
Membrane Interaction Characteristics of the RTX Toxins and the Cholesterol-Dependence of Their Cytolytic/Cytotoxic Activity

1
Department of Biochemistry and Molecular Biology, Faculty of Science and Technology, University of the Basque Country UPV/EHU, 48080 Bilbao, Spain
2
Biofisika Institute (UPV/EHU, CSIC), 48940 Leioa, Spain
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(6), 3131; https://doi.org/10.3390/ijms25063131
Submission received: 20 February 2024
/
Revised: 4 March 2024
/
Accepted: 6 March 2024
/
Published: 8 March 2024
(This article belongs to the Collection Feature Paper Collection in Biochemistry)
Abstract
:RTX toxins are important virulence factors produced by a wide range of Gram-negative bacteria. They are secreted as water-soluble proteins that are able to bind to the host cell membrane and insert hydrophobic segments into the lipid bilayer that ultimately contribute to the formation of transmembrane pores. Ion diffusion through these pores leads then to cytotoxic and cytolytic effects on the hosts. Several reports have evidenced that the binding of several RTX toxins to the target cell membrane may take place through a high-affinity interaction with integrins of the β2 family that is highly expressed in immune cells of the myeloid lineage. However, at higher toxin doses, cytotoxicity by most RTX toxins has been observed also on β2-deficient cells in which toxin binding to the cell membrane has been proposed to occur through interaction with glycans of glycosylated lipids or proteins present in the membrane. More recently, cumulative pieces of evidence show that membrane cholesterol is essential for the mechanism of action of several RTX toxins. Here, we summarize the most important aspects of the RTX toxin interaction with the target cell membrane, including the cholesterol dependence, the recent identification in the sequences of several RTX toxins of linear motifs coined as the Cholesterol Recognition/interaction Amino acid Consensus (CRAC), and the reverse or mirror CARC motif, which is involved in the toxin–cholesterol interaction.
1. RTX Toxins
Pore-forming RTX toxins constitute a family of pore-forming cytotoxins/cytolysins secreted by many Gram-negative pathogens, including the bacteria of the genera Actinobacillus, Aggregatibacter, Bordetella, Escherichia, Kingella, Mannheimia, Moraxella, Morganella, Pasteurella, Proteus, and Vibrio [1,2] and are implicated in the infectious diseases caused by said pathogens.
RTX toxins are large protein toxins with molecular masses between ~100 and 200 kDa (Kingella kingae RtxA cytotoxin, 105 kDa; uropathogenic Escherichia coli (UPEC) α hemolysin (HlyA), 110 kDa; Bordetella pertussis adenylate cyclase toxin (CyaA), 177 kDa) and consist of single polypeptide chains lacking cysteine residues. Synthesis, maturation, and secretion of RTX toxins are determined by the rtxCABD operon [2,3,4]. Gene product A is the polypeptide corresponding to a protoxin (pro-RTX) that matures in the bacterial cytosol to the active form by post-translational acylation at two conserved internal lysine residues [5]. Fatty acylation is mediated by a specific acyltransferase encoded by the product of the gene C and an acyl carrier protein (Acyl-ACP) [5,6,7,8,9,10]. The mature, acylated RTX toxin is then directly secreted across both membranes by the type I secretion system (T1SS) constituted by the genes products B and D and the bacterial outer membrane TolC protein [11,12,13,14,15] (see Figure 1). There may be some minor exceptions to this general mechanism of secretion for some RTX toxins.
The mature polypeptide chain of each RTX toxin consists of four conserved regions [1,2,3,4] (Figure 2). An N-terminal hydrophobic domain harbors several putative transmembrane α-helices, which is important for insertion of the RTX toxin into the host cell membrane and pore formation [1,16,17,18,19,20,21,22,23]. This hydrophobic domain is followed by an acylation domain, a segment of variable length that contains the conserved acylated lysine residues [5,7]. The C-terminal portion of each RTX toxin contains a calcium-binding domain comprising tandem repeats of a nine-residue calcium-binding motif that upon Ca2+ binding adopts a “β-roll” fold [1,2,3,4,24,25,26]. The common presence of these repetitions is precisely at the origin of the historical name of the RTX protein family, where RTX stands for repeat-in-toxin [1]. The C-terminus of the toxin contains a secretion signal that is recognized by the T1SS [27,28,29]. The only exception to the arrangement of typical RTX toxins is the B. pertussis CyaA toxin in which an enzymatic adenylate cyclase (AC) domain is fused to the N-terminus of the RTX hemolysin via a specific linker also named translocation region (Figure 2) [2].
RTX toxins are initially expressed as inactive protoxins that require post-translational acylation to become mature toxins [2,5]. This post-translational acylation results in the addition of fatty acids with fourteen to eighteen carbons to two conserved internal lysine residues located downstream of the hydrophobic domain [5,6,7,8,9]. The fatty acids, which can be saturated, unsaturated, and hydroxylated, are attached via amide linkages to the ε-amino groups of the conserved lysines [5,9,30,31]. Fatty acylation of the RTX toxins is required for host cell cytotoxicity [7,32], but the exact role of the post-translational modification in the mechanism of action of these toxins is not fully understood. Interestingly, this activation is not necessary for the secretion of the RTX protein, since nonacylated proHlyA is secreted as efficiently as acylated HlyA [33]. The nonacylated proHlyA and proCyaA form pores in planar lipid bilayers with a much-reduced propensity, but the formed pores have apparently quite similar properties to the pores generated by the mature toxin form [34,35]. Both nonacylated proHlyA and proCyaA are also quite active in permeabilizing a naked liposome membrane [36,37], suggesting that the fatty acids are not essential for toxin penetration into the lipid bilayer. However, the post-translational modification of proRTXA is critical for the folding of the RTX toxin outside of the bacterial cell [38]. It was shown that the acyl chains covalently bound to B. pertussis CyaA affect toxin folding and stability [38]. Future work may demonstrate a similar role in other RTX toxins.
In addition to the post-translational activation for exerting biological activity, all RTX toxins need to be activated by binding Ca2+ within the acidic glycine- and aspartate-rich nonapeptide repeats. The binding of calcium to the repeats occurs at the extracellular medium, upon toxin secretion, since toxin affinity for this cation is in the mM range, while the intracellular Ca2+ is about 0.1 µM [39,40,41]. It is believed that in the bacterial cytosol, the repeats domain exists in a disordered state [42]. This may allow the RTX toxin to readily pass through the T1SS machinery. Once in the extracellular medium, calcium binding to the nonapeptide repeats then appears to promote folding and adoption of a functional conformation on the secreted RTX proteins [26,43,44].
Upon release from the bacterial interior, the RTX toxins presumably interact with the target cell membrane, after which helices of the pore-forming domain insert into the lipid bilayer forming hydrophilic pores [17,19,21,45,46,47]. The uncontrolled flow of ions through these pores then leads to alterations in the target cell functions and eventually to cell lysis [2].
2. Interaction of the RTX Toxins with the Target Cell Membrane
Based on species and cellular specificity, RTX toxins have been historically divided into two groups, namely, the RTX hemolysins, which are capable of lysing erythrocytes and exhibit toxicity to various cell types isolated from different species, and the RTX leukotoxins, which exhibit narrow species and cell specificity because they bind via a proteinaceous receptor expressed on leukocytes [2]. Numerous reports have recently evidenced, however, that binding of almost any RTX toxin to the target cell membrane may take place through a high-affinity interaction to a proteinaceous receptor, but also in the absence of a specific protein receptor.
2.1. Binding through Cell-Specific Receptors
Various RTX toxins have been shown to specifically recognize several members of the β2 integrin family expressed on leukocytes [2,48,49,50,51], making the white cells highly sensitive to the cytotoxic action of those RTX toxins. This is the case of the leukotoxins LtxA and LktA secreted by Aggregatibacter actinomycetemcomitans and Mannheimia haemolytica, respectively, that use the αLβ2 (LFA-1 or CD11a/CD18) integrin as a receptor for binding to leukocytes [50]. More recently, HlyA of UPEC E. coli has also been shown to bind to αLβ2 in leukocytes [49]. The CyaA of B. pertussis, in turn, binds to myeloid cells through the αMβ2 (Mac-1 or CD11b/CD18) integrin [48]. One of the RTX toxins of Actinobacillus pleuropneumoniae, ApxIIIA, also interacts with the CD18 subunit of the β2 integrins [51]. High-affinity binding to a proteinaceous receptor allows these toxins to be effective at very low concentrations, and it induces a variety of non-lytic effects on the target cells: disruption of bactericidal functions, stimulation or suppression of the release of pro-inflammatory cytokines, modulation of various signaling and proteolytic cascades, induction of cell cycle arrest, or activation of caspases, among others [52,53,54,55,56,57,58,59].
The interaction of B. pertussis CyaA with αMβ2 has been the most explored [60,61]. In 2001, Guermonprez and colleagues reported that CyaA uses the αMβ2 integrin (CD11b/CD18) as a cell receptor in macrophages, neutrophils, and dendritic and natural killer cells [48] (Figure 3). Two years later, in 2003, the same group reported that the αMβ2-binding site was localized to within residues 1166–1281 of the CyaA RTX domain [60]. More recently, and using cryoelectron microscopy, the structure at 2.7 Å resolution of a CyaA fragment (RTX751, residues 751–1706) bound to αMβ2 ectodomain has been determined [62]. This structure has revealed that CyaA interacts with the headpiece (β-propeller and thigh domains) and calf-2 domain of the αM subunit of the integrin in a non-canonical manner specific to bent, inactive αMβ2. CyaA, in turn, engages αM using the inter-blocks linkers L1 and L2 that connect, respectively, the blocks I and II, and II and III, of the calcium-binding domain. Interestingly, the binding of the RTX domain to the αMβ2 ectodomain positions the two acylation sites, Lys860 and Lys983, at the plane of the host-cell membrane, with both Lys side chains pointing toward the membrane, suggesting that the essential acylations in pore-forming RTX toxins are involved in direct insertion into the target membrane. Strikingly, in this study, a non-acylated CyaA RTX fragment was used as a binding partner for the integrin. In contrast, acylation was shown to be required for a productive and tight interaction of the toxin with cells expressing CD11b [60]. Moreover, it has been reported that the presence of acyl chains in CyaA induces a significant stabilization of the apolar segments of the hydrophobic domain and of most of the acylation region so that CyaA acylation is essential for the protein refolding into its active conformation [38].
Regarding A. actinomycetemcomitans LtxA, surface plasmon resonance (SPR) experiments showed a strong affinity of LtxA for the cytosolic domains of both the CD11a and CD18 subunits, whereas the affinity of the toxin for the cytoplasmic domains of the CD11b and CD11c subunits was significantly lower [63]. More recently, the group of Dr. Welch has performed an unbiased genome-wide positive selection in a mutant library of U-937 cells [64]. The selection results have shown that the CD18 subunit is necessary and sufficient for the cytotoxic activity of HlyA, whereas all four alpha subunits are not required at all for the cytotoxic activity of the toxin. In the same study, it was shown that LtxA toxin also binds to the β2 integrin β subunit [64]. One interesting point is that, different to the recently discovered binding site in B. pertussis CyaA (linkers L1 and L2) at binding to αMβ2 [62], the calcium-binding domain of the other RTX toxins do not possess inter-block linkers, raising the question as to how they bind their integrin receptors. In this regard, it was recently noted by Masin and colleagues that swapping the CyaA RTX domain for the RTX domain of E. coli HlyA altered the specificity of CyaA such that it required αLβ2 instead of αMβ2 on the target cell [65]. Thus, at least some pore-forming RTX toxins may bind to integrin receptors via RTX domains that lack linker modules.
2.2. Binding in the Absence of a Specific Proteinaceous Receptor
Although the high-affinity, specific interaction to β2 integrins converts the leukocytes as the most likely physiological targets for the RTX toxins, cytotoxicity by most RTX is also observed on β2-deficient cells at higher toxin doses. Moreover, for some RTX toxins such as HlyA, CyaA, and LtxA, interaction with protein-free liposomes and planar lipid bilayers has been documented [66,67,68,69].
Human leukocytes are 100-fold more sensitive to E. coli HlyA than either bladder or kidney epithelial cells [64]. However, at higher concentrations, HlyA is also cytotoxic to those renal cells, and to a wide range of hosts and cell types including erythrocytes, granulocytes, monocytes, endothelial cells, or renal epithelial cells from mice, ruminants, and primates [70,71,72,73,74,75,76]. Similarly, A. actinomycetemcomitans LtxA and M. haemolytica LktA exhibit detectable hemolytic activity on erythrocytes at high toxin concentrations [77,78], and B. pertussis CyaA at high concentrations can intoxicate epithelial cells, fibroblasts, and erythrocytes [2]. Another RTX toxin, the K. kingae RtxA toxin, is cytotoxic to synovial cells, bone osteosarcoma cells, respiratory epithelial cells, and sheep erythrocytes [79,80]. Moreover, several RTX toxins have been shown to bind and permeabilize even artificial lipid vesicles (liposomes) composed only of phospholipids [81,82,83]. All this suggests, thus, that RTX toxins may productively interact with the cell membrane through a “protein-receptor”-independent way.
Using model lipid membranes devoid of specific proteinaceous receptors, our laboratory documented that the interaction of UPEC HlyA with the lipid bilayer appears to occur in two steps, beginning with a reversible adsorption step that is sensitive to electrostatic forces, which is followed by an irreversible membrane insertion step [84,85]. Adsorption of RTX toxins is detectable in both toxin-sensitive cells and certain toxin-resistant cells [86]. Studies with the isolated calcium-binding domain of HlyA revealed that this part of the protein adsorbs on the membrane in the early stages of HlyA–membrane interaction [87].
