
Changes in Phytohormones and Transcriptomic Reprogramming in Strawberry Leaves under Different Light Qualities

Abstract
:1. Introduction
2. Results
2.1. Analysis of Physiological Indicators
2.2. Analysis of Phytohormones
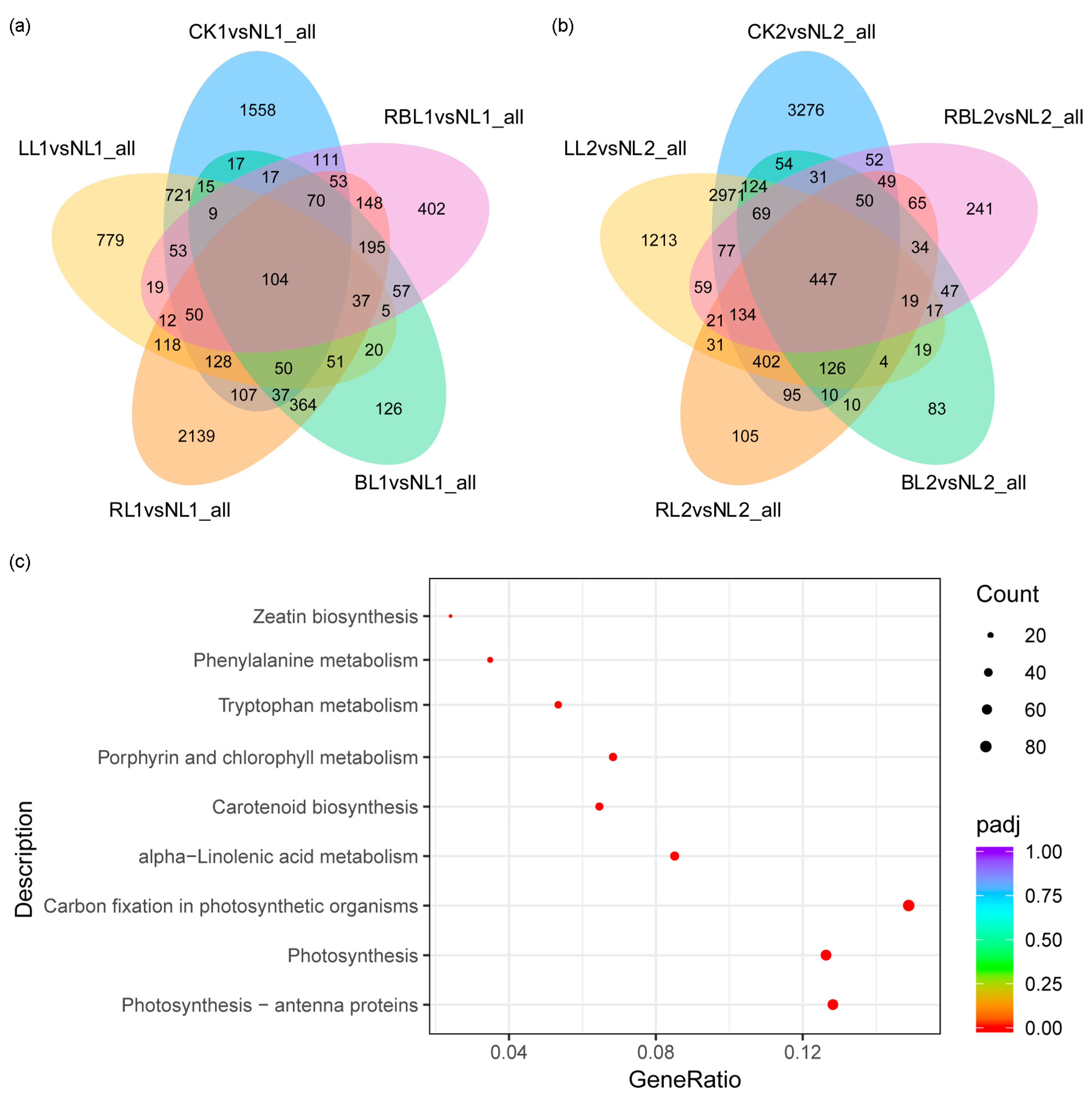
2.3. Analysis of Differential Genes in Transcriptome
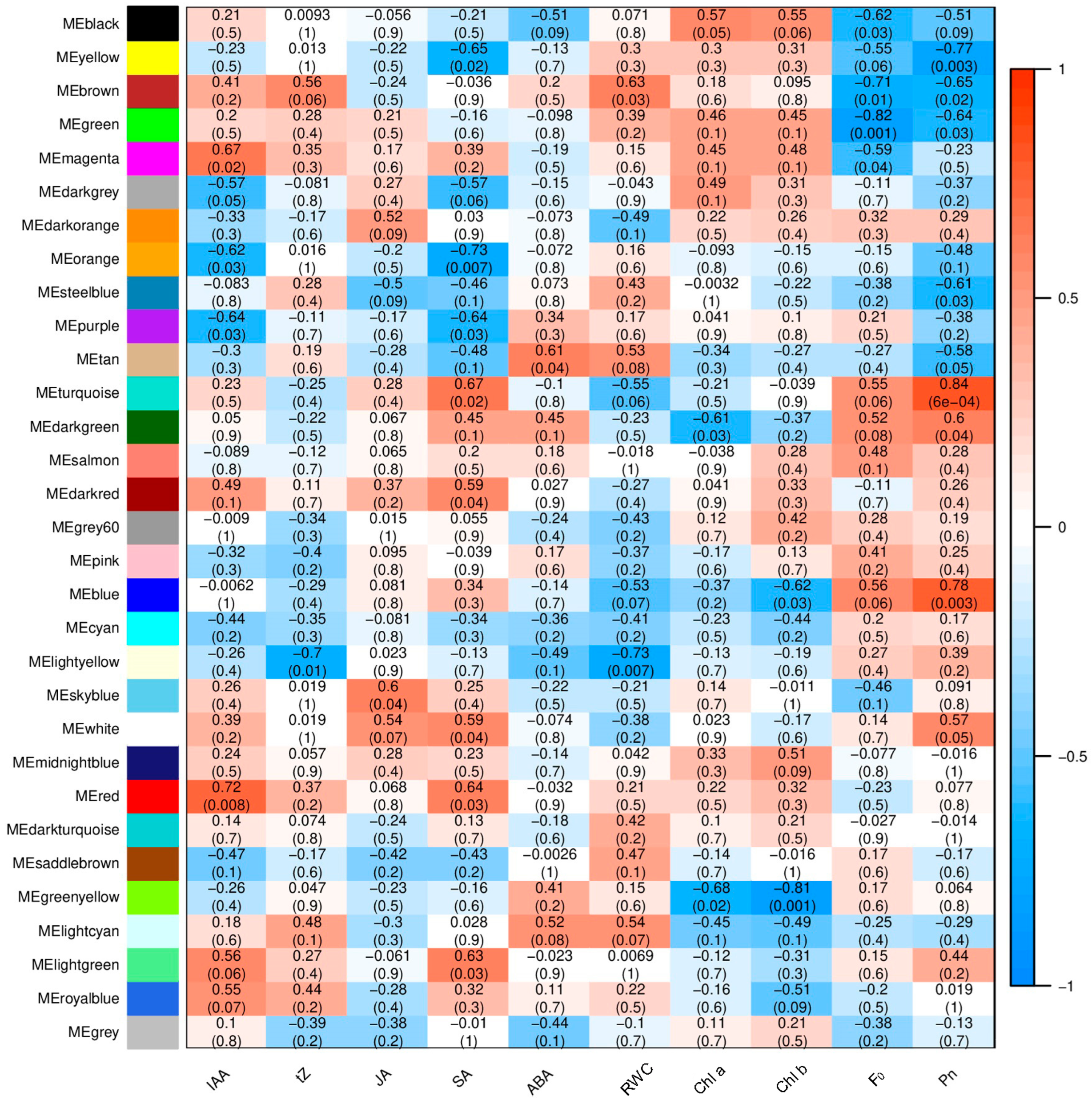
2.4. Analysis of Module–Trait Relationships
2.5. Analysis of 4 Photosynthesis-Related Pathways
2.5.1. Chlorophyll Metabolism
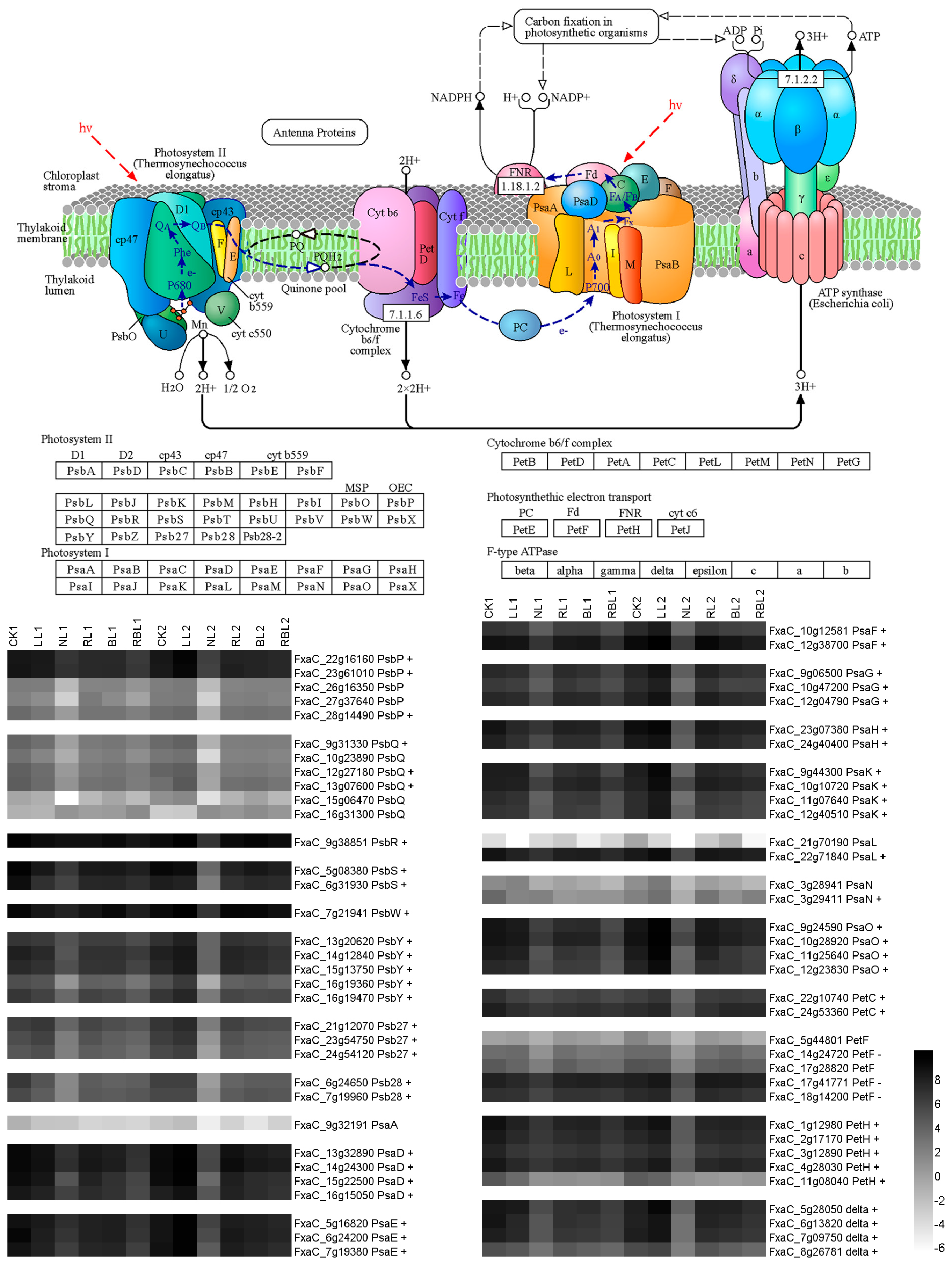
2.5.2. Photosynthesis
2.5.3. Photosynthesis Antenna Proteins
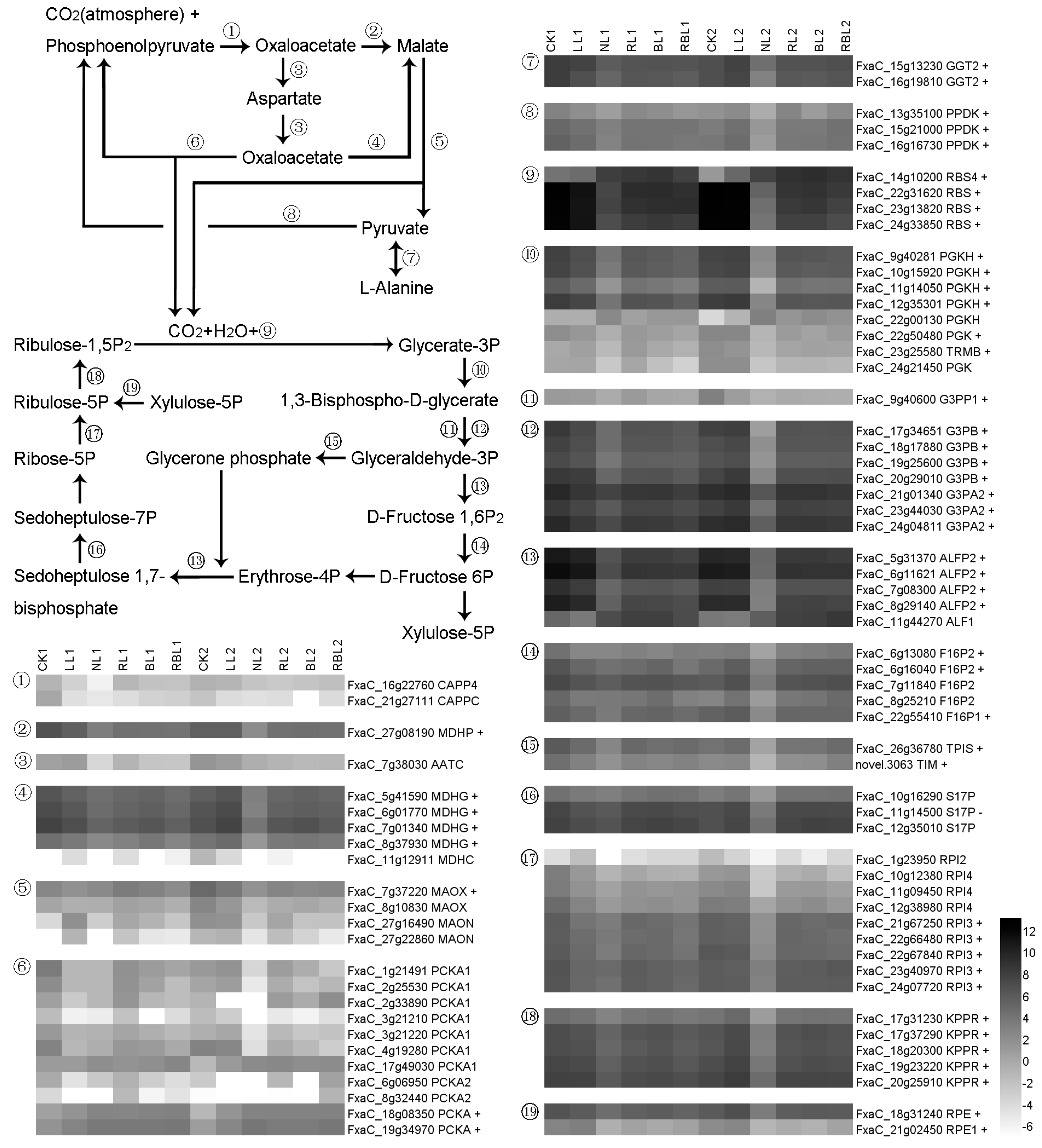
2.5.4. Carbon Fixation in Photosynthetic Organisms
2.6. Analysis of Hormone Synthesis and Signal Transduction
2.6.1. ABA Synthesis and Signal Transduction
2.6.2. IAA Synthesis and Signal Transduction
2.6.3. tZ Synthesis and Signal Transduction
2.6.4. Jasmonic Acid (JA) Synthesis and Signal Transduction
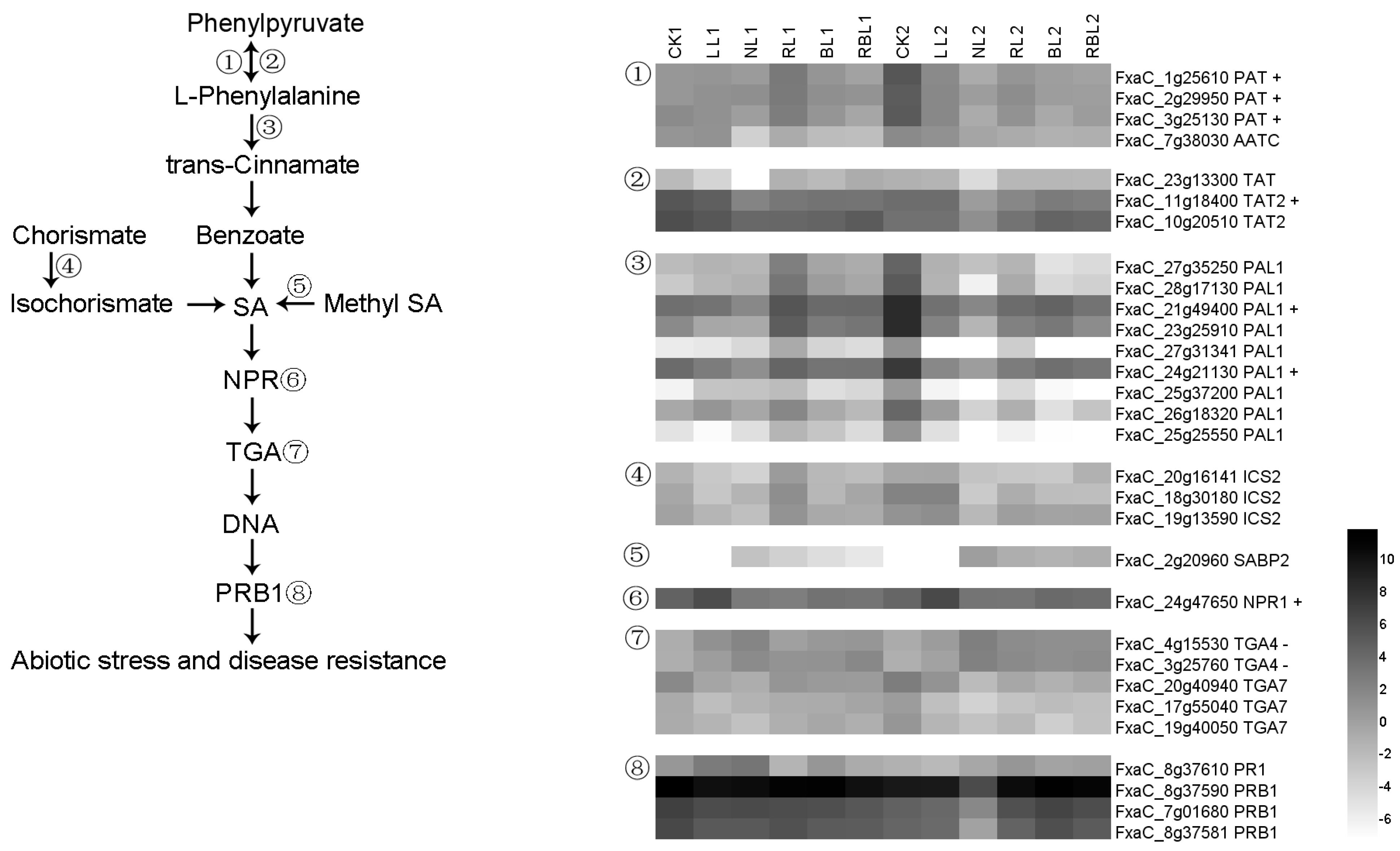
2.6.5. Salicylic Acid (SA) Synthesis and Signal Transduction
2.7. PPI Analysis
3. Discussion
3.1. Effects of Different Light Qualities on Chlorophyll Metabolism
3.2. Effects of Different Light Qualities on Photosynthesis
3.3. Effects of Different Light Qualities on Photosynthesis Antenna Proteins
3.4. Effects of Different Light Qualities on Carbon Fixation
3.5. Effects of Different Light Qualities on Phytohormone Synthesis and Signal Transduction
4. Materials and Methods
4.1. Plant Treatment and Sampling
4.2. Measurement of Relative Water Content
4.3. Measurement of Chl a and Chl b Contents
4.4. Measurement of Pn and F0
4.5. Measurement of Phytohormones
4.6. Measurement of Transcriptomic Reprogramming
4.7. Maps of Metabolic, Module–Trait Relationship, and Protein–Protein Interaction Networks
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Choi, H.G. Correlation among Phenotypic Parameters Related to the Growth and Photosynthesis of Strawberry (Fragaria × ananassa Duch.) Grown Under Various Light Intensity Conditions. Front. Plant. Sci. 2021, 12, 647585. [Google Scholar] [CrossRef]
- Warner, R.; Wu, B.S.; MacPherson, S.; Lefsrud, M. A Review of Strawberry Photobiology and Fruit Flavonoids in Controlled Environments. Front. Plant. Sci. 2021, 12, 611893. [Google Scholar] [CrossRef]
- Malekzadeh Shamsabad, M.R.; Esmaeilizadeh, M.; Roosta, H.R.; Dąbrowski, P.; Telesiński, A.; Kalaji, H.M. Supplemental light application can improve the growth and development of strawberry plants under salinity and alkalinity stress conditions. Sci. Rep. 2022, 12, 9272. [Google Scholar] [CrossRef] [PubMed]
- Guiamba, H.; Zhang, X.; Sierka, E.; Lin, K.; Ali, M.M.; Ali, W.M.; Lamlom, S.F.; Kalaji, H.M.; Telesiński, A.; Yousef, A.F.; et al. Enhancement of photosynthesis efficiency and yield of strawberry (Fragaria ananassa Duch.) plants via LED systems. Front. Plant. Sci. 2022, 13, 918038. [Google Scholar] [CrossRef]
- Yoshida, H.; Mizuta, D.; Fukuda, N.; Hikosaka, S.; Goto, E. Effects of varying light quality from single-peak blue and red light-emitting diodes during nursery period on flowering, photosynthesis, growth, and fruit yield of everbearing strawberry. Plant Biotechnol. 2016, 33, 267–276. [Google Scholar] [CrossRef]
