, e0188007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surwit, R.S.; Kuhn, C.M.; Cochrane, C.; McCubbin, J.A.; Feinglos, M.N. Diet-induced type II diabetes in C57BL/6J mice. Diabetes 1988, 37, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.; Martin, T.L.; Surwit, R.S.; Robidoux, J. Genetic vulnerability to diet-induced obesity in the C57BL/6J mouse: Physiological and molecular characteristics. Physiol. Behav. 2004, 81, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, D.; Benham, V.; Bullard, B.; Kearney, T.; Hsia, H.C.; Gibbon, D.; Demireva, E.Y.; Lunt, S.Y.; Bernard, J.J. Fibroblast growth factor receptor is a mechanistic link between visceral adiposity and cancer. Oncogene 2017, 36, 6668–6679. [Google Scholar] [CrossRef] [Green Version]
- Hao, R.H.; Guo, Y.; Dong, S.S.; Weng, G.Z.; Yan, H.; Zhu, D.L.; Chen, X.F.; Chen, J.B.; Yang, T.L. Associations of Plasma FGF2 Levels and Polymorphisms in the FGF2 Gene with Obesity Phenotypes in Han Chinese Population. Sci. Rep. 2016, 6, 19868. [Google Scholar] [CrossRef] [Green Version]
- Kühn, M.C.; Willenberg, H.S.; Schott, M.; Papewalis, C.; Stumpf, U.; Flohé, S.; Scherbaum, W.A.; Schinner, S. Adipocyte-secreted factors increase osteoblast proliferation and the OPG/RANKL ratio to influence osteoclast formation. Mol. Cell Endocrinol. 2012, 349, 180–188. [Google Scholar] [CrossRef]
- Hankemeier, S.; Keus, M.; Zeichen, J.; Jagodzinski, M.; Barkhausen, T.; Bosch, U.; Krettek, C.; Van Griensven, M. Modulation of proliferation and differentiation of human bone marrow stromal cells by fibroblast growth factor 2: Potential implications for tissue engineering of tendons and ligaments. Tissue Eng. 2005, 11, 41–49. [Google Scholar] [CrossRef]
- Zhu, Z.; Gan, X.; Yu, H. NF-κB-miR15a-bFGF/VEGFA axis contributes to the impaired angiogenic capacity of BM-MSCs in high fat diet-fed mice. Mol. Med. Rep. 2017, 16, 7609–7616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, I.; Muraglia, A.; Campanile, G.; Cancedda, R.; Quarto, R. Fibroblast growth factor-2 supports ex vivo expansion and maintenance of osteogenic precursors from human bone marrow. Endocrinology 1997, 138, 4456–4462. [Google Scholar] [CrossRef]
- Callegari, A.J.; Kelly, T.J. Shedding Light on the DNA Damage Checkpoint. Cell Cycle 2007, 6, 660–666. [Google Scholar] [CrossRef]
- Lv, F.-J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. Concise review: The surface markers and identity of human mesenchymal stem cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Ambrosi, T.H.; Scialdone, A.; Graja, A.; Gohlke, S.; Jank, A.M.; Bocian, C.; Woelk, L.; Fan, H.; Logan, D.W.; Schürmann, A.; et al. Adipocyte Accumulation in the Bone Marrow during Obesity and Aging Impairs Stem Cell-Based Hematopoietic and Bone Regeneration. Cell Stem Cell 2017, 20, 771–784.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, R.; Zhou, B.O.; Shimada, I.S.; Zhao, Z.; Morrison, S.J. Leptin receptor promotes adipogenesis and reduces osteogenesis by regulating mesenchymal stromal cells in adult bone marrow. Cell Stem Cell 2016, 18, 782–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.X.; Zhang, N.; Wang, H.W.; Gao, P.; Yang, Q.P.; Wen, Q.P. CXCR4 receptor overexpression in mesenchymal stem cells facilitates treatment of acute lung injury in rats. J. Biol. Chem. 2015, 290, 1994–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Mohammad, K.; Pelus, L. CXCR4 expression in the bone marrow microenvironment is required for hematopoietic stem and progenitor cell maintenance and early hematopoietic regeneration after myeloablation. Stem Cells 2020, 38, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Chang, C.Y.; Lin, B.Z.; Wu, Y.L.; Wu, M.H.; Lin, L.T.; Huang, W.C.; Holz, J.D.; Sheu, T.J.; Lee, J.S.; et al. Up-regulation of cofilin-1 in cell senescence associates with morphological change and p27(kip1) -mediated growth delay. Aging Cell 2021, 20, e13288. [Google Scholar] [CrossRef]