For several RTX toxins, it has been reported that toxin binding to the cell membrane occurs through interaction with glycans present in glycosylated lipids or proteins. In 2006, Balashova and colleagues documented that purified A. actinomycetemcomitans LtxA was able to lyse human and sheep erythrocytes, though the toxin concentration required to lyse erythrocytes was higher than that required to kill leukocytes, and concluded that binding to erythrocytes might be mediated by “low-affinity interaction” to some component present in the erythrocyte membrane [77]. A few years later, the same group showed that each of the five different gangliosides (GM1, GM3, GD1a, GD1b, and GT1b), containing at least one sialic acid residue, could completely block LtxA-mediated hemolysis in a dose-dependent manner [88] (Structure of GM1 and GM3 shown in Figure 4). In contrast, asialo-GM1 and free sialic acid were unable to completely block hemolysis. This suggested that the sialic acid residue is a necessary component of gangliosides required for the interaction of LtxA with erythrocytes but is not sufficient on its own to inhibit hemolysis. The results were confirmed in ganglioside-rich C6 rat glioma cells, which are recognized but not killed by LtxA and to which binding of the toxin was successfully blocked by several different gangliosides (GM1, GM3, GD1a, and GD3). In contrast, gangliosides could only partially block the LtxA-mediated killing of β2 integrin-expressing THP-1 cells when the ratio of gangliosides to LtxA was high and the toxin was incubated with THP-1 cells for a short incubation period [88]. The authors concluded that gangliosides act as functional receptors on erythrocytes but not on leukocytes or other cells (e.g., C6 glioma cells) [88]. By contrast, other groups reported that sialic acid residues are important for LtxA-induced cell lysis, regardless of whether the sialic acid residues are linked to the glycosylated β2 integrins or other glycosylated cell surface structures [89]. The authors found that preincubation of human or mouse erythrocytes with neuraminidase, an enzyme that catalyzes the hydrolysis of sialic acid residues from various substrates (glycoproteins, glycolipids, and oligosaccharides), significantly decreased LtxA-mediated hemolysis in a concentration-dependent manner. Similarly, removal of sialic acid residues significantly decreased LtxA-induced lysis of β2 integrin-expressing K562 cells [89]. Another group had similarly reported that β2 integrin-expressing Jurkat T cells pretreated with a mixture of neuraminidase and two other glycosidases, PNGase F and Endo H, were less sensitive to LtxA than untreated Jurkat T cells [90]. This suggests, hence, that in β2 integrin-negative cells, negatively charged sialic acid residues that are part of numerous glycosylated cell surface structures, such as glycoproteins, glycolipids, and even gangliosides may act as low-specific binding sites for LtxA. In the case of the β2 integrin-expressing white blood cells, it can be hypothesized that some saccharide(s) attached to the integrin molecules might be relevant in the interaction with LtxA.
Various early reports had pointed as well to the involvement of gangliosides in the B. pertussis CyaA interaction with the cell membrane, even in integrin-expressing cells. Gable and colleagues (1985) showed that cytotoxicity of CyaA to polymorphonuclear leukocytes could be inhibited by pretreating the cells with neuraminidase or by preincubation of the toxin with bovine brain gangliosides [91]. Later, another laboratory showed that preincubation of CyaA with different types of gangliosides (GM1, GM3, and GT1b) inhibits the CyaA-catalyzed cAMP intoxication of CHO cells lacking CD11b/CD18 [92]. Further studies showed that the pretreatment of GM1-positive human erythrocytes and the CD11b/CD18-negative K562 cells with GM1-binding cholera toxin subunit B (CTB) decreased CyaA binding by ~30%, indicating that CTB competes with CyaA for a binding site on GM1 [93]. Another group reported that CyaA binding to several types of CD11b/CD18-expressing cells (CHO-CD11b/CD18, J774A.1, and human neutrophils) was decreased by ~80% when terminal sialic acid residues of CD11b/CD18 and other cell surface glycoproteins were removed by neuraminidase [90]. Further, they observed an almost complete loss of CyaA binding to those cells that were removed by the glycosidase PNGase F when N-linked oligosaccharides of surface glycoproteins or when N-glycosylation of newly synthesized proteins was blocked with the nucleoside antibiotic tunicamycin [90,91,92,93,94]. The authors concluded that the N-glycosylation of CR3 is crucial for the initial recognition of the integrin receptor by CyaA and subsequent cytotoxic activities of the toxin and suggested that CyaA selectively recognized sugar residues of N-linked oligosaccharides of integrins [90]. Interestingly, the ternary complex (Fab M1F5-RTX751-αMβ2 ectodomain) map recently solved by cryomicroscopy also contains a well-resolved N-linked glycan at Asn1059 in αM calf-2 domain, with the core fucose residue of the glycan packing onto RTX751, Leu1124, and Phe1125 residues [62]. Earlier, a study by Hasan and colleagues (2015) showed that the glutamine substitutions of asparagine residues at positions 469, 692, 801, 900, 978, 1021, and 1075 of the CD11b subunit each individually reduced binding of CyaA by about 8–44%. Further, the intracellular cAMP levels in cells expressing mutant integrins with substitutions at positions 469, 801, 978, and 1075 of CD11b were reproducibly reduced by about 30–38% after treatment with CyaA. The solved structure does not reveal, however, interactions involving any of those mutated asparagine residues, suggesting that those glycans would not be strictly required for CyaA binding to the CD11b/CD18 integrin.
For K. kingae RtxA toxin, Rahman and colleagues have recently reported that the toxin depends on oligosaccharides present on the host cell surface for cell binding [95]. The authors noted that pre-incubation of different cell types with neuraminidase significantly reduces the binding and cytotoxicity of RtxA. Moreover, free sialic acid partially blocked the binding of RtxA to the cells. In addition, the same authors showed that both enzymatic (PNGase F, O-glycosidase) and inhibitor-mediated (tunicamycin, benzyl-2-acetamido-2-deoxy-α-D-galactopyranoside) removal of N- or O-linked oligosaccharide chains from cell surface glycosylated structures resulted in a significant loss of RtxA binding, and deglycosylated cells were more resistant to the cytotoxic effect of RtxA than untreated cells [95]. This suggested that RtxA not only recognizes sialic acid residues but also other saccharide units of the cell surface glycoproteins on the cell surface [95]. All these results have led some authors to raise the possibility that the initial unsaturated binding of RTX cytotoxins to various cells might occur through the recognition of glycosylated membrane components, such as glycoproteins and gangliosides.
3. Cholesterol Dependence of the Cytolytic/Cytotoxic Activity of RTX Toxins
Cumulative pieces of evidence show that membrane cholesterol is essential for the mechanism of action of several RTX toxins. Even in the target cells that express the β2 integrins that act as specific toxin receptors, RTX toxins show cholesterol-dependence.
For A. actinomycetemcomitans LtxA, removal of cholesterol from the host cell membrane with methyl-β-cyclodextrin (MβC) was shown to significantly inhibit the toxin’s ability to kill Jurkat (Jn.9) and THP-1 cells [96,97]. After replenishment of the plasma membrane cholesterol using MβC, followed by MβC-cholesterol incubation, the immune cells became again susceptible to LtxA [97]. Similarly, it was reported that a decrease in the cholesterol content of the plasma membrane of J774A.1 macrophages by MβC yielded a significant decrease in the capacity of B. pertussis CyaA to translocate the AC domain across the cell membrane [98]. Much earlier, in 2004, Martin and colleagues had reported that cholesterol substantially increases the rate of CyaA-induced membrane lysis, measured as the efflux of fluorescent liposomal content, in a dose-dependent manner [66]. In 2009, Herlax and colleagues showed that cholesterol-depleted erythrocytes are less sensitive to the hemolytic activity of UPEC HlyA than control erythrocytes [99]. Later, they noted that the incorporation of cholesterol into phospholipid bilayers promoted the irreversible insertion of the toxin into the membrane, which increased the toxin’s lytic activity [100]. More recently, our laboratory observed that treatment of erythrocytes with cholesterol oxidase notably reduces the CyaA-induced hemolysis [101] and that incorporation of cholesterol in pure Dipalmitoylphosphatidylcholine (DOPC) liposomes enhances the lytic capacity of CyaA on the vesicles [101]. Cholesterol is also important for the cytotoxic activity of K. kingae RtxA [79]. The requirement of cholesterol for toxin activity in membranes might then be another common feature shared in the family of RTX toxins.
3.1. Binding of the RTX Toxins to Membrane Cholesterol
Characterizing protein-cholesterol interactions is difficult due to cholesterol’s dual roles as a modulator of intrinsic protein function through direct binding (i.e., specific) and as an indirect (i.e., nonspecific) effector of membrane fluidity [102,103,104,105,106,107].
For a number of RTX toxins, it has been documented that the cholesterol dependence for the biological activity is sterol-specific, and it is due to direct toxin binding to the sterol molecules in the membrane and not to indirect effects of this lipid on the physical state of the phospholipid bilayer [69,79,100,108]. In 2014, Vazquez and colleagues, using different biochemical and biophysical assays, demonstrated the direct interaction of UPEC HlyA with cholesterol but not with sphingomyelin [100]. Previously, using surface plasmon resonance (SPR), Brown and colleagues determined that A. actinomycetemcomitans LtxA’s affinity for cholesterol-containing membranes was approximately four orders of magnitude greater than for cholesterol-free membranes [79]. K. kingae RtxA cytolysin was also shown to specifically bind cholesterol [69]. The authors showed that RtxA preincubated with free cholesterol exhibited significantly reduced capacity to lyse erythrocytes. They also observed a strong binding of fluorescently labeled RtxA to giant unilamellar vesicles (GUVs) composed of 75% 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) and 25% cholesterol, whereas the binding of the toxin to GUVs composed of 100% POPC was rather weak. SPR measurements then showed a stronger affinity of RtxA for the cholesterol-containing POPC membrane than for the pure POPC membrane. Moreover, RtxA bound with 2–3-fold higher efficacy to the wells of an ELISA plate coated with cholesterol-BSA than to the wells coated with free BSA, showing that the toxin is able to interact with cholesterol independently of the presence of other membrane components [66]. Our group has recently shown that pre-incubation of B. pertussis CyaA with free cholesterol notably diminishes the toxin-induced hemolysis, and that binding of CyaA to pure lipid vesicles is notably increased proportionally to the cholesterol concentration present in the lipid bilayer (10–50%) and that ergosterol (ergosta-5,7,22-trien-3β-ol), an analog of cholesterol, cannot reproduce this effect [108].
Regarding the direct interaction of RTX toxins with cholesterol, two things should be underlined. One is that the great variability in the equilibrium dissociation constant (KD) values determined by several groups (mainly using SPR) for the interaction of cholesterol with different RTX toxins, which differ in many orders of magnitude, from 10−5 to 10−12 M (Summarized in Table 1). The second point is that the values of equilibrium constants (Kd) determined for cholesterol-binding are in some cases smaller (thus, with greater affinity) than the values determined for the toxin’s association with the reported specific β2 integrin receptors. For A. actinomycetemcomitans LtxA, the maximal affinity was obtained for membranes containing 40% cholesterol using SPR with a value of KD ≈ 10−12 M, approximately four orders of magnitude greater than the affinity determined for cholesterol-free membranes (KD ≈ 10−8 M) [69]. Using Differential Scanning Calorimetry (DSC), the same group determined the KD of association of LtxA with POPC 8.75 × 10−4 M, while the KD for the interaction between LtxA and cholesterol was determined to be 2.31 × 10−10 M, which was six orders of magnitude more favorable than the affinity of LtxA for POPC. Comparatively, the KD values determined by SPR for LtxA binding to the cytoplasmic domains of both αL and β2 integrin chains were KD = 15 and 4.2 × 10−9 M, respectively, and for the cytoplasmic domains of other integrin αM, αX, and β3 subunits (KD = 400, 180, and 230 × 10−9 M, respectively), which were used as controls [109]. For K. kingae RtxA, the KD for POPC membranes was approximately 1.5 × 10−9 M, while the KD for cholesterol-containing membranes (POPC/CHOL 3:1, molar ratio) was 1.71 × 10−10 M [79]. In the case of UPEC HlyA, a KD value of 1.6 × 10−5 M was determined for the interaction with liposomes composed of DOPC/CHOL (4:1 molar ratio) [100]. Comparatively, the same group determined a higher affinity for the HlyA-glycophorin interaction (KD = 6.1 × 10−7 M) than for HlyA-CHOL [100]. Glycophorin acts as a high-affinity binding partner for HlyA in erythrocytes [110].
3.2. CRAC/CARC Motifs in the RTX Toxins as Possible Molecular Determinants of the Interaction with Membrane Cholesterol
If characterizing protein–cholesterol interaction is difficult, deciphering the exact molecular determinants of such interaction is even more complicated. As we summarize below, for several RTX toxins there is evidence indicating the involvement of cholesterol-recognizing motifs as determinants of the toxin–cholesterol interaction.
Cholesterol is an amphipathic molecule derived from the sterane backbone. Its polar section is restricted to a single hydroxyl (3β3-OH) group, which can form two distinct types of hydrogen bonds (acceptor and donor) with a polar group belonging to either a membrane lipid or a protein. The apolar section of cholesterol, in turn, has an asymmetric structure with two distinct faces, a planar α face and a β face, which has a significantly rougher surface owing to the presence of several aliphatic groups (two methyl groups and a terminal isooctyl chain that are linked to the sterane backbone) [111] (Figure 5). Overall, these structural features open up a number of possible interactions between cholesterol and membrane lipids and proteins [112].
Experimental research indicates that cholesterol exhibits a more favorable interaction with sphingomyelin than with phosphatidylcholine in model lipid bilayers [113]. This preference is attributed to the presence of a saturated acyl chain in sphingomyelin compared to a cis-unsaturated chain in phosphatidylcholine [102]. The saturated chain in sphingomyelin, coupled with the trans-unsaturated sphingosine backbone, allows for maximal van der Waals interactions with cholesterol, resulting in the formation of condensed cholesterol/sphingolipids complexes [114] adopting a specific liquid-ordered (Lo) phase [111,115]. It is important to note that while cholesterol is enriched in “nanodomains” or lipid “rafts” [116,117], it is also present outside these nanodomains in the liquid-disordered (Ld) phase of the plasma membrane, containing high amounts of glycerophospholipids like phosphatidylcholine [102]. Some authors have named this pool as “accessible cholesterol” [118]. Phosphatidylcholine, unlike sphingomyelin, possesses carbonyl groups serving as hydrogen bond acceptors but lacks hydrogen bond donor groups like the amino group in sphingomyelin. Consequently, the association between cholesterol and phosphatidylcholine relies on weakly discriminative van der Waals forces and limited hydrogen bond capabilities. In cholesterol-phosphatidylcholine complexes, both α and β faces of cholesterol [119] are potentially available for interaction with a transmembrane (TM) domain. Furthermore, the −OH group of cholesterol is not buried in the complex, remaining accessible for hydrogen bond formation with a TM domain. In contrast, cholesterol forms condensed complexes with sphingolipids (either sphingomyelin or glycosphingolipids), where the −OH group is available for stabilizing hydrogen bonds with the polar head group of the sphingolipid, limiting its initial availability for interaction with a TM domain. Sphingolipids typically interact with the α face of cholesterol, leaving the β face available for the TM domain [111]. In the case of pore-forming toxins that directly bind to membrane cholesterol, the atoms of the lipid that are accessible for binding are generally restricted to the hydroxyl group, which is available only in the Ld phase. In the Lo phase, cholesterol is totally masked by the polar head groups of sphingolipids (sphingomyelins and glycosphingolipids) through a well-characterized “umbrella effect” [120,121]. This is due to the formation of a hydrogen bond network that involves the –OH group of cholesterol. Therefore, there is little chance of the –OH group of raft-associated cholesterol being targeted by an extracellular protein [106,111].
