
Otilonium Bromide Prevents Cholinergic Changes in the Distal Colon Induced by Chronic Water Avoidance Stress, a Rat Model of Irritable Bowel Syndrome

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Body Weight Gain, Water Intake, and Fecal Pellet Production
2.2. Histochemistry
Periodic Acid and Schiff’s Reagent (PAS) and Toluidine Blue Staining
2.3. Functional Studies
2.3.1. Contractile Responses Elicited by Electrical Field Stimulation (EFS) in Strips from the Different Animal Groups
2.3.2. Direct Smooth Muscle Contractions Elicited by Methacholine in Strips from the Different Animal Groups
2.4. Immunohistochemistry
2.4.1. Protein Gene Product 9.5 (PGR9.5) Immunoreactivity (IR)
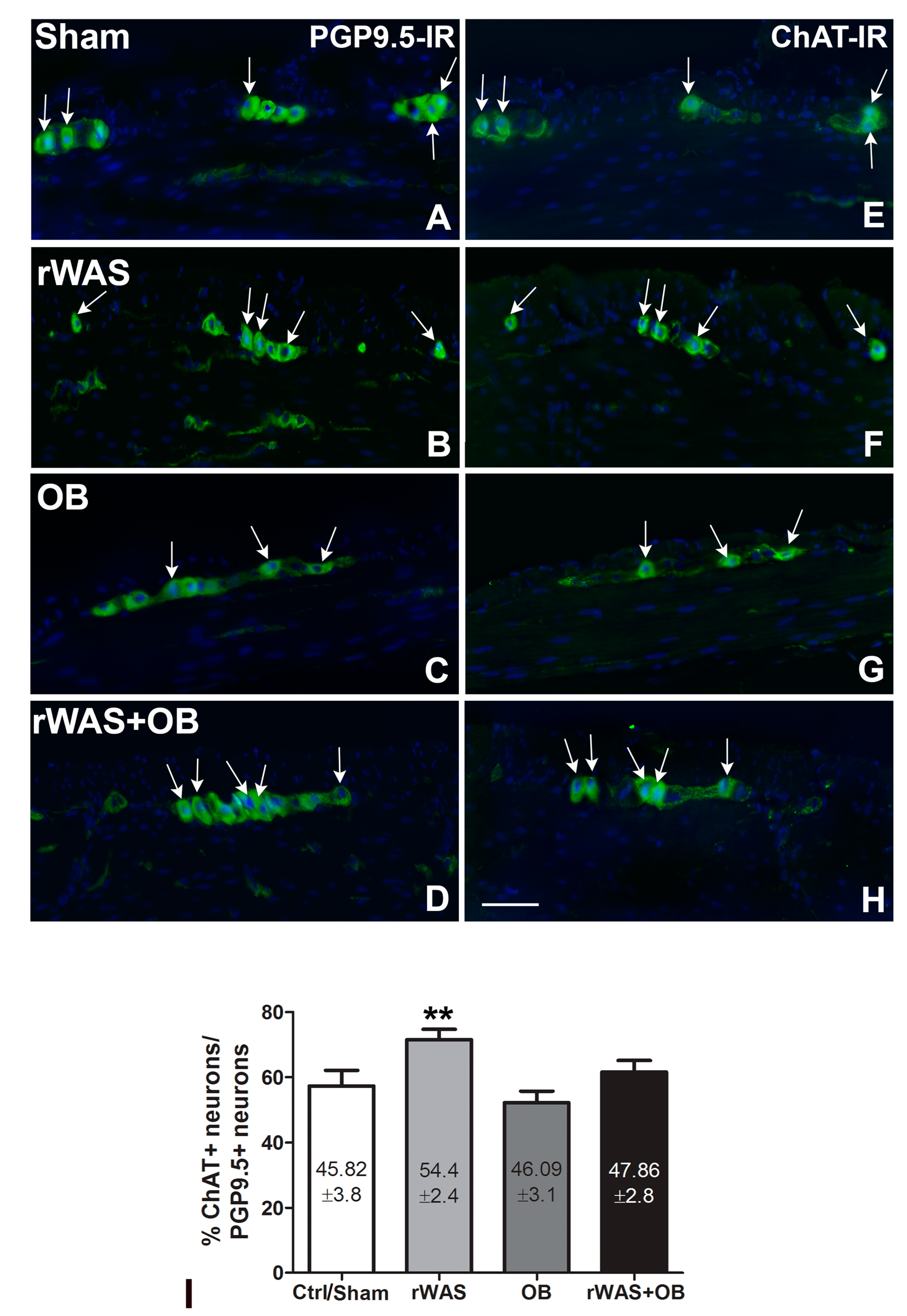
2.4.2. Choline Acetyl Transferase (ChAT)-IR
3. Discussion
4. Materials and Methods
4.1. Animals
- (1)
- Group of control rats who were not exposed to stress or to pharmacological treatment (Ctrl; n = 4);
- (2)
- Group of rats who were subjected to repeated water avoidance stress (rWAS; n = 7) for 10 consecutive days;
- (3)
- Group of rats who experienced the stress environment without being subjected to it (Sham; n = 5);
- (4)
- Group of rats who were subjected to rWAS for 10 consecutive days and, meanwhile, they were orally treated with OB (rWAS+OB; n = 7);
- (5)
- Group of rats who were treated orally with OB for 10 days and not subjected to stress (OB; n = 7).