- Liu, J.; Ding, Y.; Liu, Z.; Liang, X. Senescence in Mesenchymal Stem Cells: Functional Alterations, Molecular Mechanisms, and Rejuvenation Strategies. Front. Cell Dev. Biol. 2020, 8, 258. [Google Scholar] [CrossRef]
- Severino, V.; Alessio, N.; Farina, A.; Sandomenico, A.; Cipollaro, M.; Peluso, G.; Galderisi, U.; Chambery, A. Insulin-like growth factor binding proteins 4 and 7 released by senescent cells promote premature senescence in mesenchymal stem cells. Cell Death Dis. 2013, 4, e911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Guo, Y.; Zhai, H.; Yin, Y.; Zhang, J.; Chen, H.; Wang, L.; Li, N.; Liu, R.; Xia, Y. Aging increases the susceptivity of MSCs to reactive oxygen species and impairs their therapeutic potency for myocardial infarction. PLoS ONE 2014, 9, e111850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Stroncek, D.F.; Zhao, Y.; Jin, P.; Castiello, L.; Civini, S.; Wang, H.; Feng, J.; Tran, K.; Kuznetsov, S.A.; et al. Intra-subject variability in human bone marrow stromal cell (BMSC) replicative senescence: Molecular changes associated with BMSC senescence. Stem Cell Res. 2013, 11, 1060–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xu, X.; Wang, L.; Liu, G.; Li, Y.; Wu, X.; Jing, Y.; Li, H.; Wang, G. Senescent mesenchymal stem cells promote colorectal cancer cells growth via galectin-3 expression. Cell Biosci. 2015, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fafián-Labora, J.A.; Morente-López, M.; Arufe, M.C. Effect of aging on behaviour of mesenchymal stem cells. World J. Stem Cells 2019, 11, 337–346. [Google Scholar] [CrossRef]
- Passos, J.F.; Nelson, G.; Wang, C.; Richter, T.; Simillion, C.; Proctor, C.J.; Miwa, S.; Olijslagers, S.; Hallinan, J.; Wipat, A.; et al. Feedback between p21 and reactive oxygen production is necessary for cell senescence. Mol. Syst. Biol. 2010, 6, 347. [Google Scholar] [CrossRef]
- Shao, L.; Li, H.; Pazhanisamy, S.K.; Meng, A.; Wang, Y.; Zhou, D. Reactive oxygen species and hematopoietic stem cell senescence. Int. J. Hematol. 2011, 94, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Mosteiro, L.; Pantoja, C.; de Martino, A.; Serrano, M. Senescence promotes in vivo reprogramming through p16(INK)(4a) and IL-6. Aging Cell 2018, 17, e12711. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Q.; Wang, Y.; Li, L.; Bu, H.; Bao, J. Senescence of mesenchymal stem cells (Review). Int. J. Mol. Med. 2017, 39, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Farr, J.N.; Fraser, D.G.; Wang, H.; Jaehn, K.; Ogrodnik, M.B.; Weivoda, M.M.; Drake, M.T.; Tchkonia, T.; LeBrasseur, N.K.; Kirkland, J.L.; et al. Identification of Senescent Cells in the Bone Microenvironment. J. Bone Miner. Res. 2016, 31, 1920–1929. [Google Scholar] [CrossRef] [Green Version]
- Farr, J.N.; Xu, M.; Weivoda, M.M.; Monroe, D.G.; Fraser, D.G.; Onken, J.L.; Negley, B.A.; Sfeir, J.G.; Ogrodnik, M.B.; Hachfeld, C.M.; et al. Targeting cellular senescence prevents age-related bone loss in mice. Nat. Med. 2017, 23, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senichkin, V.V.; Pervushin, N.V.; Zamaraev, A.V.; Sazonova, E.V.; Zuev, A.P.; Streletskaia, A.Y.; Prikazchikova, T.A.; Zatsepin, T.S.; Kovaleva, O.V.; Tchevkina, E.M.; et al. Bak and Bcl-xL Participate in Regulating Sensitivity of Solid Tumor Derived Cell Lines to Mcl-1 Inhibitors. Cancers 2021, 14, 181. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Canfield, K.; Feng, W.; Kurokawa, M. Metabolic Regulation of Apoptosis in Cancer. Int. Rev. Cell Mol. Biol. 2016, 327, 43–87. [Google Scholar] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; She, P.; Mozzoli, M.; Cheung, P.; Gumireddy, K.; Reddy, P.; Xiang, X.; Luo, Z.; Ruderman, N. Free fatty acids produce insulin resistance and activate the proinflammatory nuclear factor-kappaB pathway in rat liver. Diabetes 2005, 54, 3458–3465. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Author | Species | Gender | Age | Diet/Duration | BMSC Origin | Main Conclusion(s) |