Different types of protein structures capable of interacting directly with membrane cholesterol have been documented, either in integral membrane proteins or in numerous bacterial toxins and viral proteins that interact with cholesterol. The presence of specific cholesterol interaction sites in bacterial toxins and viral proteins reinforces the idea that such motifs could have evolved as mechanisms for selective targeting of eukaryotic membranes. Two of the best-explored cholesterol-binding motifs are the so-called cholesterol recognition/interaction amino acid consensus CRAC and the reverse CARC motifs [107,122,123] (Figure 6). The CRAC motif is a short linear segment of 5–13 amino acids that are generally present at the end of TM helices and fulfills very simple biochemical rules to associate with cholesterol. It is defined by the following algorithm in the direction N- to C-terminus: a branched apolar leucine or valine residue, followed by a segment containing 1–5 of any residue, then an aromatic residue, tyrosine, then again a segment containing 1–5 of any residues, and finally a basic lysine or arginine (L/V)-X1–5-(Y)-X1–5-(R/K) [122]. The CARC motif, in turn, corresponds to the (R/K)-X1–5-(Y/F/W)-X1–5-(L/V) pattern [107]. Unlike CRAC, the CARC motif can accept tyrosine, phenylalanine, or tryptophan as the central amino acid residue [124], and in this case, the lysine or arginine polar residues are found in the N-terminus. Nevertheless, both motifs share a similar organization, and thus, the biochemical rules that apply to the CRAC-cholesterol interactions also apply to CARC [106,125,126,127].
In the last ten years, the presence of a variable number of both CRAC and CARC motifs in the primary structure has been documented for an increasing number of RTX toxins [69,79,100,108] (see Table 2). The first RTX toxin in which CRAC/CARC cholesterol-binding sites were identified was A. actinomycetemcomitans LtxA [66]. Primary sequence analysis revealed two CRAC motifs with the pattern (L/V)-X1–5-(Y)-X1–5-(R/K): one within the pore-forming domain, the CRAC336 (333LEEYSKR339) site, which is highly conserved among RTX toxins except for B. pertussis CyaA, and a second one, upstream the hydrophobic domain, the CRAC503 (501VDYLK505) site, which is unique to LtxA [69]. If the search algorithm is broadened to also include CARC sites, one CARC motif, the CARC346 (340RFKKFGYNGDSL351) site, which is immediately posterior to the CRAC336 motif, can also be predicted in the LtxA pore-forming region (see Table 3). In 2014, Vazquez and colleagues identified twenty potential cholesterol-binding motifs, seven CRAC motifs, and thirteen CARC sites in the full-length sequence of UPEC HlyA; from them, four CRAC and three CARC sites are located in the pore-forming domain of HlyA [100]. In the case of K. kingae RtxA, five potential cholesterol-binding sites located within or adjacent to the predicted pore-forming domain (residues 140 to 410) were identified [79]. Two of them are CRAC sites, CRAC54 (48LTIPKDYDIEK58) and CRAC352 (349LAEYQR354), and three are CARC motifs, CARC285 (280KAISSYVL287), CARC343 (340KFGYDGDSL348), and CARC448 (444RHAHYLERNL453). In 2017, Masin and colleagues predicted five CRAC motifs in the pore-forming domain of the CyaA sequence, the CRAC627–638 (627VQQSHYADQLDK638), CRAC654–661 (LAQLYRDK), CRAC721–728 (721LANDYARK728), and CRAC732–741 (732LGGPQAYFEK741) sites [129]. More recently, our laboratory has identified four additional CRAC/CARC sites in the B. pertussis CyaA primary structure: two of them in two helices of the pore-forming domain, the CRAC521 and CARC532 motifs, and the other two sites, CARC415 and CRAC485, in two helices of the translocation region [108] (see Table 3 and Table 4).
Two lines of evidence have been used mainly to test the relationship and function of CRAC/CARC segments. One is to mutate the CRAC/CARC segment in the protein and determine if this alters function and/or interaction with cholesterol. An alternative strategy is to utilize a synthetic peptide corresponding to the CRAC/CARC site and test if the peptide has the ability to preferentially interact with cholesterol. Using full-length A. actinomycetemcomitans LtxA CRAC mutants (LtxA-CRACY336P and LtxA-CRACY503P), Brown and colleagues demonstrated in 2013 that from the two CRAC motifs identified, only the CRAC336 site (333LEEYSKR339) located within the pore-forming domain was essential for LtxA cytotoxicity [69]. Using short peptides corresponding to both motifs they found that both bound cholesterol, but only the peptide corresponding to the CRAC site between residues 333–339 competitively inhibited the binding of LtxA to this sterol and the ability of the toxin to kill Jurkat (Jn.9) cells [69]. The authors reported KD values of 2.31 × 10−10 M and 5.05 × 10−8 M for the LtxA-cholesterol and CRAC336 peptide-cholesterol interactions, respectively, as determined by Isothermal Titration Calorimetry (ITC) [130]. In the case of UPEC HlyA, two peptides were synthesized and analyzed, one was derived from the CARC347 site (RFKKLGYDGDSLL, residues 341–353) located in the pore-forming domain, and the other one was from the CRAC641 site (VVYYDK residues 639–644) from the domain between the two acylated lysines [131]. Using SPR and molecular dynamic (MD) simulations, the authors explored the interaction of both peptides with membranes of different lipid composition (POPC and POPC/CHOL at 4:1 and 2:1 molar ratios) and showed that both peptides interact preferentially with cholesterol-containing membranes, though the peptide harboring the CRAC641 site presented a lower KD value. In addition, only this last peptide was capable of inhibiting the HlyA-induced hemolytic activity [131]. In 2018, Osickova and colleagues replaced the key tyrosine residues of the five CRAC/CARC sites identified in the hydrophobic domain of K. kingae RtxA, namely, Y54, Y285, Y343, Y352, and Y448, with phenylalanine residues. The authors showed that the Y54F, Y285F, and Y448F substitutions did not significantly affect the lytic activity toward erythrocytes. In contrast, the Y343F and Y352F substitutions in the CARC340–348 and CRAC349–354 motifs, respectively, reduced the lytic activity to ~50% that of wild-type RtxA [79]. Regarding B-pertussis CyaA, Masin and colleagues (2017) reported that Y/F substitutions in the respective central tyrosine residues of the four CRAC motifs (Y632, Y658, Y725, and Y738) predicted in the pore-forming domain of this toxin had no effect on the translocation or the hemolytic activity of the toxin [129]. On the contrary, our laboratory has more recently found that a single F/A substitution of the central phenylalanine residue in the four CRAC/CARC sites we identified in the first two helices of the pore-forming domain and in the translocation region very notably reduces the toxin translocation capacity and affects the hemolytic potency of CyaA [108]. In sum, it appears that the use of cholesterol recognition motifs of the CRAC/CARC type as molecular determinants for directly binding this sterol in the target cell membrane is shared by several members of the RTX toxin family.
Using basic bioinformatics programs, it can be responded to the question of to what extent the functional CRAC/CARC motifs identified in the RTX toxins family are conserved and relevant for the action mechanisms. We have run the Clustal Omega program (EMBL-EBI, Cambridge, UK), a popular software for the alignment of multiple protein sequences, for several RTX toxins (see Supplementary Figure S1). From that exploration, it is confirmed that the CRAC333–339 motif firstly identified by [69] in A. actinomycetemcomitans LtxA is highly conserved among several RTX toxins, including HlyA, LktA, ApxIA and ApxIIA, MbxA, EhxA, MmxA, and RtxA, but not in the B. pertussis CyaA toxin. In addition, another two adjacently located CARC340–348 and CRAC349–359 motifs are also conserved in most of these RTX toxins, except in CyaA. The segment between residues ≈ 300–400 might thus be a kind of “hot spot” containing several conserved cholesterol-binding sites. Given that this segment is within the pore-forming domain of the respective RTX toxins, and that two hydrophobic/amphipathic α-helices can be predicted in it, which supposedly insert into the membrane, it is very tempting to speculate that cholesterol recognition through those conserved CRAC/CARC sites may be essential for proper productive insertion of those helixes into the membrane bilayer. This would explain the cholesterol dependence shown by these toxins to be cytotoxic for their target cells. Importantly, it might be a conserved mechanism shared by many RTX toxins. It is also of note that the B pertussis CyaA toxin seems to be a kind of rara avis regarding the conservation of the CRAC/CARC sites in the family. We have found that neither of the two CRAC/CARC sites (CARC413–417 and CRAC481–487) identified in the CyaA region comprising residues 400–500 (translocation region) are conserved in the rest of the RTX toxins analyzed. This may not be so strange in the end, given that this segment of the protein is involved in a function that does not exist in the rest of the RTX toxins, namely, the transport of the AC domain, and therefore could be exclusive to CyaA as believed. But more remarkable is perhaps the fact that none of the other six CRAC/CARC sites identified in the CyaA pore-forming domain [108,129] are conserved (CRAC518–527, CARC527–534, CRAC626–636, CRAC653–661, CRAC721–728, and CRAC732–741) in the rest of the RTX toxin family either (see Supplementary Figure S1). The exact meaning of the no conservation of the CRAC/CARC motifs in the CyaA sequence, and whether it may be related or not with different structural elements or different mechanisms involved in the insertion of the pore-forming domain of CyaA within the target cell membrane, is yet unknown.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms25063131/s1.
Author Contributions
H.O. wrote the paper with contributions from J.A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Spanish Ministerio de Economía y Competitividad (grant number BFU2017–82758-P). J.A. was the recipient of a fellowship from the University of Basque Country (UPV/EHU).
Data Availability Statement
Correspondence and requests for materials should be addressed to H. Ostolaza.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Welch, R.A. Pore-Forming Cytolysins of Gram-Negative Bacteria. Mol. Microbiol. 1991, 5, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Linhartová, I.; Bumba, L.; Mašín, J.; Basler, M.; Osička, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX Proteins: A Highly Diverse Family Secreted by a Common Mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.A. RTX Toxin Structure and Function: A Story of Numerous Anomalies and Few Analogies in Toxin Biology. Curr. Top. Microbiol. Immunol. 2001, 257, 85–111. [Google Scholar] [CrossRef] [PubMed]
- Coote, J.G. Structural and Functional Relationships among the RTX Toxin Determinants of Gram-Negative Bacteria. FEMS Microbiol. Rev. 1992, 8, 137–161. [Google Scholar] [CrossRef] [PubMed]
- Stanley, P.; Packman, L.C.; Koronakis, V.; Hughes, C. Fatty Acylation of Two Internal Lysine Residues Required for the Toxic Activity of Escherichia coli Hemolysin. Science 1994, 266, 1992–1996. [Google Scholar] [CrossRef] [PubMed]
- Balashova, N.V.; Shah, C.; Patel, J.K.; Megalla, S.; Kachlany, S.C. Aggregatibacter actinomycetemcomitans LtxC Is Required for Leukotoxin Activity and Initial Interaction between Toxin and Host Cells. Gene 2009, 443, 42–47. [Google Scholar] [CrossRef]