- Ye, Y.; Liu, Y.; Li, X.; Chen, Q.; Zhang, Y.; Luo, Y.; Liu, Z.; Wang, Y.; Lin, Y.; Zhang, Y.; et al. Transcriptome Profile Analysis of Strawberry Leaves Reveals Flowering Regulation under Blue Light Treatment. Int. J. Genom. 2021, 2021, 5572076. [Google Scholar] [CrossRef]
- Nadalini, S.; Zucchi, P.; Andreotti, C. Effects of blue and red LED lights on soilless cultivated strawberry growth performances and fruit quality. Eur. J. Hortic. Sci. 2017, 82, 12–20. [Google Scholar] [CrossRef]
- Rantanen, M.; Kurokura, T.; Mouhu, K.; Pinho, P.; Tetri, E.; Halonen, L.; Palonen, P.; Elomaa, P.; Hytönen, T. Light quality regulates flowering in FvFT1/FvTFL1 dependent manner in the woodland strawberry Fragaria vesca. Front. Plant Sci. 2014, 5, 271. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, L.; Wang, Y.; Zhang, L.; Xu, S.; Wang, X.; He, W.; Zhang, Y.; Lin, Y.; Wang, Y.; et al. The blue light signal transduction module FaCRY1-FaCOP1-FaHY5 regulates anthocyanin accumulation in cultivated strawberry. Front. Plant. Sci. 2023, 14, 1144273. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, W.; Peng, X.; Sun, B.; Wang, X.; Tang, H. Characterization of anthocyanin and proanthocyanidin biosynthesis in two strawberry genotypes during fruit development in response to different light qualities. J. Photochem. Photobiol. B 2018, 186, 225–231. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, L.; Li, Y.; Chen, Q.; Ye, Y.; Zhang, Y.; Luo, Y.; Sun, B.; Wang, X.; Tang, H. Effect of Red and Blue Light on Anthocyanin Accumulation and Differential Gene Expression in Strawberry (Fragaria × ananassa). Molecules 2018, 23, 820. [Google Scholar] [CrossRef]
- Chen, X.; Cai, W.; Xia, J.; Yu, H.; Wang, Q.; Pang, F.; Zhao, M. Metabolomic and Transcriptomic Analyses Reveal that Blue Light Promotes Chlorogenic Acid Synthesis in Strawberry. J. Agric. Food Chem. 2020, 68, 12485–12492. [Google Scholar] [CrossRef]
- Jiang, L.; Chen, X.; Gu, X.; Deng, M.; Li, X.; Zhou, A.; Suo, M.; Gao, W.; Lin, Y.; Wang, Y. Light Quality and Sucrose-Regulated Detached Ripening of Strawberry with Possible Involvement of Abscisic Acid and Auxin Signaling. Int. J. Mol. Sci. 2023, 24, 5681. [Google Scholar] [CrossRef]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Takahashi, A.; Masuda, T.; Noji, S. Light and abscisic acid independently regulated FaMYB10 in Fragaria × ananassa fruit. Planta 2015, 241, 953–965. [Google Scholar] [CrossRef]
- Yue, M.; Jiang, L.; Zhang, N.; Zhang, L.; Liu, Y.; Wang, Y.; Li, M.; Lin, Y.; Zhang, Y.; Zhang, Y. Importance of FaWRKY71 in Strawberry (Fragaria × ananassa) Fruit Ripening. Int. J. Mol. Sci. 2022, 23, 12483. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Deng, M.; Gui, R.; Liu, Y.; Chen, X.; Lin, Y.; Li, M.; Wang, Y.; He, W. Blue light combined with salicylic acid treatment maintained the postharvest quality of strawberry fruit during refrigerated storage. Food Chem. X 2022, 15, 100384. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, M.; Yang, K.N.; Zheng, C.X. Salicylic acid-primed defence response in octoploid strawberry ‘Benihoppe’ leaves induces resistance against Podosphaera aphanis through enhanced accumulation of proanthocyanidins and upregulation of pathogenesis-related genes. BMC Plant Biol. 2020, 20, 149. [Google Scholar] [CrossRef]