|---|---|---|---|---|---|---|
| Cao et al. [31] | C57BL/6 mice | Male | 6 weeks + 3 days | A control purified diet (10% energy as fat, D12450B, Research Diet, Inc., New Brunswick, NJ, United States) based on AIN-93G or an HFD (45% energy as fat) with extra fat from lard for 14 weeks | Tibias and femurs | Increased osteogenic potential of obese BMSCs |
| Lv et al. [33] | C57BL/6 mice | Male | 2 weeks | A normal chow or an HFD for 24 weeks | Tibias and femurs | Enhanced osteogenesis and inhibited adipogenesis in obese BMSCs, which might be associated with factors secreted by FFA-treated adipocytes |
| Cortez et al. [34] | Wistar rats | Male | 2 months + 10 days | A control diet (AIN-93M diet; total energy: 75.8% carbohydrates, 9.3% fat, and 14.9% protein) or an HFD (an AIN-93M-based diet that was enriched with lard; total energy: 24.2% carbohydrates, 60.9% fat, and 14.9% protein) for 12 weeks | Femurs | Increased inflammatory cytokine (i.e., IL-1, IL-6, and TNF-α) secretion, upregulated NF-κB and RUNX2 expression, and reduced PPARγ expression in obese BMSCs |
| Wu et al. [30] | C57BL/6J mice | Male | — | A low-fat diet (D12450B, 10% energy from fat, Research Diets, Inc.) or an HFD (D12492, 60% energy from fat, Research Diets, Inc.) for 14 weeks | Tibias and femurs | No significant change in proliferation; reduced trilineage differentiation, decreased CD105, and increased PDGFRα expression in obese BMSCs; FFAs increase adipogenic and osteogenic differentiation and have no impact on chondrogenic differentiation of BMSCs |
| Shu et al. [35] | C57BL/6J mice | Male | 5 weeks | A low-fat diet (10% kcal, D12450B, Research Diets, Inc.) or an HFD (60%kcal, D12492, Research Diets, Inc.) for 12 weeks | Tibias and femurs | No significant difference in clonogenicity between obese and lean BMSCs; increased adipogenic and osteogenic differentiation abilities of obese BMSCs; increased SOX9 mRNA level in obese BMSCs |
| da Silva et al. [36] | C57BL/6 mice | Male | 4 weeks | A regular chow (396 kcal/100g, 13% of energy derived from fat) or an HFD (470 kcal/100 g, 45% of energy derived from fat) for 10 weeks | Tibias and femurs | Obesity increases the proliferation of BMSCs and commits these cells to adipogenesis; similar expressions of BMSC markers between lean and obese mice |
| Gautam et al. [37] | C57BL/6 mice | Male | 4 weeks | A standard chow diet or an HFD (D12109, 40% kcal% fat; Research Diets, Inc.) or an HFD (D12109, 40% kcal% fat; Research Diets, Inc.) and formononetin (0.1,1,10 mg/kg per day) for 12 weeks | Femurs | Formononetin improves osteogenic differentiation and inhibits adipogenic differentiation of obese BMSCs |
| Picke et al. [32] | C57BL/6J mice | Male | 4–5 weeks | An HFD (EF R/M D12331 diet modified by Surwit, ssniff) or a chow diet for 12 to 20 weeks Note: CD90 knock-out mice on C57BL/6J background and wild-type C57BL/6J mice for functional validation | Tibias and femurs | The similar clonogenic ability between lean and obese BMSCs; reduced CD90 expression in obese BMSCs; CD90-deficient BMSCs have increased adipogenic differentiation, attenuated osteogenic differentiation, and a decreasing trend in apoptosis |
| Tencerova et al. [38] | C57BL/6J mice | Male | 8 weeks | A normal chow diet (Altromin, Lage, Germany, 132003, containing 6% fat, 30% protein, 63% carbohydrate, 7.7% sucrose) or a 60 kcal% high-fat diet (Research Diet, Inc., D12492, containing 35% fat, 26% protein, 26% carbohydrate, 8.8% sucrose) for 12 weeks | The bones of the front and hind limbs | Inhibited proliferation and colony-forming potential in obese BMSCs; downregulated expressions of CD73+ and Sca1/CD140a+ and upregulated adipogenesis-associated genes in obese BMSCs; no significant change in senescent phenotype |