- Stanley, P.; Koronakis, V.; Hughes, C. Acylation of Escherichia coli Hemolysin: A Unique Protein Lipidation Mechanism Underlying Toxin Function. Microbiol. Mol. Biol. Rev. 1998, 62, 309–333. [Google Scholar] [CrossRef]
- Barry, E.M.; Weiss, A.A.; Ehrmann, I.E.; Gray, M.C.; Hewlett, E.L.; Goodwin, M.S. Bordetella pertussis Adenylate Cyclase Toxin and Hemolytic Activities Require a Second Gene, cyaC, for Activation. J. Bacteriol. 1991, 173, 720–726. [Google Scholar] [CrossRef]
- Osickova, A.; Khaliq, H.; Masin, J.; Jurnecka, D.; Sukova, A.; Fiser, R.; Holubova, J.; Stanek, O.; Sebo, P.; Osicka, R. Acyltransferase-Mediated Selection of the Length of the Fatty Acyl Chain and of the Acylation Site Governs Activation of Bacterial RTX Toxins. J. Biol. Chem. 2020, 295, 9268–9280. [Google Scholar] [CrossRef]
- Greene, N.P.; Crow, A.; Hughes, C.; Koronakis, V. Structure of a Bacterial Toxin-Activating Acyltransferase. Proc. Natl. Acad. Sci. USA 2015, 112, E3058–E3066. [Google Scholar] [CrossRef]
- Holland, I.B.; Schmitt, L.; Young, J. Type 1 Protein Secretion in Bacteria, the ABC-Transporter Dependent Pathway (Review). Mol. Membr. Biol. 2005, 22, 29–39. [Google Scholar] [CrossRef]
- Holland, I.B.; Peherstorfer, S.; Kanonenberg, K.; Lenders, M.; Reimann, S.; Schmitt, L. Type I Protein Secretion-Deceptively Simple yet with a Wide Range of Mechanistic Variability across the Family. EcoSal Plus 2016, 7, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Gygi, D.; Nicolet, J.; Frey, J.; Cross, M.; Koronakis, V.; Hughes, C. Isolation of the Actinobacillus pleuropneumoniae Haemolysin Gene and the Activation and Secretion of the Prohaemolysin by the HlyC, HlyB and HlyD Proteins of Escherichia coli. Mol. Microbiol. 1990, 4, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Wandersman, C.; Delepelaire, P. TolC, an Escherichia coli Outer Membrane Protein Required for Hemolysin Secretion. Proc. Natl. Acad. Sci. USA 1990, 87, 4776–4780. [Google Scholar] [CrossRef] [PubMed]
- Glaser, P.; Sakamoto, H.; Bellalou, J.; Ullmann, A.; Danchin, A. Secretion of Cyclolysin, the Calmodulin-Sensitive Adenylate Cyclase-Haemolysin Bifunctional Protein of Bordetella pertussis. EMBO J. 1988, 7, 3997–4004. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Knapp, O.; Masin, J.; Fiser, R.; Maier, E.; Benz, R.; Sebo, P.; Osicka, R. Segments Crucial for Membrane Translocation and Pore-Forming Activity of Bordetella Adenylate Cyclase Toxin. J. Biol. Chem. 2007, 282, 12419–12429. [Google Scholar] [CrossRef] [PubMed]
- Roderova, J.; Osickova, A.; Sukova, A.; Mikusova, G.; Fiser, R.; Sebo, P.; Osicka, R.; Masin, J. Residues 529 to 549 Participate in Membrane Penetration and Pore-Forming Activity of the Bordetella Adenylate Cyclase Toxin. Sci. Rep. 2019, 9, 5758. [Google Scholar] [CrossRef] [PubMed]
- Osičková, A.; Osička, R.; Maier, E.; Benz, R.; Šebo, P. An Amphipathic α-Helix Including Glutamates 509 and 516 Is Crucial for Membrane Translocation of Adenylate Cyclase Toxin and Modulates Formation and Cation Selectivity of Its Membrane Channels. J. Biol. Chem. 1999, 274, 37644–37650. [Google Scholar] [CrossRef] [PubMed]
- Valeva, A.; Walev, I.; Kemmer, H.; Weis, S.; Siegel, I.; Boukhallouk, F.; Wassenaar, T.M.; Chavakis, T.; Bhakdi, S. Binding of Escherichia coli Hemolysin and Activation of the Target Cells Is Not Receptor-Dependent. J. Biol. Chem. 2005, 280, 36657–36663. [Google Scholar] [CrossRef]
- Benz, R.; Schmid, A.; Wagner, W.; Goebel, W. Pore Formation by the Escherichia coli Hemolysin: Evidence for an Association-Dissociation Equilibrium of the Pore-Forming Aggregates. Infect. Immun. 1989, 57, 887–895. [Google Scholar] [CrossRef]
- Bhakdi, S.; Mackman, N.; Nicaud, J.M.; Holland, I.B. Escherichia coli Hemolysin May Damage Target Cell Membranes by Generating Transmembrane Pores. Infect. Immun. 1986, 52, 63–69. [Google Scholar] [CrossRef]
- Bárcena-Uribarri, I.; Benz, R.; Winterhalter, M.; Zakharian, E.; Balashova, N. Pore Forming Activity of the Potent RTX-Toxin Produced by Pediatric Pathogen Kingella kingae: Characterization and Comparison to Other RTX-Family Members. Biochim. Biophys. Acta 2015, 1848, 1536–1544. [Google Scholar] [CrossRef]
- Ludwig, A.; Schmid, A.; Benz, R.; Goebel, W. Mutations Affecting Pore Formation by Haemolysin from Escherichia coli. Mol. Gen. Genet. 1991, 226, 198–208. [Google Scholar] [CrossRef]
- Baumann, U.; Wu, S.; Flaherty, K.M.; McKay, D.B. Three-Dimensional Structure of the Alkaline Protease of Pseudomonas Aeruginosa: A Two-Domain Protein with a Calcium Binding Parallel Beta Roll Motif. EMBO J. 1993, 12, 3357–3364. [Google Scholar] [CrossRef]
- Bauche, C.; Chenal, A.; Knapp, O.; Bodenreider, C.; Benz, R.; Chaffotte, A.; Ladant, D. Structural and Functional Characterization of an Essential RTX Subdomain of Bordetella pertussis Adenylate Cyclase Toxin. J. Biol. Chem. 2006, 281, 16914–16926. [Google Scholar] [CrossRef]
- Chenal, A.; Karst, J.C.; Sotomayor Pérez, A.C.; Wozniak, A.K.; Baron, B.; England, P.; Ladant, D. Calcium-Induced Folding and Stabilization of the Intrinsically Disordered RTX Domain of the CyaA Toxin. Biophys. J. 2010, 99, 3744–3753. [Google Scholar] [CrossRef]
- Nicaud, J.-M.; Mackman, N.; Gray, L.; Holland, I.B. The C-Terminal, 23 kDa Peptide of E. coli Haemolysin 2001 Contains All the Information Necessary for Its Secretion by the Haemolysin (Hly) Export Machinery. FEBS Lett. 1986, 204, 331–335. [Google Scholar] [CrossRef]
- Mackman, N.; Baker, K.; Gray, L.; Haigh, R.; Nicaud, J.M.; Holland, I.B. Release of a Chimeric Protein into the Medium from Escherichia coli Using the C-Terminal Secretion Signal of Haemolysin. EMBO J. 1987, 6, 2835–2841. [Google Scholar] [CrossRef] [PubMed]
- Koronakis, V.; Koronakis, E.; Hughes, C. Isolation and Analysis of the C-Terminal Signal Directing Export of Escherichia coli Hemolysin Protein across Both Bacterial Membranes. EMBO J. 1989, 8, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.B.; Walker, C.R.B.; Guo, L.; Pellett, S.; Shabanowitz, J.; Hunt, D.F.; Hewlett, E.L.; Ludwig, A.; Goebel, W.; Welch, R.A.; et al. Escherichia coli α-Hemolysin (HlyA) Is Heterogeneously Acylated in Vivo with 14-, 15-, and 17-Carbon Fatty Acids. J. Biol. Chem. 2000, 275, 36698–36702. [Google Scholar] [CrossRef] [PubMed]
- Fong, K.P.; Tang, H.-Y.; Brown, A.C.; Kieba, I.R.; Speicher, D.W.; Boesze-Battaglia, K.; Lally, E.T. Aggregatibacter actinomycetemcomitans Leukotoxin Is Post-Translationally Modified by Addition of Either Saturated or Hydroxylated Fatty Acyl Chains. Mol. Oral Microbiol. 2011, 26, 262–276. [Google Scholar] [CrossRef]
- Basar, T.; Havlíček, V.; Bezoušková, S.; Hackett, M.; Šebo, P. Acylation of Lysine 983 Is Sufficient for Toxin Activity of Bordetella pertussis Adenylate Cyclase: Substitutions of Alanine 140 Modulate Acylation Site Selectivity of The Toxin Acyltransferase CyaC. J. Biol. Chem. 2001, 276, 348–354. [Google Scholar] [CrossRef]
- Nicaud, J.-M.; Mackman, N.; Gray, L.; Holland, I.B. Characterisation of HlyC and Mechanism of Activation and Secretion of Haemolysin from E. coli 2001. FEBS Lett. 1985, 187, 339–344. [Google Scholar] [CrossRef]
- Masin, J.; Osicka, R.; Bumba, L.; Sebo, P. Bordetella Adenylate Cyclase Toxin: A Unique Combination of a Pore-Forming Moiety with a Cell-Invading Adenylate Cyclase Enzyme. Pathog. Dis. 2015, 73, ftv075. [Google Scholar] [CrossRef]
- Döbereiner, A.; Schmid, A.; Ludwig, A.; Goebel, W.; Benz, R. The Effects of Calcium and Other Polyvalent Cations on Channel Formation by Escherichia coli Alpha-Hemolysin in Red Blood Cells and Lipid Bilayer Membranes. Eur. J. Biochem. 1996, 240, 454–460. [Google Scholar] [CrossRef]
- Soloaga, A.; Ostolaza, H.; Goñi, F.M.; De La Cruz, F. Purification of Escherichia coli Pro-Haemolysin, and a Comparison with the Properties of Mature α-Haemolysin. Eur. J. Biochem. 1996, 238, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Masín, J.; Konopásek, I.; Svobodová, J.; Sebo, P. Different Structural Requirements for Adenylate Cyclase Toxin Interactions with Erythrocyte and Liposome Membranes. Biochim. Biophys. Acta 2004, 1660, 144–154. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D.P.; Cannella, S.E.; Voegele, A.; Raoux-Barbot, D.; Davi, M.; Douché, T.; Matondo, M.; Brier, S.; Ladant, D.; Chenal, A.A. Post-translational Acylation Controls the Folding and Functions of the CyaA RTX Toxin. FASEB J. 2019, 33, 10065–10076. [Google Scholar] [CrossRef] [PubMed]
- Ostolaza, H.; Soloaga, A.; Goñi, F.M. The Binding of Divalent Cations to Escherichia coli Alpha-Haemolysin. Eur. J. Biochem. 1995, 228, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Karst, J.C.; Ntsogo Enguéné, V.Y.; Cannella, S.E.; Subrini, O.; Hessel, A.; Debard, S.; Ladant, D.; Chenal, A. Calcium, Acylation, and Molecular Confinement Favor Folding of Bordetella pertussis Adenylate Cyclase CyaA Toxin into a Monomeric and Cytotoxic Form. J. Biol. Chem. 2014, 289, 30702–30716. [Google Scholar] [CrossRef] [PubMed]
- Blenner, M.A.; Shur, O.; Szilvay, G.R.; Cropek, D.M.; Banta, S. Calcium-Induced Folding of a Beta Roll Motif Requires C-Terminal Entropic Stabilization. J. Mol. Biol. 2010, 400, 244–256. [Google Scholar] [CrossRef]
- Chenal, A.; Guijarro, J.I.; Raynal, B.; Delepierre, M.; Ladant, D. RTX Calcium Binding Motifs Are Intrinsically Disordered in the Absence of Calcium: Implication for Protein Secretion. J. Biol. Chem. 2009, 284, 1781–1789. [Google Scholar] [CrossRef]
- Rose, T.; Sebo, P.; Bellalou, J.; Ladant, D. Interaction of Calcium with Bordetella pertussis Adenylate Cyclase Toxin. Characterization of Multiple Calcium-Binding Sites and Calcium-Induced Conformational Changes. J. Biol. Chem. 1995, 270, 26370–26376. [Google Scholar] [CrossRef]
- Rhodes, C.R.; Gray, M.C.; Watson, J.M.; Muratore, T.L.; Kim, S.B.; Hewlett, E.L.; Grisham, C.M. Structural Consequences of Divalent Metal Binding by the Adenylyl Cyclase Toxin of Bordetella pertussis. Arch. Biochem. Biophys. 2001, 395, 169–176. [Google Scholar] [CrossRef]
- Benz, R. Channel Formation by RTX-Toxins of Pathogenic Bacteria: Basis of Their Biological Activity. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 526–537. [Google Scholar] [CrossRef]
- Hyland, C.; Vuillard, L.; Hughes, C.; Koronakis, V. Membrane Interaction of Escherichia coli Hemolysin: Flotation and Insertion-Dependent Labeling by Phospholipid Vesicles. J. Bacteriol. 2001, 183, 5364–5370. [Google Scholar] [CrossRef]
- Powthongchin, B.; Angsuthanasombat, C. Effects on Haemolytic Activity of Single Proline Substitutions in the Bordetella pertussis CyaA Pore-Forming Fragment. Arch. Microbiol. 2009, 191, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guermonprez, P.; Khelef, N.; Blouin, E.; Rieu, P.; Ricciardi-Castagnoli, P.; Guiso, N.; Ladant, D.; Leclerc, C. The Adenylate Cyclase Toxin of Bordetella pertussis Binds to Target Cells via the αMβ2 Integrin (Cd11b/Cd18). J. Exp. Med. 2001, 193, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Ristow, L.C.; Tran, V.; Schwartz, K.J.; Pankratz, L.; Mehle, A.; Sauer, J.-D.; Welch, R.A. The Extracellular Domain of the Β2 Integrin β Subunit (CD18) Is Sufficient for Escherichia coli Hemolysin and Aggregatibacter actinomycetemcomitans Leukotoxin Cytotoxic Activity. mBio 2019, 10, e01459-19. [Google Scholar] [CrossRef] [PubMed]
- Lally, E.T.; Kieba, I.R.; Sato, A.; Green, C.L.; Rosenbloom, J.; Korostoff, J.; Wang, J.F.; Shenker, B.J.; Ortlepp, S.; Robinson, M.K.; et al. RTX Toxins Recognize a Beta2 Integrin on the Surface of Human Target Cells. J. Biol. Chem. 1997, 272, 30463–30469. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bergh, P.G.A.C.; Zecchinon, L.L.M.; Fett, T.; Desmecht, D. Porcine CD18 Mediates Actinobacillus pleuropneumoniae ApxIII Species-Specific Toxicity. Vet. Res. 2009, 40, 33. [Google Scholar] [CrossRef] [PubMed]