- Wang, S.Y. Methyl Jasmonate Reduces Water Stress in Strawberry. J. Plant Growth Regul. 1999, 18, 127–134. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, Z.; Tai, Q.; Shen, C.; Rao, Y.; Zhang, W. Construction of a photosynthetic rate prediction model for greenhouse strawberries with distributed regulation of light environment. Math. Biosci. Eng. 2022, 19, 12774–12791. [Google Scholar] [CrossRef]
- Cao, J.; Govindjee. Chlorophyll a fluorescence transient as an indicator of active and inactive Photosystem II in thylakoid membranes. Biochim. Biophys. Acta 1990, 1015, 180–188. [Google Scholar] [CrossRef]
- Moser, J.; Schubert, W.D.; Beier, V.; Bringemeier, I.; Jahn, D.; Heinz, D.W. V-shaped structure of glutamyl-tRNA reductase, the first enzyme of tRNA-dependent tetrapyrrole biosynthesis. EMBO J. 2001, 20, 6583–6590. [Google Scholar] [CrossRef]
- Sattler, I.; Roessner, C.A.; Stolowich, N.J.; Hardin, S.H.; Harris-Haller, L.W.; Yokubaitis, N.T.; Murooka, Y.; Hashimoto, Y.; Scott, A.I. Cloning, sequencing, and expression of the uroporphyrinogen III methyltransferase cobA gene of Propionibacterium freudenreichii (shermanii). J. Bacteriol. 1995, 177, 1564–1569. [Google Scholar] [CrossRef]
- Frustaci, J.M.; O’Brian, M.R. Characterization of a Bradyrhizobium japonicum ferrochelatase mutant and isolation of the hemH gene. J. Bacteriol. 1992, 174, 4223–4229. [Google Scholar] [CrossRef]
- Bareth, B.; Dennerlein, S.; Mick, D.U.; Nikolov, M.; Urlaub, H.; Rehling, P. The heme a synthase Cox15 associates with cytochrome c oxidase assembly intermediates during Cox1 maturation. Mol. Cell. Biol. 2013, 33, 4128–4137. [Google Scholar] [CrossRef]
- Ibata, H.; Nagatani, A.; Mochizuki, N. CHLH/GUN5 Function in Tetrapyrrole Metabolism Is Correlated with Plastid Signaling but not ABA Responses in Guard Cells. Front. Plant Sci. 2016, 7, 1650. [Google Scholar] [CrossRef]
- Luo, J.; Wang, H.; Chen, S.; Ren, S.; Fu, H.; Li, R.; Wang, C. CmNAC73 Mediates the Formation of Green Color in Chrysanthemum Flowers by Directly Activating the Expression of Chlorophyll Biosynthesis Genes HEMA1 and CRD1. Genes 2021, 12, 704. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, S.; Zhang, M.; Zhang, Y.; Feng, H. Mapping of a Pale Green Mutant Gene and Its Functional Verification by Allelic Mutations in Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Front. Plant Sci. 2021, 12, 699308. [Google Scholar] [CrossRef]
- Zhang, S.; Godwin, A.R.F.; Taylor, A.; Hardman, S.J.O.; Jowitt, T.A.; Johannissen, L.O.; Hay, S.; Baldock, C.; Heyes, D.J. Scrutton NS. Dual role of the active site ‘lid’ regions of protochlorophyllide oxidoreductase in photocatalysis and plant development. FEBS J. 2021, 288, 175–189. [Google Scholar] [CrossRef]
- Lin, Y.P.; Shen, Y.Y.; Shiu, Y.B.; Charng, Y.Y.; Grimm, B. Chlorophyll dephytylase 1 and chlorophyll synthase: A chlorophyll salvage pathway for the turnover of photosystems I and II. Plant J. 2022, 111, 979–994. [Google Scholar] [CrossRef]