4.2. Repeated Water Avoidance Stress (rWAS) Protocol
4.3. OB Preparation and Administration
4.4. Tissue Sampling
4.5. Functional Experiments
4.6. Morphological Studies
4.7. Quantitative and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mayer, E.A.; Ryu, H.J.; Bhatt, R.R. The neurobiology of irritable bowel syndrome. Mol. Psychiatry 2023. [Google Scholar] [CrossRef]
- Drossman, D.A. Functional gastrointestinal disorders: History, pathophysiology, clinical features, and Rome IV. Gastroenterology 2016, 150, 1262–1279. [Google Scholar] [CrossRef] [PubMed]
- Fond, G.; Loundou, A.; Hamdani, N.; Boukouaci, W.; Dargel, J.O.; Roger, M.; Tamouza, R.; Leboyer, M.; Boyer, L. Anxiety and depression comorbidities in irritable bowel syndrome (IBS): A systematic review and meta-analysis. Eur. Arch. Psychiatry Clin. Neurosci. 2014, 264, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Moloney, R.D.; O’Mahony, S.M.; Dinan, T.G.; Cryan, J.F. Stress-induced visceral pain: Toward animal models of irritable-bowel syndrome and associated comorbidities. Front. Psychiatry 2015, 16, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G.; Evangelista, S. Experimental model of irritable bowel syndrome and the role of enteric neurotransmission. J. Clin. Med. 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Simrén, M.; Törnblom, H.; Palsson, O.S.; Van Oudenhove, L.; Whitehead, W.E.; Tack, J. Cumulative effects of psychologic distress, visceral hypersensitivity, and abnormal transit on patient-reported outcomes in irritable bowel Syndrome. Gastroenterology 2019, 157, 391–402.e2. [Google Scholar] [CrossRef] [PubMed]
- Larauche, M.; Mulak, A.; Taché, Y. Stress and visceral pain: From animal models to clinical therapies. Exp. Neurol. 2012, 233, 49–67. [Google Scholar] [CrossRef]
- Surdea-Blaga, T.; Băban, A.; Dumitrascu, D.L. Psychosocial determinants of irritable bowel syndrome. World J. Gastroenterol. 2012, 18, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, K.; Nawaz, H.; Abid, S. Functional gastrointestinal disorders and gut-brain axis: What does the future hold? World J. Gastroenterol. 2019, 25, 552–566. [Google Scholar] [CrossRef]
- Bercik, P. The Brain-Gut-Microbiome Axis and Irritable Bowel Syndrome. Gastroenterol. Hepatol. 2020, 16, 322–324. [Google Scholar]
- Moloney, R.D.; Desbonnet, L.; Clarke, G.; Dinan, T.G.; Cryan, J.F. The microbiome: Stress, health and disease. Mamm. Genome 2014, 25, 49–74. [Google Scholar] [CrossRef]
- Zhuang, X.; Xiong, L.; Li, L.; Li, M.; Chen, M. Alterations of gut microbiota in patients with irritable bowel syndrome: A systematic review and meta-analysis. J. Gastroenterol. Hepatol. 2017, 32, 28–38. [Google Scholar] [CrossRef]
- Bradesi, S.; Schwetz, I.; Ennes, H.S.; Lamy, C.M.R.; Ohning, G.; Fanselow, M.; Charalabos Pothoulakis, C.; McRoberts, J.A.; Mayer, E.M. Repeated exposure to water avoidance stress in rats: A new model for sustained visceral hyperalgesia. Am J. Physiol. Gatrointest. Liver Physiol. 2005, 289, G42–G53. [Google Scholar] [CrossRef]
- Larauche, M.; Mulak, A.; Taché, Y. Stress-related alterations of visceral sensation: Animal models for irritable bowel syndrome study. J. Neurogastroenterol. Motil 2011, 17, 213–234. [Google Scholar] [CrossRef] [PubMed]
- Traini, C.; Idrizaj, E.; Garella, R.; Faussone-Pellegrini, M.S.; Baccari, M.C.; Vannucchi, M.G. Otilonium Bromide treatment prevents nitrergic functional and morphological changes caused by chronic stress in the distal colon of a rat IBS model. J. Cell Mol. Med. 2021, 25, 6988–7000. [Google Scholar] [CrossRef]