- Confer, D.L.; Eaton, J.W. Phagocyte Impotence Caused by an Invasive Bacterial Adenylate Cyclase. Science 1982, 217, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Gueirard, P.; Le Blay, K.; Le Coustumier, A.; Chaby, R.; Guiso, N. Variation in Bordetella bronchiseptica Lipopolysaccharide during Human Infection. FEMS Microbiol. Lett. 1998, 162, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Kamanova, J.; Kofronova, O.; Masin, J.; Genth, H.; Vojtova, J.; Linhartova, I.; Benada, O.; Just, I.; Sebo, P. Adenylate Cyclase Toxin Subverts Phagocyte Function by RhoA Inhibition and Unproductive Ruffling. J. Immunol. 2008, 181, 5587–5597. [Google Scholar] [CrossRef]
- Khelef, N.; Guiso, N. Induction of Macrophage Apoptosis by Bordetella pertussis Adenylate Cyclase-Hemolysin. FEMS Microbiol. Lett. 1995, 134, 27–32. [Google Scholar] [CrossRef]
- Pearson, R.D.; Symes, P.; Conboy, M.; Weiss, A.A.; Hewlett, E.L. Inhibition of Monocyte Oxidative Responses by Bordetella pertussis Adenylate Cyclase Toxin. J. Immunol. 1987, 139, 2749–2754. [Google Scholar] [CrossRef]
- Weingart, C.L.; Weiss, A.A. Bordetella pertussis Virulence Factors Affect Phagocytosis by Human Neutrophils. Infect. Immun. 2000, 68, 1735–1739. [Google Scholar] [CrossRef]
- Dhakal, B.K.; Mulvey, M.A. The UPEC Pore-Forming Toxin α-Hemolysin Triggers Proteolysis of Host Proteins to Disrupt Cell Adhesion, Inflammatory, and Survival Pathways. Cell Host Microbe 2012, 11, 58–69. [Google Scholar] [CrossRef]
- Wiles, T.J.; Kulesus, R.R.; Mulvey, M.A. Origins and Virulence Mechanisms of Uropathogenic Escherichia coli. Exp. Mol. Pathol. 2008, 85, 11–19. [Google Scholar] [CrossRef]
- El-Azami-El-Idrissi, M.; Bauche, C.; Loucka, J.; Osicka, R.; Sebo, P.; Ladant, D.; Leclerc, C. Interaction of Bordetella pertussis Adenylate Cyclase with CD11b/CD18: Role of Toxin Acylation and Identification of the Main Integrin Interaction Domain. J. Biol. Chem. 2003, 278, 38514–38521. [Google Scholar] [CrossRef] [PubMed]
- Osicka, R.; Osickova, A.; Hasan, S.; Bumba, L.; Cerny, J.; Sebo, P. Bordetella Adenylate Cyclase Toxin Is a Unique Ligand of the Integrin Complement Receptor 3. eLife 2014, 4, e10766. [Google Scholar] [CrossRef]
- Goldsmith, J.A.; DiVenere, A.M.; Maynard, J.A.; McLellan, J.S. Structural Basis for Non-Canonical Integrin Engagement by Bordetella Adenylate Cyclase Toxin. Cell Rep. 2022, 40, 111196. [Google Scholar] [CrossRef] [PubMed]
- Nygren, P.; Balashova, N.; Brown, A.C.; Kieba, I.; Dhingra, A.; Boesze-Battaglia, K.; Lally, E.T. Aggregatibacter actinomycetemcomitans Leukotoxin Causes Activation of Lymphocyte Function-Associated Antigen 1. Cell. Microbiol. 2019, 21, e12967. [Google Scholar] [CrossRef] [PubMed]
- Ristow, L.C.; Welch, R.A. RTX Toxins Ambush Immunity’s First Cellular Responders. Toxins 2019, 11, 720. [Google Scholar] [CrossRef] [PubMed]
- Masin, J.; Osickova, A.; Jurnecka, D.; Klimova, N.; Khaliq, H.; Sebo, P.; Osicka, R. Retargeting from the CR3 to the LFA-1 Receptor Uncovers the Adenylyl Cyclase Enzyme–Translocating Segment of Bordetella Adenylate Cyclase Toxin. J. Biol. Chem. 2020, 295, 9349–9365. [Google Scholar] [CrossRef]
- Martín, C.; Requero, M.-A.; Masin, J.; Konopasek, I.; Goñi, F.M.; Sebo, P.; Ostolaza, H. Membrane Restructuring by Bordetella pertussis Adenylate Cyclase Toxin, a Member of the RTX Toxin Family. J. Bacteriol. 2004, 186, 3760–3765. [Google Scholar] [CrossRef]
- Ostolaza, H.; Bartolomé, B.; Serra, J.L.; de la Cruz, F.; Goñi, F.M. α-Haemolysin from E. coli Purification and Self-Aggregation Properties. FEBS Lett. 1991, 280, 195–198. [Google Scholar] [CrossRef]
- Bakás, L.; Veiga, M.P.; Soloaga, A.; Ostolaza, H.; Goñi, F.M. Calcium-Dependent Conformation of E. coli α-Haemolysin. Implications for the Mechanism of Membrane Insertion and Lysis. Biochim. Biophys. Acta (BBA)-Biomembr. 1998, 1368, 225–234. [Google Scholar] [CrossRef]
- Brown, A.C.; Balashova, N.V.; Epand, R.M.; Epand, R.F.; Bragin, A.; Kachlany, S.C.; Walters, M.J.; Du, Y.; Boesze-Battaglia, K.; Lally, E.T. Aggregatibacter actinomycetemcomitans Leukotoxin Utilizes a Cholesterol Recognition/Amino Acid Consensus Site for Membrane Association. J. Biol. Chem. 2013, 288, 23607–23621. [Google Scholar] [CrossRef]
- Keane, W.F.; Welch, R.; Gekker, G.; Peterson, P.K. Mechanism of Escherichia coli Alpha-Hemolysin-Induced Injury to Isolated Renal Tubular Cells. Am. J. Pathol. 1987, 126, 350–357. [Google Scholar]
- Suttorp, N.; Flöer, B.; Schnittler, H.; Seeger, W.; Bhakdi, S. Effects of Escherichia coli Hemolysin on Endothelial Cell Function. Infect. Immun. 1990, 58, 3796–3801. [Google Scholar] [CrossRef]
- Gadeberg, O.V.; Orskov, I. In Vitro Cytotoxic Effect of Alpha-Hemolytic Escherichia coli on Human Blood Granulocytes. Infect. Immun. 1984, 45, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Muhly, M.; Korom, S.; Hugo, F. Release of Interleukin-1 Beta Associated with Potent Cytocidal Action of Staphylococcal Alpha-Toxin on Human Monocytes. Infect. Immun. 1989, 57, 3512–3519. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Muhly, M.; Korom, S.; Schmidt, G. Effects of Escherichia coli Hemolysin on Human Monocytes. Cytocidal Action and Stimulation of Interleukin 1 Release. J. Clin. Investig. 1990, 85, 1746–1753. [Google Scholar] [CrossRef] [PubMed]
- Mobley, H.L.; Green, D.M.; Trifillis, A.L.; Johnson, D.E.; Chippendale, G.R.; Lockatell, C.V.; Jones, B.D.; Warren, J.W. Pyelonephritogenic Escherichia coli and Killing of Cultured Human Renal Proximal Tubular Epithelial Cells: Role of Hemolysin in Some Strains. Infect. Immun. 1990, 58, 1281–1289. [Google Scholar] [CrossRef]
- Crosby, J.A.; Kachlany, S.C. TdeA, a TolC-like Protein Required for Toxin and Drug Export in Aggregatibacter (Actinobacillus) actinomycetemcomitans. Gene 2007, 388, 83–92. [Google Scholar] [CrossRef]
- Balashova, N.V.; Crosby, J.A.; Al Ghofaily, L.; Kachlany, S.C. Leukotoxin Confers Beta-Hemolytic Activity to Actinobacillus actinomycetemcomitans. Infect. Immun. 2006, 74, 2015–2021. [Google Scholar] [CrossRef]
- Murphy, G.L.; Whitworth, L.C.; Clinkenbeard, K.D.; Clinkenbeard, P.A. Hemolytic Activity of the Pasteurella Haemolytica Leukotoxin. Infect. Immun. 1995, 63, 3209–3212. [Google Scholar] [CrossRef] [PubMed]
- Osickova, A.; Balashova, N.; Masin, J.; Sulc, M.; Roderova, J.; Wald, T.; Brown, A.C.; Koufos, E.; Chang, E.H.; Giannakakis, A.; et al. Cytotoxic Activity of Kingella kingae RtxA Toxin Depends on Post-Translational Acylation of Lysine Residues and Cholesterol Binding. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef]
- Kehl-Fie, T.E.; St. Geme, J.W. Identification and Characterization of an RTX Toxin in the Emerging Pathogen Kingella kingae. J. Bacteriol. 2007, 189, 430–436. [Google Scholar] [CrossRef]
- Ostolaza, H.; Bartolomé, B.; de Zárate, I.O.; de la Cruz, F.; Goñi, F.M. Release of Lipid Vesicle Contents by the Bacterial Protein Toxin α-Haemolysin. Biochim. Biophys. Acta (BBA)-Biomembr. 1993, 1147, 81–88. [Google Scholar] [CrossRef]
- Brown, A.C.; Boesze-Battaglia, K.; Du, Y.; Stefano, F.P.; Kieba, I.R.; Epand, R.F.; Kakalis, L.; Yeagle, P.L.; Epand, R.M.; Lally, E.T. Aggregatibacter actinomycetemcomitans Leukotoxin Cytotoxicity Occurs through Bilayer Destabilization. Cell. Microbiol. 2012, 14, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Fiser, R.; Konopásek, I. Different Modes of Membrane Permeabilization by Two RTX Toxins: HlyA from Escherichia coli and CyaA from Bordetella pertussis. Biochim. Biophys. Acta 2009, 1788, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Bakás, L.; Ostolaza, H.; Vaz, W.L.; Goñi, F.M. Reversible Adsorption and Nonreversible Insertion of Escherichia coli Alpha-Hemolysin into Lipid Bilayers. Biophys. J. 1996, 71, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Ostolaza, H.; Bakás, L.; Goñi, F.M. Balance of Electrostatic and Hydrophobic Interactions in the Lysis of Model Membranes by E. coliα-Haemolysin. J. Membr. Biol. 1997, 158, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Iwase, M.; Lally, E.T.; Berthold, P.; Korchak, H.M.; Taichman, N.S. Effects of Cations and Osmotic Protectants on Cytolytic Activity of Actinobacillus actinomycetemcomitans Leukotoxin. Infect. Immun. 1990, 58, 1782–1788. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Magraner, L.; Viguera, A.R.; García-Pacios, M.; Garcillán, M.P.; Arrondo, J.-L.R.; de la Cruz, F.; Goñi, F.M.; Ostolaza, H. The Calcium-Binding C-Terminal Domain of Escherichia coli α-Hemolysin Is a Major Determinant in the Surface-Active Properties of the Protein. J. Biol. Chem. 2007, 282, 11827–11835. [Google Scholar] [CrossRef]
- Forman, M.S.; Nishikubo, J.B.; Han, R.K.; Le, A.; Balashova, N.V.; Kachlany, S.C. Gangliosides Block Aggregatibacter actinomycetemcomitans Leukotoxin (LtxA)-Mediated Hemolysis. Toxins 2010, 2, 2824–2836. [Google Scholar] [CrossRef] [PubMed]
- Munksgaard, P.S.; Skals, M.; Reinholdt, J.; Poulsen, K.; Jensen, M.R.; Yang, C.; Leipziger, J.; Vorup-Jensen, T.; Praetorius, H.A. Sialic Acid Residues Are Essential for Cell Lysis Mediated by Leukotoxin from Aggregatibacter actinomycetemcomitans. Infect. Immun. 2014, 82, 2219–2228. [Google Scholar] [CrossRef] [PubMed]
- Morova, J.; Osicka, R.; Masin, J.; Sebo, P. RTX Cytotoxins Recognize Beta2 Integrin Receptors through N-Linked Oligosaccharides. Proc. Natl. Acad. Sci. USA 2008, 105, 5355–5360. [Google Scholar] [CrossRef]
- Gable, P.; Eaton, J.; Confer, D. Intoxication of Human Phagocytes by Bordetella Adenylate-Cyclase Toxin-Implication of a Ganglioside Receptor. Clin. Res. 1985, 33, A844. [Google Scholar]
- Gordon, V.M.; Young, W.W.; Lechler, S.M.; Gray, M.C.; Leppla, S.H.; Hewlett, E.L. Adenylate Cyclase Toxins from Bacillus anthracis and Bordetella pertussis. J. Biol. Chem. 1989, 264, 14792–14796. [Google Scholar] [CrossRef]
- Mrówczyńska, L.; Bobrowska-Hägerstrand, M.; Lindqvist, C.; Hägerstrand, H. Bordetella Adenylate Cyclase Toxin Can Bind Ganglioside GM1. BIO 2011, 1, 67–71. [Google Scholar] [CrossRef]
- Hasan, S.; Osickova, A.; Bumba, L.; Novák, P.; Sebo, P.; Osicka, R. Interaction of Bordetella Adenylate Cyclase Toxin with Complement Receptor 3 Involves Multivalent Glycan Binding. FEBS Lett. 2015, 589, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Rahman, W.U.; Osickova, A.; Klimova, N.; Lora, J.; Balashova, N.; Osicka, R. Binding of Kingella kingae RtxA Toxin Depends on Cell Surface Oligosaccharides, but Not on Β2 Integrins. Int. J. Mol. Sci. 2020, 21, 9092. [Google Scholar] [CrossRef]
- Fong, K.P.; Pacheco, C.M.F.; Otis, L.L.; Baranwal, S.; Kieba, I.R.; Harrison, G.; Hersh, E.V.; Boesze-Battaglia, K.; Lally, E.T. Actinobacillus actinomycetemcomitans Leukotoxin Requires Lipid Microdomains for Target Cell Cytotoxicity. Cell Microbiol. 2006, 8, 1753–1767. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.; Koufos, E.; Balashova, N.V.; Boesze-Battaglia, K.; Lally, E.T. Inhibition of LtxA Toxicity by Blocking Cholesterol Binding with Peptides. Mol. Oral Microbiol. 2016, 31, 94–105. [Google Scholar] [CrossRef]
- Bumba, L.; Masin, J.; Fiser, R.; Sebo, P. Bordetella Adenylate Cyclase Toxin Mobilizes Its Beta2 Integrin Receptor into Lipid Rafts to Accomplish Translocation across Target Cell Membrane in Two Steps. PLoS Pathog. 2010, 6, e1000901. [Google Scholar] [CrossRef] [PubMed]
- Herlax, V.; Maté, S.; Rimoldi, O.; Bakás, L. Relevance of Fatty Acid Covalently Bound to Escherichia coli α-Hemolysin and Membrane Microdomains in the Oligomerization Process. J. Biol. Chem. 2009, 284, 25199–25210. [Google Scholar] [CrossRef]