- Tian, Y.N.; Tian, Y.N.; Zhong, R.H.; Wei, J.B.; Luo, H.H.; Eyal, Y.; Jin, H.L.; Wu, L.J.; Liang, K.Y.; Li, Y.M. Arabidopsis CHLOROPHYLLASE 1 protects young leaves from long-term photodamage by facilitating FtsH-mediated D1 degradation in photosystem II repair. Mol. Plant 2021, 14, 1149–1167. [Google Scholar] [CrossRef]
- Bell, A.; Moreau, C.; Chinoy, C.; Spanner, R.; Dalmais, M.; Le Signor, C.; Bendahmane, A.; Klenell, M.; Domoney, C.S. SGRL can regulate chlorophyll metabolism and contributes to normal plant growth and development in Pisum sativum L. Plant Mol. Biol. 2015, 89, 539–558. [Google Scholar] [CrossRef]
- He, F.; Shi, Y.J.; Chen, Q.; Li, J.L.; Niu, M.X.; Feng, C.H.; Lu, M.M.; Tian, F.F.; Zhang, F.; Lin, T.T. Genome-Wide Investigation of the PtrCHLP Family Reveals That PtrCHLP3 Actively Mediates Poplar Growth and Development by Regulating Photosynthesis. Front. Plant Sci. 2022, 13, 870970. [Google Scholar] [CrossRef]
- Croce, R.; Morosinotto, T.; Castelletti, S.; Breton, J.; Bassi, R. The Lhca antenna complexes of higher plants photosystem I. Biochim. Biophys. Acta 2002, 1556, 29–40. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Johnson, M.P.; Zia, A.; Ruban, A.V.; Horton, P. The Lhcb protein and xanthophyll composition of the light harvesting antenna controls the DeltapH-dependency of non-photochemical quenching in Arabidopsis thaliana. FEBS Lett. 2008, 582, 1477–1482. [Google Scholar] [CrossRef]
- Shu, J.P.; Yan, Y.H.; Wang, R.J. Convergent molecular evolution of phosphoenolpyruvate carboxylase gene family in C4 and crassulacean acid metabolism plants. PeerJ 2022, 10, e12828. [Google Scholar] [CrossRef]
- Minárik, P.; Tomásková, N.; Kollárová, M.; Antalík, M. Malate dehydrogenases—Structure and function. Gen. Physiol. Biophys. 2002, 21, 257–265. [Google Scholar]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 biochemistry. Cardiovasc. Drugs Ther. 2009, 23, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Kim, C.H.; Choi, Y.; Park, K.M.; Chang, P.S. Catalytic characterization of heterodimeric linoleate 13S-lipoxygenase from black soybean (Glycine max (L.) Merr.). Enzyme Microb. Technol. 2020, 139, 109595. [Google Scholar] [CrossRef]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef]
- Zavaliev, R.; Mohan, R.; Chen, T.; Dong, X. Formation of NPR1 Condensates Promotes Cell Survival during the Plant Immune Response. Cell 2020, 182, 1093–1108.e18. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, M.; Zribi, I.; Missaoui, K.; Drira-Fakhfekh, M.; Azzouzi, B.; Brini, F. Differential regulation of the durum wheat Pathogenesis-related protein (PR1) by Calmodulin TdCaM1.3 protein. Mol. Biol. Rep. 2021, 48, 347–362. [Google Scholar] [CrossRef]
- Minai, L.; Cohen, Y.; Chitnis, P.R.; Nechushtai, R. The precursor of PsaD assembles into the photosystem I complex in two steps. Proc. Natl. Acad. Sci. USA 1996, 93, 6338–6342. [Google Scholar] [CrossRef] [PubMed]
- Krieger-Liszkay, A.; Shimakawa, G.; Sétif, P. Role of the two PsaE isoforms on O2 reduction at photosystem I in Arabidopsis thaliana. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148089. [Google Scholar] [CrossRef]