- Balestra, B.; Vicini, R.; Cremon, C.; Zecchi, L.; Dothel, G.; Vasina, V.; De Giorgio, R.; Paccapelo, A.; Pastoris, O.; Stanghellini, V.; et al. Colonic mucosal mediators from patients with irritable bowel syndrome excite enteric cholinergic motor neurons. Neurogastroenterol. Motil. 2012, 24, 1118-e570. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.J.; Yu, B.P. Upregulation of the high-affinity choline transporter in colon relieves stress-induced hyperalgesia. Pain Res. 2018, 1, 1971–1982. [Google Scholar] [CrossRef] [PubMed]
- Triantafillidis, J.K.; Malgarinos, G. Long-term efficacy and safety of otilonium bromide in the management of irritable bowel syndrome: A literature review. Clin. Exp. Gastroenterol. 2014, 7, 75–82. [Google Scholar] [CrossRef]
- Cunningham, A.L.; Esarte Palomero, O.; Voss, B.J.; Trent, M.S.; Davies, B.W. Irritable Bowel Syndrome Therapeutic Has Broad-Spectrum Antimicrobial Activity. Antimicrob. Agents Chemother. 2021, 65, e0044321. [Google Scholar] [CrossRef]
- Cipriani, G.; Gibbons, S.J.; Saravanaperumal, S.A.; Malysz, J.; Sha, L.; Szurszewski, J.H.; Linden, D.R.; Evangelista, S.; Faussone-pellegrini, M.S.; Vannucchi, M.G.; et al. Changes in nitrergic and tachykininergic pathways in rat proximal colon in response to chronic treatment with otilonium bromide. Neurogastroenterol. Motil. 2015, 27, 997–1009. [Google Scholar] [CrossRef]
- Traini, C.; Cipriani, G.; Evangelista, S.; Santicioli, P.; Faussone-Pellegrini, M.S.; Vannucchi, M.G. Chronic treatment with otilonium bromide induces changes in L-type Ca2+ channel, tachykinins, and nitric oxide synthase expression in rat colon muscle coat. Neurogastroent. Motil. 2013, 25, e728–e739. [Google Scholar] [CrossRef]
- Traini, C.; Evangelista, S.; Girod, V.; Faussone-Pellegrini, M.S.; Vannucchi, M.G. Repeated otilonium bromide administration prevents neurotransmitter changes in colon of rats underwent to wrap restraint stress. J. Cell Mol. Med. 2017, 21, 735–745. [Google Scholar] [CrossRef]
- Evangelista, S.; Traini, C.; Vannucchi, M.G. Otilonium Bromide: A drug with a complex mechanism of action. Curr. Pharm. Des. 2018, 24, 1772–1779. [Google Scholar] [CrossRef]
- Zhou, L.; She, P.; Tan, F.; Zeng, L.S.; Chen, L.; Luo, Z.; Wu, Y. Repurposing Antispasmodic Agent Otilonium Bromide for Treatment of Staphylococcus aureus. Infect. Front. Microbiol. 2020, 11, 1720. [Google Scholar] [CrossRef]
- Mayer, E.A.; Collins, S.M. Evolving pathophysiologic models of functional gastrointestinal disorders. Gastroenterology 2002, 122, 2032–2048. [Google Scholar] [CrossRef]
- Sicard, J.F.; Le Bihan, G.; Vogeleer, P.; Jacques, M.; Harel, J. Interactions of Intestinal Bacteria with Components of the Intestinal Mucus. Front. Cell Infect. Microbiol. 2017, 7, 387. [Google Scholar] [CrossRef] [PubMed]
- Juge, N. Relationship between mucosa-associated gut microbiota and human diseases. Biochem. Soc. Trans. 2022, 50, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Simrén, M.; Barbara, G.; Flint, H.J.; Spiegel, B.M.R.; Spiller, R.C.; Vanner, S.; Verdu, E.F.; Whorwell, P.J.; Zoetendal, E.G. Rome Foundation Committee. Intestinal microbiota in functional bowel disorders: A Rome foundation report. Gut 2013, 62, 159–176. [Google Scholar] [CrossRef]