- Vazquez, R.F.; Maté, S.M.; Bakás, L.S.; Fernández, M.M.; Malchiodi, E.L.; Herlax, V.S. Novel Evidence for the Specific Interaction between Cholesterol and α-Haemolysin of Escherichia coli. Biochem. J. 2014, 458, 481–489. [Google Scholar] [CrossRef]
- González Bullón, D.; Uribe, K.B.; Amuategi, J.; Martín, C.; Ostolaza, H. Cholesterol Stimulates the Lytic Activity of Adenylate Cyclase Toxin on Lipid Membranes by Promoting Toxin Oligomerization and Formation of Pores with a Greater Effective Size. FEBS J. 2021, 288, 6795–6814. [Google Scholar] [CrossRef]
- Fantini, J.; Garmy, N.; Mahfoud, R.; Yahi, N. Lipid Rafts: Structure, Function and Role in HIV, Alzheimer’s and Prion Diseases. Expert. Rev. Mol. Med. 2002, 4, 1–22. [Google Scholar] [CrossRef]
- Chang, H.M.; Reitstetter, R.; Mason, R.P.; Gruener, R. Attenuation of Channel Kinetics and Conductance by Cholesterol: An Interpretation Using Structural Stress as a Unifying Concept. J. Membr. Biol. 1995, 143, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Cherezov, V.; Griffith, M.T.; Roth, C.B.; Jaakola, V.-P.; Chien, E.Y.T.; Velasquez, J.; Kuhn, P.; Stevens, R.C. A Specific Cholesterol Binding Site Is Established by the 2.8 A Structure of the Human Beta2-Adrenergic Receptor. Structure 2008, 16, 897–905. [Google Scholar] [CrossRef]
- Rosenhouse-Dantsker, A.; Noskov, S.; Durdagi, S.; Logothetis, D.E.; Levitan, I. Identification of Novel Cholesterol-Binding Regions in Kir2 Channels. J. Biol. Chem. 2013, 288, 31154–31164. [Google Scholar] [CrossRef]
- Fantini, J.; Di Scala, C.; Baier, C.J.; Barrantes, F.J. Molecular Mechanisms of Protein-Cholesterol Interactions in Plasma Membranes: Functional Distinction between Topological (Tilted) and Consensus (CARC/CRAC) Domains. Chem. Phys. Lipids 2016, 199, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Baier, C.J.; Fantini, J.; Barrantes, F.J. Disclosure of Cholesterol Recognition Motifs in Transmembrane Domains of the Human Nicotinic Acetylcholine Receptor. Sci. Rep. 2011, 1, 69. [Google Scholar] [CrossRef]
- Amuategi, J.; Alonso, R.; Ostolaza, H. Four Cholesterol-Recognition Motifs in the Pore-Forming and Translocation Domains of Adenylate Cyclase Toxin Are Essential for Invasion of Eukaryotic Cells and Lysis of Erythrocytes. Int. J. Mol. Sci. 2022, 23, 8703. [Google Scholar] [CrossRef] [PubMed]
- Krueger, E.; Brown, A.C. Aggregatibacter actinomycetemcomitans Leukotoxin: From Mechanism to Targeted Anti-Toxin Therapeutics. Mol. Oral Microbiol. 2020, 35, 85–105. [Google Scholar] [CrossRef]
- Cortajarena, A.L.; Goñi, F.M.; Ostolaza, H. Glycophorin as a Receptor for Escherichia coli α-Hemolysin in Erythrocytes. J. Biol. Chem. 2001, 276, 12513–12519. [Google Scholar] [CrossRef]
- Fantini, J.; Barrantes, F.J. Sphingolipid/Cholesterol Regulation of Neurotransmitter Receptor Conformation and Function. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 2345–2361. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Chattopadhyay, A. Transbilayer Organization of Membrane Cholesterol at Low Concentrations: Implications in Health and Disease. Biochim. Biophys. Acta (BBA)-Biomembr. 2011, 1808, 19–25. [Google Scholar] [CrossRef]
- Mattjus, P.; Slotte, J.P. Does Cholesterol Discriminate between Sphingomyelin and Phosphatidylcholine in Mixed Monolayers Containing Both Phospholipids? Chem. Phys. Lipids 1996, 81, 69–80. [Google Scholar] [CrossRef]
- Radhakrishnan, A.; Anderson, T.G.; McConnell, H.M. Condensed Complexes, Rafts, and the Chemical Activity of Cholesterol in Membranes. Proc. Natl. Acad. Sci. USA 2000, 97, 12422–12427. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; London, E. Structure and Origin of Ordered Lipid Domains in Biological Membranes. J. Membr. Biol. 1998, 164, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ikonen, E. Functional Rafts in Cell Membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.G.W.; Jacobson, K. A Role for Lipid Shells in Targeting Proteins to Caveolae, Rafts, and Other Lipid Domains. Science 2002, 296, 1821–1825. [Google Scholar] [CrossRef] [PubMed]
- Infante, R.E.; Abi-Mosleh, L.; Radhakrishnan, A.; Dale, J.D.; Brown, M.S.; Goldstein, J.L. Purified NPC1 Protein. I. Binding of Cholesterol and Oxysterols to a 1278-Amino Acid Membrane Protein. J. Biol Chem 2008, 283, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Rose, I.A.; Hanson, K.R.; Wilkinson, K.D.; Wimmer, M.J. A Suggestion for Naming Faces of Ring Compounds. Proc. Natl. Acad. Sci. USA 1980, 77, 2439–2441. [Google Scholar] [CrossRef]
- Ramstedt, B.; Slotte, J.P. Interaction of Cholesterol with Sphingomyelins and Acyl-Chain-Matched Phosphatidylcholines: A Comparative Study of the Effect of the Chain Length. Biophys. J. 1999, 76, 908–915. [Google Scholar] [CrossRef]
- Fantini, J.; Yahi, N.; Garmy, N. Cholesterol Accelerates the Binding of Alzheimer’s β-Amyloid Peptide to Ganglioside GM1 through a Universal Hydrogen-Bond-Dependent Sterol Tuning of Glycolipid Conformation. Front. Physiol. 2013, 4, 120. [Google Scholar] [CrossRef]
- Li, H.; Papadopoulos, V. Peripheral-Type Benzodiazepine Receptor Function in Cholesterol Transport. Identification of a Putative Cholesterol Recognition/Interaction Amino Acid Sequence and Consensus Pattern. Endocrinology 1998, 139, 4991–4997. [Google Scholar] [CrossRef]
- Jamin, N.; Neumann, J.-M.; Ostuni, M.A.; Vu, T.K.N.; Yao, Z.-X.; Murail, S.; Robert, J.-C.; Giatzakis, C.; Papadopoulos, V.; Lacapère, J.-J. Characterization of the Cholesterol Recognition Amino Acid Consensus Sequence of the Peripheral-Type Benzodiazepine Receptor. Mol. Endocrinol. 2005, 19, 588–594. [Google Scholar] [CrossRef]
- Ulmschneider, M.B.; Sansom, M.S. Amino Acid Distributions in Integral Membrane Protein Structures. Biochim. Biophys. Acta 2001, 1512, 1–14. [Google Scholar] [CrossRef]
- Fantini, J.; Barrantes, F.J. How Cholesterol Interacts with Membrane Proteins: An Exploration of Cholesterol-Binding Sites Including CRAC, CARC, and Tilted Domains. Front. Physiol. 2013, 4, 31. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Di Scala, C.; Evans, L.S.; Williamson, P.T.F.; Barrantes, F.J. A Mirror Code for Protein-Cholesterol Interactions in the Two Leaflets of Biological Membranes. Sci. Rep. 2016, 6, 21907. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Epand, R.M.; Barrantes, F.J. Cholesterol-Recognition Motifs in Membrane Proteins. Adv. Exp. Med. Biol. 2019, 1135, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Azzaz, F.; Chahinian, H.; Yahi, N.; Di Scala, C.; Baier, C.J.; Barrantes, F.J.; Fantini, J. Chapter 7—Cholesterol-Recognizing Amino Acid Consensus Motifs in Transmembrane Proteins: Comparative Analysis of in Silico Studies and Structural Data. In Cholesterol; Bukiya, A.N., Dopico, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 127–145. ISBN 978-0-323-85857-1. [Google Scholar]
- Masin, J.; Roderova, J.; Osickova, A.; Novak, P.; Bumba, L.; Fiser, R.; Sebo, P.; Osicka, R. The Conserved Tyrosine Residue 940 Plays a Key Structural Role in Membrane Interaction of Bordetella Adenylate Cyclase Toxin. Sci. Rep. 2017, 7, 9330. [Google Scholar] [CrossRef] [PubMed]
- Koufos, E.; Chang, E.H.; Rasti, E.S.; Krueger, E.; Brown, A.C. Use of a Cholesterol Recognition Amino Acid Consensus Peptide to Inhibit Binding of a Bacterial Toxin to Cholesterol. Biochemistry 2016, 55, 4787–4797. [Google Scholar] [CrossRef] [PubMed]
- Cané, L.; Guzmán, F.; Balatti, G.; Daza Millone, M.A.; Pucci Molineris, M.; Maté, S.; Martini, M.F.; Herlax, V. Biophysical Analysis to Assess the Interaction of CRAC and CARC Motif Peptides of Alpha Hemolysin of Escherichia coli with Membranes. Biochemistry 2023, 62, 1994–2011. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
General scheme of the synthesis, post-translational modification, and secretion for the RTX toxins. The schematic organization of the operon is represented by boxes labelled from A to D. Gene product A (red) is the polypeptide corresponding to a protoxin (pro-RTX) that matures in the bacterial cytosol to the active form by post-translational acylation at two conserved internal lysine residues [5]. Fatty acylation is mediated by a specific acyltransferase encoded by the product of the gene C (dark green) and an acyl carrier protein (Acyl-ACP) [5,6,7,8,9,10]. The mature, acylated RTX toxin is then directly secreted across both membranes by the type I secretion system (T1SS) constituted by the gene products B (light green) and D (pink) and the bacterial outer membrane TolC protein [11,12,13,14,15]. Adapted from Stanley and cols. [7]. Created with Biorender.com (accessed on 10 January 2024).
Figure 1.
General scheme of the synthesis, post-translational modification, and secretion for the RTX toxins. The schematic organization of the operon is represented by boxes labelled from A to D. Gene product A (red) is the polypeptide corresponding to a protoxin (pro-RTX) that matures in the bacterial cytosol to the active form by post-translational acylation at two conserved internal lysine residues [5]. Fatty acylation is mediated by a specific acyltransferase encoded by the product of the gene C (dark green) and an acyl carrier protein (Acyl-ACP) [5,6,7,8,9,10]. The mature, acylated RTX toxin is then directly secreted across both membranes by the type I secretion system (T1SS) constituted by the gene products B (light green) and D (pink) and the bacterial outer membrane TolC protein [11,12,13,14,15]. Adapted from Stanley and cols. [7]. Created with Biorender.com (accessed on 10 January 2024).

Figure 2.
Schematic representation of the structure of the RTX toxins UPEC HlyA (top) and B. pertussis CyaA (bottom). These toxins consist of a pore-forming domain (H, dark blue), an acylated segment with two post-translationally acylated lysine residues (indicated with two turquois arrows), a repeat domain (RD dark green), a receptor binding domain (RBD, light blue), and a C-terminal secretion signal (SS, purple). Unlike other RTX toxins, CyaA contains a unique adenylate cyclase AC domain (AC, green) and a translocation region (TR, orange).
Figure 2.
Schematic representation of the structure of the RTX toxins UPEC HlyA (top) and B. pertussis CyaA (bottom). These toxins consist of a pore-forming domain (H, dark blue), an acylated segment with two post-translationally acylated lysine residues (indicated with two turquois arrows), a repeat domain (RD dark green), a receptor binding domain (RBD, light blue), and a C-terminal secretion signal (SS, purple). Unlike other RTX toxins, CyaA contains a unique adenylate cyclase AC domain (AC, green) and a translocation region (TR, orange).
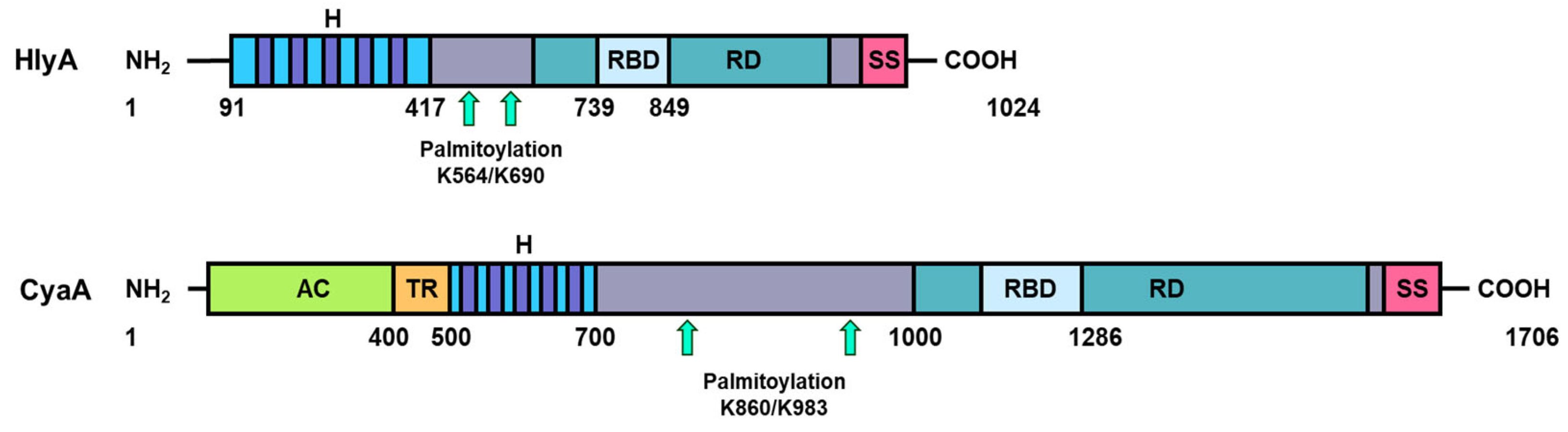
Figure 3.
Structure of the αMβ2 integrin ectodomain. Structure at 2.7 Å resolution of the heterodimeric αMβ2 integrin published by Goldsmith and cols [62], showing the αM subunit in yellow and the β2 subunit in red. A cartoon representation of the cell membrane lipid bilayer with phospholipid molecules (in purple) and cholesterol molecules (in orange) has been included as well. Figure redrawn from the integrin structure solved by Goldsmith and cols and deposited in PDB ID: 7USL [62].
Figure 3.
Structure of the αMβ2 integrin ectodomain. Structure at 2.7 Å resolution of the heterodimeric αMβ2 integrin published by Goldsmith and cols [62], showing the αM subunit in yellow and the β2 subunit in red. A cartoon representation of the cell membrane lipid bilayer with phospholipid molecules (in purple) and cholesterol molecules (in orange) has been included as well. Figure redrawn from the integrin structure solved by Goldsmith and cols and deposited in PDB ID: 7USL [62].
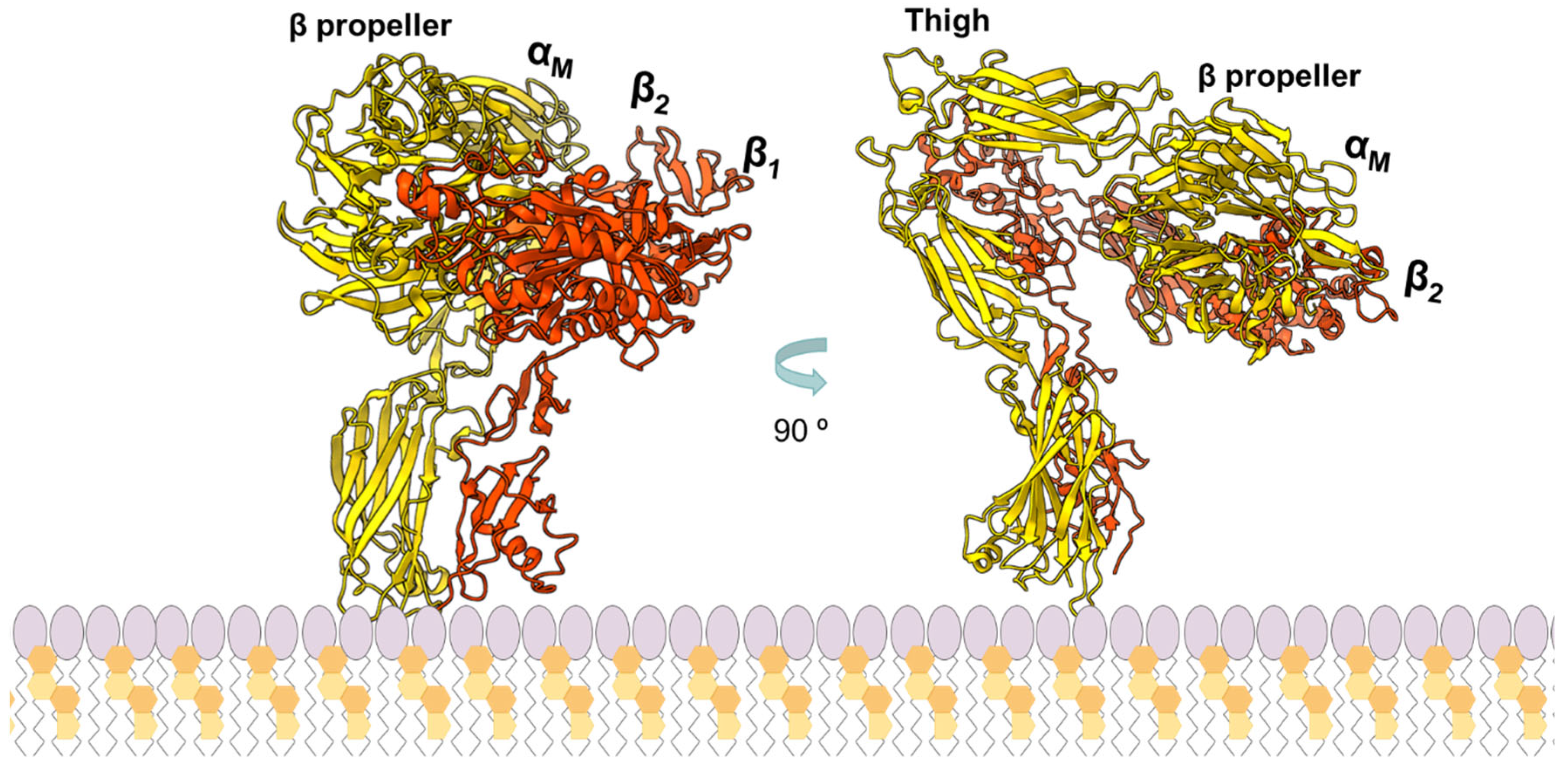
Figure 4.