- Farkas, D.; Franzén, L.G.; Hansson, Ö. Cloning, expression and purification of the luminal domain of spinach photosystem 1 subunit PsaF functional in binding to plastocyanin and with a disulfide bridge required for folding. Protein Expr. Purif. 2011, 78, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Hirano, A.; Hiyama, T.; Inoue, Y. Polypeptide composition of higher plant photosystem I complex. Identification of psaI, psaJ and psaK gene products. FEBS Lett. 1990, 263, 274–278. [Google Scholar] [CrossRef]
- Chitnis, V.P.; Chitnis, V.P.; Xu, Q.; Yu, L.; Golbeck, J.H.; Nakamoto, H.; Xie, D.L.; Chitnis, P.R. Targeted inactivation of the gene psaL encoding a subunit of photosystem I of the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem. 1993, 268, 11678–11684. [Google Scholar] [CrossRef]
- Okada, K. PetH is rate-controlling in the interaction between PetH, a component of the supramolecular complex with photosystem II, and PetF, a light-dependent electron transfer protein. Biochem. Biophys. Res. Commun. 2009, 389, 394–398. [Google Scholar] [CrossRef]
- Johnson, V.M.; Biswas, S.; Roose, J.L.; Pakrasi, H.B.; Liu, H. Psb27, a photosystem II assembly protein, enables quenching of excess light energy during its participation in the PSII lifecycle. Photosynth. Res. 2022, 152, 297–304. [Google Scholar] [CrossRef]
- Suorsa, M.; Sirpiö, S.; Allahverdiyeva, Y.; Paakkarinen, V.; Mamedov, F.; Styring, S.; Aro, E.M. PsbR, a missing link in the assembly of the oxygen-evolving complex of plant photosystem II. J. Biol. Chem. 2006, 281, 145–150. [Google Scholar] [CrossRef]
- García-Cerdán, J.G.; García-Cerdán, J.G.; Kovács, L.; Tóth, T.; Kereïche, S.; Aseeva, E.; Boekema, E.J.; Mamedov, F.; Funk, C.; Schröder, W.P. The PsbW protein stabilizes the supramolecular organization of photosystem II in higher plants. Plant J. 2011, 65, 368–381. [Google Scholar] [CrossRef]
- Alboresi, A.; Ballottari, M.; Hienerwadel, R.; Giacometti, G.M.; Morosinotto, T. Antenna complexes protect Photosystem I from photoinhibition. BMC Plant Biol. 2009, 9, 71. [Google Scholar] [CrossRef]
- Novoderezhkin, V.I.; Croce, R. The location of the low-energy states in Lhca1 favors excitation energy transfer to the core in the plant PSI-LHCI supercomplex. Photosynth. Res. 2023, 156, 59–74. [Google Scholar] [CrossRef]
- Wientjes, E.; Croce, R. The light-harvesting complexes of higher-plant Photosystem I: Lhca1/4 and Lhca2/3 form two red-emitting heterodimers. Biochem. J. 2011, 433, 477–485. [Google Scholar] [CrossRef]
- Ishii, A.; Shan, J.; Sheng, X.; Kim, E.; Watanabe, A.; Yokono, M.; Noda, C.; Song, C.; Murata, K.; Liu, Z.; et al. The photosystem I supercomplex from a primordial green alga Ostreococcus tauri harbors three light-harvesting complex trimers. eLife 2023, 12, e84488. [Google Scholar] [CrossRef] [PubMed]
- Caffarri, S.; Croce, R.; Cattivelli, L.; Bassi, R. A look within LHCII: Differential analysis of the Lhcb1-3 complexes building the major trimeric antenna complex of higher-plant photosynthesis. Biochemistry 2004, 43, 9467–9476. [Google Scholar] [CrossRef]