| Adhikary et al. [39] | BALB/c mice | Male | 9 weeks | A standard chow diet or an HFD (D12492, 60% energy from fat; Research Diets, Inc.) for 10 weeks | Femurs | Impaired mineralization ability and increased adipogenic differentiation in obese BMSCs |
| Ayaz-Guner et al. [40] | C57BL/6 mice | Male | 3 weeks | A normal diet (containing 10% fat, 70% carbohydrates, and 20% protein (total 3.82 kcal/g)) or an HFD (containing 60% fat from lard, 20% carbohydrates, and 20% protein (total 5.21 kcal/g), Research Diets, Inc.) for 10 weeks | Tibias and femurs | Comparable expressions of CD105, CD90, and CD73 between normal and obese BMSCs |
| Li et al. [41] | Wild-type and IL-6-deficient mice generated on C57BL/6 background | Male | 8 weeks | A standard diet (19.2% protein, 67.3% carbohydrate, 4.3% fat) or an HFD (26% protein, 26% carbohydrate, 35% fat) for 12 weeks | Tibias and femurs | Obesity attenuates the clonogenic ability and induces senescence of BMSCs, which can be ameliorated by IL-6 knockout |
| Alessio et al. [42] | C57BL/6 mice | Male | 3 weeks | A normal diet (10% fat, 70% carbohydrate, and 20% protein (total 3.82 kcal/g)) or an HFD (containing 60% fat from lard, 20% carbohydrate, and 20% protein (total 5.21 kcal/g), Research Diets, Inc.) for 10 weeks | Tibias and femurs | Suppressed proliferation, increased senescence, unchanged apoptosis, clonogenicity and trilineage differentiation in obese BMSCs |
| Wang et al. [43] | C57BL/6J mice | Male | 7 weeks | A normal diet or an HFD | Tibias and femurs | Obesity reduces the secretion of BMSC-derived exosomes and the level of carried LncRNA H19, thereby affecting the miR-467/HoxA10 axis and ultimately inhibiting the osteogenic process |
| Bi et al. [44] | Sprague-Dawley rats | Female | 6 weeks | A normal diet (GB14924.3-2001, 100% basal diet) and a customized HFD (containing 60% basal diet, 10% sucrose, 20% lard, and 10% egg yolk powder) for 4 months and 6 months | Tibias and femurs | Inhibited proliferation, unchanged apoptosis, reduced adipogenic and osteogenic differentiation, increased senescence in obese BMSCs; CXCL2 impairs adipogenesis and promotes senescence of BMSCs |
| de Oliveira et al. [45] | Swiss mice | Male | 21 days | A standard AIN93G diet (65.6% carbohydrates, 17.3% proteins, and 17.1% lipids) or a Western diet rich in saturated fat and simple carbohydrate from clarified butter (Ghee) (43.3% carbohydrates, 14% proteins, and 42.7% lipids) (PragSoluções, Brazil) for 12 weeks Note: After 11 weeks, half of the obese mice received IGF-1 treatment. | Tibias and femurs | Obesity has a pro-apoptotic effect on BMSCs, which can be improved by IGF-1 |
| Li et al. [46] | C57BL/6 mice | Male | 8 weeks | A normal chow or an HFD (60% kcal in fat) for 12 weeks Note: Chemerin knock-out (Rarres2−/−) and Ap2−drive Rarres2 overexpression transgenic mice for functional validation | Tibias and femurs | Decreased chemerin in the obese bone marrow is associated with the suppressed osteogenesis of BMSCs |
| Ali et al. [47] | C57BL/6J mice and OVX C57BL/6J mice | Female | 8 weeks | A R-70 normal diet (Lantmännen, containing kcal%: protein 14.5%, carbohydrates 60%, and fat 4.5%) in which fat content was from oatmeal, barley, wheat bran, wheat flour or an HFD (Research Diet, D12492 containing kcal%: protein 20%, carbohydrates 20%, and fat 60%) in which fat content was from soybean oil and lard for 12 weeks | The bones of the front and hind limbs | Obesity upregulates senescence-related genes and downregulates osteogenic genes in BMSCs of OVX mice |