Schematic representation of the chemical structure of GM1 and GM3 gangliosides.
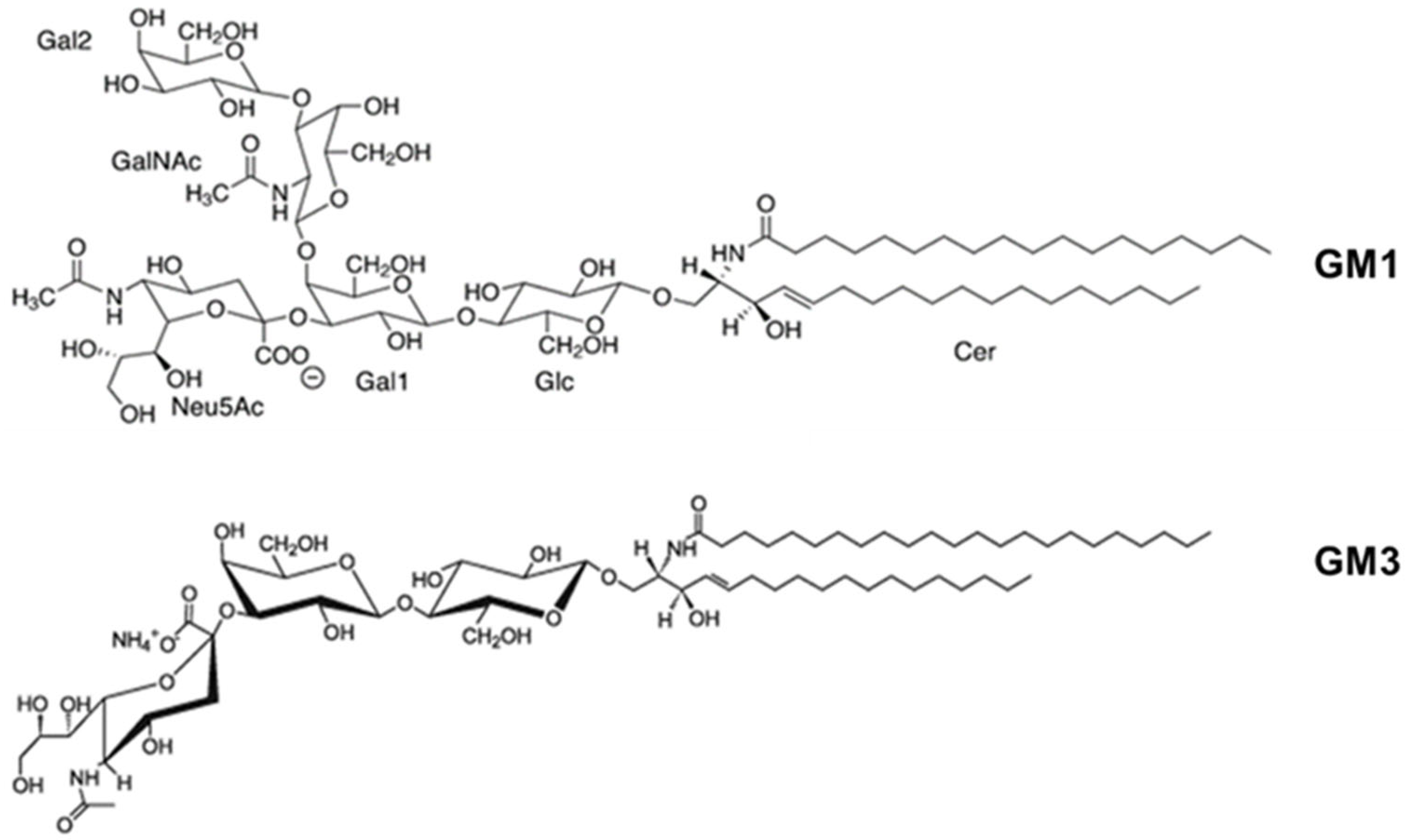
Figure 5.
Chemical structure of cholesterol. Cholesterol has a global conical shape, and it can be divided into two parts. On the one hand, the 4-ring sterane system with the linked hydroxyl group occupies 50% of the spatial volume of the molecule, and it has very low flexibility; on the other, the terminal isooctyl chain is very flexible and, hence, it can adopt numerous conformations when bound to membrane proteins. The combination of polar (3β-hydroxyl group) and apolar (the sterol ring and the isooctyl side chain) regions impart an amphipathic nature to cholesterol, making it conducive to interaction with other membrane components (lipids and proteins). An interesting structural feature of cholesterol is the inherent asymmetry of the sterol ring plane owing to methyl substitutions on one of its faces. The smooth α face is constituted of only axial hydrogen atoms and contributes to favorable van der Waals interaction with the saturated fatty acyl chains of phospholipids. On the other hand, the rough β face characterized by the protruding methyl groups (C18 and C19) can snugly interact with the bumpy topology of a membrane protein.
Figure 5.
Chemical structure of cholesterol. Cholesterol has a global conical shape, and it can be divided into two parts. On the one hand, the 4-ring sterane system with the linked hydroxyl group occupies 50% of the spatial volume of the molecule, and it has very low flexibility; on the other, the terminal isooctyl chain is very flexible and, hence, it can adopt numerous conformations when bound to membrane proteins. The combination of polar (3β-hydroxyl group) and apolar (the sterol ring and the isooctyl side chain) regions impart an amphipathic nature to cholesterol, making it conducive to interaction with other membrane components (lipids and proteins). An interesting structural feature of cholesterol is the inherent asymmetry of the sterol ring plane owing to methyl substitutions on one of its faces. The smooth α face is constituted of only axial hydrogen atoms and contributes to favorable van der Waals interaction with the saturated fatty acyl chains of phospholipids. On the other hand, the rough β face characterized by the protruding methyl groups (C18 and C19) can snugly interact with the bumpy topology of a membrane protein.
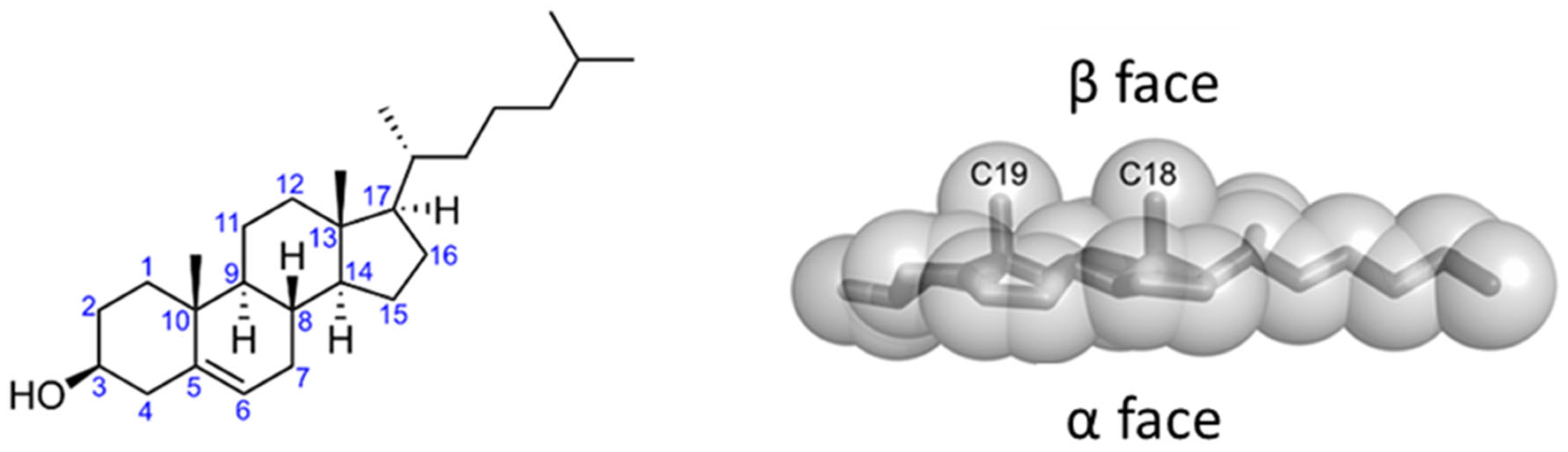
Figure 6.
Cholesterol recognition/interaction amino acid consensus motifs. (A) Mirror topology of CRAC/CARC motifs within the same TM domain of a multispanning membrane-protein. The dashed line indicates the border between the inner and outer leaflets. Many cholesterol-binding proteins possess an amino acid sequence in the juxtamembrane region conforming to the pattern (L/V)-X1–5-(Y)-X1–5-(K/R) (CRAC) or the opposite (K/R)-X1–5-(Y/F/W)-X1–5-(L/V) (CARC). (B) Schematic illustration of the interaction between the cholesterol molecule and particular amino acid residues corresponding to CRAC/CARC-like motifs. The CRAC/cholesterol complex displays a parallel head-to-head/tail-to-tail geometry [127,128]. The branched N-terminal Leu or Val apolar residues bind to the isooctyl chain through van der Waals interactions (London forces); the mandatory aromatic residue (Tyr, Phe, or Trp) stacks onto one of the steranes four rings (CH-π stacking); and the C-terminal Lys or Arg polar residues establish hydrogen bonds with the hydroxyl group of CHOL [127]. The aromatic residues are able to stack onto the smooth α face of cholesterol or intercalate between the aliphatic spikes that emerge from the rough β face. The position of the aromatic residue is determined by the length of the X1–5 linkers [127,128]. Redrawn from Fantini and colleagues [127].
Figure 6.
Cholesterol recognition/interaction amino acid consensus motifs. (A) Mirror topology of CRAC/CARC motifs within the same TM domain of a multispanning membrane-protein. The dashed line indicates the border between the inner and outer leaflets. Many cholesterol-binding proteins possess an amino acid sequence in the juxtamembrane region conforming to the pattern (L/V)-X1–5-(Y)-X1–5-(K/R) (CRAC) or the opposite (K/R)-X1–5-(Y/F/W)-X1–5-(L/V) (CARC). (B) Schematic illustration of the interaction between the cholesterol molecule and particular amino acid residues corresponding to CRAC/CARC-like motifs. The CRAC/cholesterol complex displays a parallel head-to-head/tail-to-tail geometry [127,128]. The branched N-terminal Leu or Val apolar residues bind to the isooctyl chain through van der Waals interactions (London forces); the mandatory aromatic residue (Tyr, Phe, or Trp) stacks onto one of the steranes four rings (CH-π stacking); and the C-terminal Lys or Arg polar residues establish hydrogen bonds with the hydroxyl group of CHOL [127]. The aromatic residues are able to stack onto the smooth α face of cholesterol or intercalate between the aliphatic spikes that emerge from the rough β face. The position of the aromatic residue is determined by the length of the X1–5 linkers [127,128]. Redrawn from Fantini and colleagues [127].
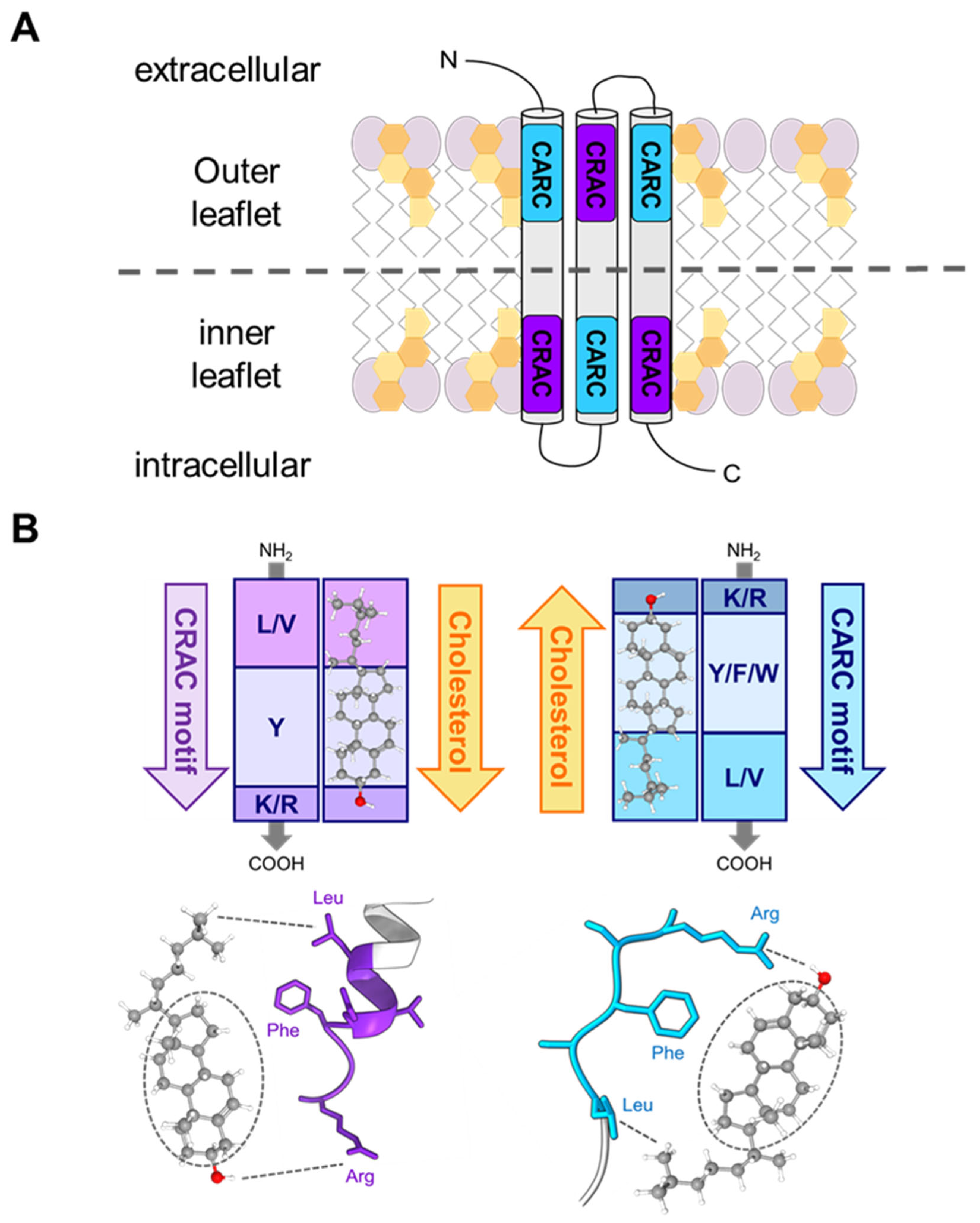

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the binding affinities of distinct RTX toxins to cholesterol. The RTX toxin and the interaction affinity to certain lipid compositions measured either with SPR or DSC are shown.