- Albanese, P.; Manfredi, M.; Marengo, E.; Saracco, G.; Pagliano, C. Structural and functional differentiation of the light-harvesting protein Lhcb4 during land plant diversification. Physiol. Plant 2019, 166, 336–350. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Deng, X.; Tang, X.; Tan, T.; Zhang, D.; Liu, B.; Lin, H. Involvement of Lhcb6 and Lhcb5 in Photosynthesis Regulation in Physcomitrella patens Response to Abiotic Stress. Int. J. Mol. Sci. 2019, 20, 3665. [Google Scholar] [CrossRef]
- Peterson, R.B.; Schultes, N.P. Light-harvesting complex B7 shifts the irradiance response of photosynthetic light-harvesting regulation in leaves of Arabidopsis thaliana. J. Plant Physiol. 2014, 171, 311–318. [Google Scholar] [CrossRef]
- Sun, Y.; Casella, S.; Fang, Y.; Huang, F.; Faulkner, M.; Barrett, S.; Liu, L.N. Light Modulates the Biosynthesis and Organization of Cyanobacterial Carbon Fixation Machinery through Photosynthetic Electron Flow. Plant Physiol. 2016, 171, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.L.; Pavlovic, T.; Torcolese, G.; Badia, M.B.; Gismondi, M.; Maurino, V.G.; Andreo, C.S.; Drincovich, M.F.; Gerrard Wheeler, M.C.; Saigo, M. NADP-Dependent Malic Enzyme 1 Participates in the Abscisic Acid Response in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1637. [Google Scholar] [CrossRef]
- Yadav, S.; Rathore, M.S.; Mishra, A. The Pyruvate-Phosphate Dikinase (C4-SmPPDK) Gene From Suaeda monoica Enhances Photosynthesis, Carbon Assimilation, and Abiotic Stress Tolerance in a C3 Plant Under Elevated CO2 Conditions. Front. Plant Sci. 2020, 11, 345. [Google Scholar] [CrossRef]
- Spreitzer, R.J. Role of the small subunit in ribulose-1,5-bisphosphate carboxylase/oxygenase. Arch. Biochem. Biophys. 2003, 414, 141–149. [Google Scholar] [CrossRef]
- Diop, M.; Gallois, J.L. Exploring New Routes for Genetic Resistances to Potyviruses: The Case of the Arabidopsis thaliana Phosphoglycerates Kinases (PGK) Metabolic Enzymes. Viruses 2022, 14, 1245. [Google Scholar] [CrossRef]
- Simkin, A.J.; Alqurashi, M.; Lopez-Calcagno, P.E.; Headland, L.R.; Raines, C.A. Glyceraldehyde-3-phosphate dehydrogenase subunits A and B are essential to maintain photosynthetic efficiency. Plant Physiol. 2023, 192, 2989–3000. [Google Scholar] [CrossRef]
- Stern, A.L.; Naworyta, A.; Cazzulo, J.J.; Mowbray, S.L. Structures of type B ribose 5-phosphate isomerase from Trypanosoma cruzi shed light on the determinants of sugar specificity in the structural family. FEBS J. 2011, 278, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Xie, Y.; Pan, X.; Zhang, H.; Cao, P.; Su, X.; Chang, W.; Li, M. Photosynthetic Phosphoribulokinase Structures: Enzymatic Mechanisms and the Redox Regulation of the Calvin-Benson-Bassham Cycle. Plant Cell 2020, 32, 1556–1573. [Google Scholar] [CrossRef] [PubMed]
- de Wit, M.; Galvão, V.C.; Fankhauser, C. Light-Mediated Hormonal Regulation of Plant Growth and Development. Ann. Rev. Plant Biol. 2016, 67, 513–537. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, K.; Hwang, H.; Bhatnagar, N.; Kim, D.Y.; Yoon, I.S.; Byun, M.O.; Kim, S.T.; Jung, K.H.; Kim, B.G. Overexpression of PYL5 in rice enhances drought tolerance, inhibits growth, and modulates gene expression. J. Exp. Bot. 2014, 65, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Dong, L.; Wang, E.; Liu, S.; Cheng, Y.; Zhao, J.; Xu, S.; Liang, Z.; Ma, H.; Nie, B. StHAB1, a negative regulatory factor in abscisic acid signaling, plays crucial roles in potato drought tolerance and shoot branching. J. Exp. Bot. 2023, 74, 6708–6721. [Google Scholar] [CrossRef]