| Benova et al. [48] | C57BL/6N mice | Male | 12 weeks | A normal diet (3.4% wt/wt as lipids) or an HFD (lipid content, ~35% wt/wt, mainly corn oil) or an HFD and pioglitazone (50 mg pioglitazone/kg diet (Actos, Takeda, Japan)) or a HFD+MSDC-0602K (330 mg MSDC-0602K/kg diet (Cirius Therapeutics, USA)) for 8 weeks | The bones of the front and hind limbs | No significant difference in proliferation and colony-forming ability between lean and obese BMSCs; MSDC-0602K increases osteogenesis and reduces adipogenesis of obese BMSCs |
| Chen et al. [49] | C57BL/6 mice | Male | 4 weeks | A normal control diet or an HFD (carbohydrates, 20.3%; fat, 61.6%; protein, 18.1%) or an HFD with 25 mg/kg asiatic acid or an HFD with 50 mg/kg asiatic acid. | Femurs | Asiatic acid ameliorates inhibited osteogenesis and suppresses enhanced adipogenesis of obese BMSCs |
| Li et al. [50] | C57BL/6 mice and OVX C57BL/6 mice | Female | 3 months | A standard diet or an HFD for 8 weeks | Tibias and femurs | Obesity has no significant effect on the osteogenic ability of BMSCs from normal and OVX female mice |
| Author | Gender | Age | BMI | BMSC Origin | Main Conclusion(s) |
|---|---|---|---|---|---|
| McCann et al. [51] | Male and Female | 28–91 years (mean age 64 years and median age 63.5 years) | Male: mean BMI: 29.6 kg/m2 median BMI: 29.3 kg/m2 Female: mean BMI: 28.1 kg/m2 median BMI: 27.4 kg/m2 | Femoral canal | BMI is positively correlated with colony area and number in males but not females; the percentage of the number and area of CFU-ALP+ has no relationship with BMI, irrespective of females or males |
| Di Bernardo et al. [52] | Male | BMSCs from 10-year-old, 12-year-old, and 13-year-old male donors; Sera from healthy and overweight adult males | Healthy weight: 21.10 ± 1.10 kg/m2 Overweight: 29.63 ± 1.80 kg/m2 | — | Overweight (BMI > 25 kg/m2) serum does not influence the proliferation, senescence, and apoptosis of hBMSCs; overweight (BMI > 25 kg/m2) serum increases adipogenesis and impairs osteogenesis of hBMSCs |
| Ulum et al. [28] | — | Normal BMI: 8–36 years High BMI: 14–58 years | Normal BMI: 17.62 ± 4.27 kg/m2 High BMI: 30.85 ± 4.16 kg/m2 | — | High-BMI (BMI > 30 kg/m2) hBMSCs have several altered expressions of surface antigens, decreased proliferation, increased senescence, impaired osteogenesis, and a relative defect in adipogenesis; the abnormal osteogenesis is associated with elevated ERS and impaired unfolded protein response; TUDCA and 4-PBA can partially rescue the osteogenic ability and facilitate adipogenesis of high-BMI (BMI > 30 kg/m2) hBMSCs |
| Tencerova et al. [29] | Male | Lean: 31 ± 3 years Overweight: 32 ± 3 years Obese: 37 ± 2 years | Lean: 22.9 ± 0.3 kg/m2 Overweight: 28.0 ± 0.4 kg/m2 Obese: 36.1 ± 0.8 kg/m2 | Iliac crest | Obese hBMSCs inhibited proliferation and clonogenicity; enhanced in vitro adipogenic and osteogenic differentiation; increased number of LEPR+, IR+, and senescent cells; increased commitment to adipogenesis Overweight hBMSCs decreased proliferation; unchanged clonogenicity, in vitro adipogenic and osteogenic differentiation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zong, Q.; Bundkirchen, K.; Neunaber, C.; Noack, S. Are the Properties of Bone Marrow-Derived Mesenchymal Stem Cells Influenced by Overweight and Obesity? Int. J. Mol. Sci. 2023, 24, 4831. https://doi.org/10.3390/ijms24054831
Zong Q, Bundkirchen K, Neunaber C, Noack S. Are the Properties of Bone Marrow-Derived Mesenchymal Stem Cells Influenced by Overweight and Obesity? International Journal of Molecular Sciences. 2023; 24(5):4831. https://doi.org/10.3390/ijms24054831
Chicago/Turabian StyleZong, Qiang, Katrin Bundkirchen, Claudia Neunaber, and Sandra Noack. 2023. "Are the Properties of Bone Marrow-Derived Mesenchymal Stem Cells Influenced by Overweight and Obesity?" International Journal of Molecular Sciences 24, no. 5: 4831. https://doi.org/10.3390/ijms24054831