- Chong, P.P.; Chin, V.K.; Looi, C.Y.; Wong, W.F.; Madhavan, P.; Yong, V.C. The Microbiome and Irritable Bowel Syndrome. A Review on the Pathophysiology, Current Research and Future Therapy. Front. Microbiol. 2019, 10, 1–23. [Google Scholar] [CrossRef]
- Lindqvist, S.; Hernon, J.; Sharp, P.; Johns, N.; Addison, S.; Watson, M.; Tighe, R.; Greer, S.; Mackay, J.; Rhodes, m.; et al. The colon-selective spasmolytic otilonium bromide inhibits muscarinic M3 receptor-coupled calcium signals in isolated human colonic crypts. Br. J. Pharmacol. 2002, 137, 1134–1142. [Google Scholar] [CrossRef]
- Hirota, C.L.; McKay, D.M. Cholinergic regulation of epithelial ion transport in the mammalian intestine. Br. J. Pharmacol. 2006, 149, 463–479. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.C.; Li, G.W.; Wang, T.T.; Gao, L.; Wang, F.F.; Shang, H.W.; Yang, Z.J.; Guo, Y.X.; Wang, B.Y.; Xu, J.D. Rhubarb extract relieves constipation by stimulating mucus production in the colon and altering the intestinal flora. Biomed. Pharmacother. 2021, 138, 111479. [Google Scholar] [CrossRef]
- Gourcerol, G.; Wu, S.V.; Yuan, P.Q.; Pham, H.; Miampamba, M.; Larauche, M.; Sanders, P.; Amano, T.; Mulak, A.; Im, E.; et al. Activation of corticotropin-releasing factor receptor 2 mediates the colonic motor coping response to acute stress in rodents. Gastroenterology 2011, 140, 1586–1596.e6. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.J.; Kim, K.B.; Yoon, S.M.; Han, J.H.; Chae, H.B.; Park, S.M.; Youn, S.J. Corticotropin-releasing factor stimulates colonic motility via muscarinic receptors in the rat. World J. Gastroenterol. 2017, 23, 3825–3831. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L.; Vale, W.W. CRF and CRF receptors: Role in stress responsivity and other behaviors. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 525–557. [Google Scholar] [CrossRef]
- Buckley, M.M.; O’Halloran, K.D.; Rae, M.G.; Dinan, T.G.; O’Malley, D. Modulation of enteric neurons by interleukin-6 and corticotropin releasing factor contributes to visceral hypersensitivity and altered colonic motility in a rat model of irritable bowel syndrome. J. Physiol. 2014, 592, 5235–5250. [Google Scholar] [CrossRef]
- Maillot, C.; Million, M.; Wei, J.Y.; Gauthier, A.; Taché, Y. Peripheral corticotropin-releasing factor and stress-stimulated colonic motor activity involve type 1 receptor in rats. Gastroenterology 2000, 119, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Larauche, M.; Gourcerol, G.; Wang, L.; Pambukchian, K.; Brunnhuber, S.; Adelson, D.W.; Rivier, J.; Million, M.; Taché, Y. Cortagine, a CRF1 agonist, induces stresslike alterations of colonic function and visceral hypersensitivity in rodents primarily through peripheral pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G215–G227. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.Q.; Million, M.; Wu, S.V.; Rivier, J.; Tachè, Y. Peripheral corticotropin releasing factor (CRF) and a novel CRF1 receptor agonist, stressin1-A activate CRF1 receptor expressing cholinergic and nitrergic myenteric neurons selectively in the colon of conscious rats. Neurogastroenterol. Motil. 2007, 19, 923–936. [Google Scholar] [CrossRef]
- Liu, S.; Ren, W.; Qu, M.H.; Bishop, G.A.; Wang, G.D.; Wang, X.Y.; Xia, Y.; Wood, J.D. Differential actions of urocortins on neurons of the myenteric division of the enteric nervous system in guinea pig distal colon. Br. J. Pharmacol 2010, 159, 222–236. [Google Scholar] [CrossRef]