- Wong, L.M.; Wong, L.M.; Abel, S.; Shen, N.; de la Foata, M.; Mall, Y.; Theologis, A. Differential activation of the primary auxin response genes, PS-IAA4/5 and PS-IAA6, during early plant development. Plant J. 1996, 9, 587–599. [Google Scholar] [CrossRef]
- Schilmiller, A.L.; Koo, A.J.; Howe, G.A. Functional diversification of acyl-coenzyme A oxidases in jasmonic acid biosynthesis and action. Plant Physiol. 2007, 143, 812–824. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q. Experimental Guidelines in Plant Physiology; China Agricultural Press: Beijing, China, 2018. (In Chinese) [Google Scholar]
- Liu, T.; Li, M.; Liu, Z.; Ai, X.; Li, Y. Reannotation of the cultivated strawberry genome and establishment of a strawberry genome database. Hortic. Res. 2021, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Tian, J.; Guo, C.; Luo, S.; Li, J. Interaction of gibberellin and other hormones in almond anthers: Phenotypic and physiological changes and transcriptomic reprogramming. Hortic. Res. 2021, 8, 94. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CK1 | LL1 | NL1 | RL1 | BL1 | RBL1 | CK2 | LL2 | NL2 | RL2 | BL2 | RBL2 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RWC (%, FW) | 67.51 ± 1.27 F | 71.23 ± 1.12 DE | 71.48 ± 0.39 CDE | 69.85 ± 0.08 E | 70.29 ± 0.37 E | 70.57 ± 0.02 E | 67.1 ± 0.11 F | 73.38 ± 0.06 AB | 74.57 ± 0.66 A | 73.04 ± 1.13 ABC | 72.93 ± 0.65 BC | 72.64 ± 0.01 BCD |
| Chl a (mg/g, FW) | 1.35 ± 0.2 AB | 1.60 ± 0.06 AB | 1.70 ± 0.06 A | 1.37 ± 0.21 AB | 1.52 ± 0.11 AB | 1.61 ± 0.04 AB | 1.34 ± 0.13 AB | 1.45 ± 0.23 AB | 1.43 ± 0.31 AB | 1.47 ± 0.38 AB | 1.11 ± 0.24 BC | 1.82 ± 0.08 A |
| Chl b (mg/g, FW) | 0.44 ± 0.05 AB | 0.53 ± 0.02 A | 0.54 ± 0.02 A | 0.42 ± 0.07 ABC | 0.44 ± 0.03 AB | 0.50 ± 0.01 AB | 0.15 ± 0.04 D | 0.42 ± 0.07 ABC | 0.35 ± 0.09 BC | 0.35 ± 0.11 BC | 0.28 ± 0.08 CD | 0.47 ± 0.02 AB |
| Pn (μmol/(m2·s)) | 27.73 ± 0.4 A | 8.30 ± 0.2 D | −3.60 ± 0.1 I | 1.90 ± 0.1 E | 0.63 ± 0.06 H | 0.87 ± 0.06 G | 32.47 ± 1.16 B | 10.00 ± 0.1 C | −3.10 ± 0.1 I | 2.43 ± 0.06 E | 1.00 ± 0.1 FG | 1.77 ± 0.06 EF |
| F0 | 3790 ± 385 AB | 3536 ± 150 AB | 2328 ± 257 C | 3305 ± 349 B | 2629 ± 362 C | 3533 ± 181 AB | 4165 ± 127 A | 3673 ± 316 AB | 2546 ± 222 C | 3637 ± 484 AB | 3423 ± 176 AB | 3988 ± 253 AB |
| CK1 | LL1 | NL1 | RL1 | BL1 | RBL1 | CK2 | LL2 | NL2 | RL2 | BL2 | RBL2 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Indole-3-acetic acid | 6.72 ± 1.21 AB | 6.43 ± 0.68 ABC | 5.07 ± 2.65 BCD | 2.02 ± 0.56 D | 5.50 ± 1.33 ABCD | 5.06 ± 1 BCD | 6.23 ± 1.76 ABC | 3.73 ± 0.83 BCD | 8.73 ± 2.32 A | 3.80 ± 0.6 BCD | 3.33 ± 0.73 BCD | 3.14 ± 0.36 CD |
| 3-Indolebutyric acid | 67.57 ± 1.48 A | 34.00 ± 2.27 C | 40.92 ± 13.85 BC | 43.87 ± 4.88 BC | 40.21 ± 5.49 BC | 56.76 ± 12.26 AB | 53.42 ± 14.2 ABC | 48.01 ± 3.23 ABC | 43.46 ± 7.07 BC | 46.40 ± 8.94 BC | 39.33 ± 8.08 BC | 37.89 ± 3.24 BC |
| Indole-3-carboxylic acid | 2.89 ± 0.93 BCDEF | 4.57 ± 0.66 AB | 5.61 ± 1.54 A | 3.99 ± 0.57 ABCD | 1.06 ± 0.46 EF | 3.76 ± 0.48 ABCD | 0.77 ± 0.67 F | 3.30 ± 0.81 ABCDE | 4.19 ± 0.89 ABC | 3.19 ± 1.85 ABCDE | 1.79 ± 0.34 CDEF | 1.71 ± 0.98 DEF |
| Indole-3-carboxaldehyde | 5.18 ± 1.86 CD | 14.53 ± 1.03 A | 5.17 ± 2.67 CD | 5.61 ± 0.9 C | 1.29 ± 0.5 E | 10.95 ± 1.33 B | 4.03 ± 2.06 CDE | 7.32 ± 0.77 C | 6.73 ± 1.12 C | 6.17 ± 1.44 C | 5.03 ± 1.6 CD | 1.69 ± 1.14 DE |
| N6-Isopentenyladenine | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.48 ± 0.03 A | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.31 ± 0.06 B | 0 ± 0 |
| Isopentenyl adenosine | 1.07 ± 0.01 DE | 1.32 ± 0.03 CD | 1.02 ± 0.19 E | 0.64 ± 0.02 F | 0.75 ± 0.07 F | 1.57 ± 0.08 C | 1.01 ± 0.05 E | 0.51 ± 0.04 F | 2.66 ± 0.26 A | 2.52 ± 0.14 A | 0.7 ± 0.04 F | 2.26 ± 0.1 B |
| Trans-zeatin-riboside | 0.75 ± 0.03 BC | 0.89 ± 0.07 B | 0.7 ± 0.11 BCD | 0.51 ± 0.03 DE | 0.39 ± 0.05 E | 0.45 ± 0.18 E | 0.59 ± 0.09 CDE | 0.54 ± 0.1 CDE | 1.75 ± 0.11 A | 1.92 ± 0.11 A | 0.47 ± 0.05 E | 0.90 ± 0.03 B |
| Kinetin | 0.8 ± 0.22 A | 0 ± 0 A | 0.15 ± 0.13 A | 0.11 ± 0.19 A | 0 ± 0 A | 0.14 ± 0.25 A | 0.7 ± 0.24 A | 0.09 ± 0.16 A | 0.42 ± 0.73 A | 0.26 ± 0.45 A | 0.21 ± 0.36 A | 0 ± 0 A |
| Methyljasmonate | 25.46 ± 9.2 B | 41.85 ± 15.91 A | 7.18 ± 1.84 C | 0 ± 0 | 0 ± 0 | 0 ± 0 | 30.96 ± 9.29 AB | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| N-Jasmonic acid isoleucine | 704.08 ± 218.74 A | 422.87 ± 67.59 B | 225.95 ± 37.27 C | 33.18 ± 12.74 D | 9.79 ± 7.01 D | 0 ± 0 D | 239.24 ± 43.57 C | 36.9 ± 2.17 D | 5.52 ± 1.06 D | 16.28 ± 9.51 D | 0 ± 0 D | 6.8 ± 0.93 D |
| (±)-Jasmonic acid | 365.15 ± 99.11 AB | 213.45 ± 36.31 BCD | 403.06 ± 96.53 A | 163.91 ± 36.25 CDE | 89.9 ± 43.19 DE | 0 ± 0 E | 269.16 ± 26.11 ABC | 25.25 ± 4.49 E | 0 ± 0 E | 378.77 ± 185.82 AB | 47.62 ± 11.82 DE | 131.88 ± 27.96 CDE |
| Salicylic acid | 5936.83 ± 1287.12 A | 4600.56 ± 621.4 BC | 3483.4 ± 382.85 CD | 2661.81 ± 429.51 D | 3373.65 ± 175.1 CD | 2946.54 ± 554.53 D | 5209.66 ± 456.41 AB | 3505.46 ± 197.15 CD | 4962.73 ± 319.12 AB | 3085.64 ± 74.07 D | 3140.04 ± 78.2 D | 2969.25 ± 245.35 D |
| Abscisic acid | 299.18 ± 39.15 ABCD | 255.36 ± 92.27 BCD | 204.43 ± 38.08 CD | 156.61 ± 30.43 D | 222.18 ± 15.68 CD | 170.41 ± 18.89 D | 303.49 ± 29.26 ABCD | 213.36 ± 37.2 CD | 346.35 ± 45.81 ABC | 406.39 ± 167.28 AB | 446.36 ± 61.02 A | 355.33 ± 15.02 ABC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Wang, Z.; Wang, X.; Liu, F.; Wang, H. Changes in Phytohormones and Transcriptomic Reprogramming in Strawberry Leaves under Different Light Qualities. Int. J. Mol. Sci. 2024, 25, 2765. https://doi.org/10.3390/ijms25052765
Li P, Wang Z, Wang X, Liu F, Wang H. Changes in Phytohormones and Transcriptomic Reprogramming in Strawberry Leaves under Different Light Qualities. International Journal of Molecular Sciences. 2024; 25(5):2765. https://doi.org/10.3390/ijms25052765
Chicago/Turabian StyleLi, Peng, Zhiqiang Wang, Xiaodi Wang, Fengzhi Liu, and Haibo Wang. 2024. "Changes in Phytohormones and Transcriptomic Reprogramming in Strawberry Leaves under Different Light Qualities" International Journal of Molecular Sciences 25, no. 5: 2765. https://doi.org/10.3390/ijms25052765