- Krukoff, T.L. Expression of c-fos in studies of central autonomic and sensory systems. Mol. Neurobiol 1993, 7, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Pongrac, L.; Rylett, R.J. Molecular mechanisms regulating NGF-mediated enhancement of cholinergic neuronal phenotype: C-fos trans-activation of the choline acetyltransferase gene. J. Mol. Neurosci. 1998, 11, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Buéno, L.; Fioramonti, J. Influence of Otilonium Bromide on Spinal c-Fos Expression Induced by Noxious Distension in Rats; Unpublished INRA report 04-005LB on file; Menarini Ricerche Spa: Firenze, Italy, 2005. [Google Scholar]
- Squecco, R.; Garella, R.; Idrizaj, E.; Nistri, S.; Francini, F.; Baccari, M.C. Relaxin Affects Smooth Muscle Biophysical Properties and Mechanical Activity of the Female Mouse Colon. Endocrinology 2015, 156, 4398–4410. [Google Scholar] [CrossRef] [PubMed]
- Garella, R.; Idrizaj, E.; Traini, C.; Squecco, R.; Vannucchi, M.G.; Baccari, M.C. Glucagon-like peptide-2 modulates the nitrergic neurotransmission in strips from the mouse gastric fundus. World J. Gastroenterol. 2017, 23, 7211–7220. [Google Scholar] [CrossRef]
- Traini, C.; Nistri, S.; Calosi, L.; Vannucchi, M.G. Chronic Exposure to Cigarette Smoke Affects the Ileum and Colon of Guinea Pigs Differently. Relaxin (RLX-2, Serelaxin) Prevents Most Local Damage. Front. Pharmacol. 2022, 12, 804623. [Google Scholar] [CrossRef] [PubMed]
- Rychter, J.; Espín, F.; Gallego, D.; Vergara, P.; Jiménez, M.; Clavé, P. Colonic smooth muscle cells and colonic motility patterns as a target for irritable bowel syndrome therapy: Mechanisms of action of otilonium bromide. Ther. Adv. Gastroenterol. 2014, 7, 156–166. [Google Scholar] [CrossRef]
- Xu, C.; Liu, C.; Chen, K.; Zeng, P.; Chan, E.W.C.; Chen, S. Otilonium bromide boosts antimicrobial activities of colistin against Gram-negative pathogens and their persisters. Commun. Biol. 2022, 5, 613. [Google Scholar] [CrossRef]
- Boeckxstaens, G.; Clavé, P.; Corazziari, E.S.; Tack, J. Irritable bowel syndrome: Focus on otilonium bromide. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 131–137. [Google Scholar] [CrossRef]
- Evangelista, S. Benefits from long-term treatment in irritable bowel syndrome. Gastroenterol. Res. Pract. 2012, 2012, 936960. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Traini, C.; Idrizaj, E.; Biagioni, C.; Baccari, M.C.; Vannucchi, M.G. Otilonium Bromide Prevents Cholinergic Changes in the Distal Colon Induced by Chronic Water Avoidance Stress, a Rat Model of Irritable Bowel Syndrome. Int. J. Mol. Sci. 2023, 24, 7440. https://doi.org/10.3390/ijms24087440
Traini C, Idrizaj E, Biagioni C, Baccari MC, Vannucchi MG. Otilonium Bromide Prevents Cholinergic Changes in the Distal Colon Induced by Chronic Water Avoidance Stress, a Rat Model of Irritable Bowel Syndrome. International Journal of Molecular Sciences. 2023; 24(8):7440. https://doi.org/10.3390/ijms24087440
Chicago/Turabian StyleTraini, Chiara, Eglantina Idrizaj, Cristina Biagioni, Maria Caterina Baccari, and Maria Giuliana Vannucchi. 2023. "Otilonium Bromide Prevents Cholinergic Changes in the Distal Colon Induced by Chronic Water Avoidance Stress, a Rat Model of Irritable Bowel Syndrome" International Journal of Molecular Sciences 24, no. 8: 7440. https://doi.org/10.3390/ijms24087440