Table 1.
Summary of the binding affinities of distinct RTX toxins to cholesterol. The RTX toxin and the interaction affinity to certain lipid compositions measured either with SPR or DSC are shown.
| RTX Toxin | Lipid Composition | Affinity (M) | Measurement Technique | Reference |
|---|---|---|---|---|
| A. Actinomycetemcomitans LtxA | POPC/CHOL (3:2 molar ratio) | KD = 10−12 | SPR | Brown and colleagues [69] |
| CHOL | KD = 2.31 × 10−10 | DSC | Krueger and Brown [106] | |
| K. kingae RtxA | POPC/CHOL (3:1 molar ratio) | KD = 1.71 × 10−10 | SPR | Osickova and colleagues [79] |
| UPEC HlyA | DOPC/CHOL (4:1 molar ratio) | KD = 1.6 × 10−5 | SPR | Vazquez and colleagues [100] |
Table 2.
Summary of the total number of CRAC/CARC motifs identified in the primary structure of RTX toxins. Searching input algorithms used with the EMBOSS explorer FuzzPro software (https://www.bioinformatics.nl/cgi-bin/emboss/fuzzpro, (accessed on 5 March 2024), Alan Bleasby, EBI, Cambridge, UK) were as follows: [L/V]-X1–5-[Y/F]-X1–5-[K/R] for CRAC and [K/R]-X1–5-[Y/F]-X1–5-[L/V] for CARC motifs, respectively.
Table 2.
Summary of the total number of CRAC/CARC motifs identified in the primary structure of RTX toxins. Searching input algorithms used with the EMBOSS explorer FuzzPro software (https://www.bioinformatics.nl/cgi-bin/emboss/fuzzpro, (accessed on 5 March 2024), Alan Bleasby, EBI, Cambridge, UK) were as follows: [L/V]-X1–5-[Y/F]-X1–5-[K/R] for CRAC and [K/R]-X1–5-[Y/F]-X1–5-[L/V] for CARC motifs, respectively.
| RTX Toxin | Bacterium | Residues | CRAC Motifs | CARC Motifs | Total No. CRAC/CARC Motifs |
|---|---|---|---|---|---|
| EhxA | Enterohemorrhagic (EHEC) Escherichia coli | 997 | 16 | 15 | 31 |
| LktA | Mannheimia haemolytica | 953 | 15 | 13 | 28 |
| PlLktA | M. varigena | 953 | 21 | 16 | 37 |
| PaxA | Pasteurella aerogenes | 1049 | 19 | 21 | 40 |
| MmxA | Morganella morganii | 1024 | 13 | 15 | 28 |
| HlyA | UPEC E. coli | 1023 | 13 | 13 | 26 |
| CyaA | Bordetella pertussis | 1706 | 22 | 18 | 40 |
| LtxA | Aggregatibacter actinomycetemcomitans | 1055 | 24 | 20 | 44 |
| ApxIA | Actinobacillus pleuropneumoniae | 1022 | 18 | 16 | 34 |
| ApxIIA | A. pleuropneumoniae | 956 | 16 | 16 | 32 |
| ApxIIIA | A. pleuropneumoniae | 1052 | 22 | 19 | 41 |
| MbxA | Moraxella bovis | 927 | 25 | 18 | 43 |
| RtxA | Kingella kingae | 956 | 12 | 14 | 26 |
Table 3.
CARC motifs identified in the sequences corresponding to the pore-forming domain of various RTX toxins. The searching input algorithm used with the EMBOSS explorer FuzzPro software (https://www.bioinformatics.nl/cgi-bin/emboss/fuzzpro, (accessed on 5 March 2024), Alan Bleasby, EBI, Cambridge, UK) was [K/R]-X1–5-[Y/F]-X1–5-[L/V]. The location of the motifs in the full-length toxin and their corresponding amino acid sequence are shown. Central phenylalanine and tyrosine residues are labeled in red.
Table 3.
CARC motifs identified in the sequences corresponding to the pore-forming domain of various RTX toxins. The searching input algorithm used with the EMBOSS explorer FuzzPro software (https://www.bioinformatics.nl/cgi-bin/emboss/fuzzpro, (accessed on 5 March 2024), Alan Bleasby, EBI, Cambridge, UK) was [K/R]-X1–5-[Y/F]-X1–5-[L/V]. The location of the motifs in the full-length toxin and their corresponding amino acid sequence are shown. Central phenylalanine and tyrosine residues are labeled in red.
| Motif | RTX Toxin | Bacterium | Residues | Sequence |
|---|---|---|---|---|
| CARC | EhxA | EHEC E. coli | 434–443 | RHAAFLEDSL |
| 400–407 | KQAMFEHV | |||
| 328–340 | RFKKLNYEGDALL | |||
| 271–278 | KAVSQYIL | |||
| 102–110 | KLLQKYQKV | |||
| LktA | M. haemolytica | 405–412 | KQAMFEHV | |
| 333–345 | RFKKLGYDGDNLL | |||
| 276–287 | KAVSSYILAQRV | |||
| 259–270 | KVGAGFELANQV | |||
| PlLktA | M. varigena | 405–412 | KQAMFEHV | |
| 333–345 | RFKKLGYDGDDLL | |||
| 276–287 | KAVSSYILAQRV | |||
| 258–270 | RKVGAGFELVNQV | |||
| 221–231 | KNFSGFSKAGL | |||
| 199–207 | KINQFGSKL | |||
| PaxA | P. aerogenes | 455–464 | RHKAFLEDSL | |
| 349–361 | RFKKLGYEGDKLL | |||
| 344–353 | REFAERFKKL | |||
| 292–303 | KAVSSYILAQRL | |||
| 275–286 | KVAAGFELSNQV | |||
| 115–124 | RGLTLFAPQL | |||
| MmxA | M. morganii | 458–469 | KLLSKYSEEYSV | |
| 414–421 | KQAMFEHV | |||
| 342–354 | RFKKFGYEGDSLL | |||
| 285–296 | KAVSQYILAQRV | |||
| 105–114 | RGIAIFAPQL | |||
| HlyA | UPEC E. coli | 457–468 | KILSQYNKEYSV | |
| 413–420 | KQAMFEHV | |||
| 341–353 | RFKKLGYDGDSLL | |||
| 105–114 | RGVTIFAPQL | |||
| CyaA | B. pertussis | 527–534 | RWAGGFGV | |
| 413–420 | RSFSLGEV | |||
| 399–410 | RQDSGYDSLDGV | |||
| LtxA | A. actinomycetemcomitans | 456–461 | KLFNEL | |
| 446–455 | RHSAFLEDSL | |||
| 340–352 | RFKKFGYNGDSLL | |||
| 326–334 | KQFDRARML | |||
| 218–227 | KHFGSFGDKL | |||
| ApxIA | A. pleuropneumoniae | 461–469 | KEYSVERVV | |
| 410–417 | KQAIFERV | |||
| 338–350 | RFKKFGYEGDSLL | |||
| 281–292 | KAVSQYIIAQRV | |||
| ApxIIA | A. pleuropneumoniae | 410–417 | KQAMFEHV | |
| 338–350 | RFQKLGYDGDRLL | |||
| 333–342 | KSYSERFQKL | |||
| 281–292 | KAVSSYILAQRV | |||
| ApxIIIA | A. pleuropneumoniae | 352–360 | KLGYDGDKL | |
| 349–361 | RFKKLGYDGDKLL | |||
| 292–303 | KAVSSYILAQRL | |||
| 275–286 | KVAAGFELSNQV | |||
| 115–124 | RGLTLFAPQL | |||
| MbxA | M. bovis | 422–431 | RYAAYLANNL | |
| 417–427 | KGYDSRYAAYL | |||
| 387–394 | KQAMFESV | |||
| 317–327 | RKFGYDGDHLL | |||
| 258–269 | KAISSYVLAQRV | |||
| 240–252 | KKVAAGFELSNQV | |||
| 202–213 | KLQNLNFSKTNL | |||
| RtxA | K. kingae | 444–453 | RHAHYLERNL | |
| 409–416 | KQAMFESV | |||
| 339–349 | KKFGYDGDSLL | |||
| 280–287 | KAISSYVL | |||
| 262–274 | KKVAAGFELSNQV |
Table 4.
CRAC motifs identified in the sequences corresponding to the pore-forming domain of various RTX toxins. The searching input algorithm used with the EMBOSS explorer FuzzPro software (https://www.bioinformatics.nl/cgi-bin/emboss/fuzzpro, (accessed on 5 March 2024), Alan Bleasby, EBI, Cambridge, UK) was [L/V]-X1–5-[Y/F]-X1–5-[K/R]. The location of the motifs in the full-length toxin and their corresponding amino acid sequence are shown. Central phenylalanine and tyrosine residues are labeled in red.
Table 4.
CRAC motifs identified in the sequences corresponding to the pore-forming domain of various RTX toxins. The searching input algorithm used with the EMBOSS explorer FuzzPro software (https://www.bioinformatics.nl/cgi-bin/emboss/fuzzpro, (accessed on 5 March 2024), Alan Bleasby, EBI, Cambridge, UK) was [L/V]-X1–5-[Y/F]-X1–5-[K/R]. The location of the motifs in the full-length toxin and their corresponding amino acid sequence are shown. Central phenylalanine and tyrosine residues are labeled in red.
| Motif | RTX Toxin | Bacterium | Residues | Sequence |
|---|---|---|---|---|
| CRAC | EhxA | EHEC E. coli | 407–414 | VADKFAAR |
| 339–345 | LLAAFHK | |||
| 322–331 | LESYSERFKK | |||
| 310–320 | LAIADKFERAK | |||
| 307–315 | LSFLAIADK | |||
| 273–281 | VSQYILAQR | |||
| LktA | M. haemolytica | 401–405 | LQYSK | |
| 344–350 | LLAEYQR | |||
| 327–336 | LESYAERFKK | |||
| 312–320 | LAFAGIADK | |||
| 278–286 | VSSYILAQR | |||
| PlLktA | M. varigena | 401–405 | LQYSK | |
| 344–350 | LLAQYQR | |||
| 327–336 | LESYAERFKK | |||
| 312–320 | LAFAGIADK | |||
| 278–286 | VSSYILAQR | |||
| 220–228 | LKNFSGFSK | |||
| PaxA | P. aerogenes | 417–421 | LEFSK | |
| 353–359 | LGYEGDK | |||
| 331–341 | LRVADNFNRSK | |||
| 328–332 | LSFLR | |||
| 294–302 | VSSYILAQR | |||
| 259–268 | VTASFTLADK | |||
| 124–133 | LDKFLQQHSK | |||
| 117–126 | LTLFAPQLDK | |||
| MmxA | M. morganii | 453–462 | LEDNFKLLSK | |
| 324–334 | LAVADKFKRAR | |||
| 321–329 | LSFLAVADK | |||
| 287–295 | VSQYILAQR | |||
| HlyA | UPEC E. coli | 459–464 | LSQYNK | |
| 352–358 | LLAAFHK | |||
| 323–334 | LSIADKFKRANK | |||
| 320–328 | LSFLSIADK | |||
| 198–210 | LNNVNSFSQQLNK | |||
| 117–127 | LLQKYQKAGNK | |||
| CyaA | B. pertussis | 732–741 | LGGPQAYFEK | |
| 721–728 | LANDYARK | |||
| 653–661 | LLAQLYRDK | |||
| 626–638 | LVQQSHYADQLDK | |||
| 518–527 | VSGFFRGSSR | |||
| 481–487 | LMTQFGR | |||
| LtxA | A. actinomycetemcomitans | 455–464 | LKLFNELREK | |
| 351–357 | LLGQFYK | |||
| 334–343 | LEEYSKRFKK | |||
| 322–332 | LGIAKQFDRAR | |||
| 319–326 | LSFLGIAK | |||
| 217–228 | VKHFGSFGDKLK | |||
| 214–226 | LGQVKHFGSFGDK | |||
| 200–209 | VDTFSKQLNK | |||
| ApxIA | A. pleuropneumoniae | 455–461 | LLSQYNK | |
| 349–355 | LLASFYR | |||
| 332–341 | LEQYSERFKK | |||
| 320–330 | LNVADKFERAK | |||
| 317–325 | LSFLNVADK | |||
| 283–291 | VSQYIIAQR | |||
| 247–257 | VVSASFILSNK | |||
| 199–208 | VDAFAEQLGK | |||
| 106–117 | LFAPQFDKLLNK | |||
| ApxIIA | A. pleuropneumoniae | 449–461 | LQDNMKFLINLNK | |
| 406–410 | LEYSK | |||
| 349–355 | LLADFHR | |||
| 342–348 | LGYDGDR | |||
| 331–341 | LIKSYSERFQK | |||
| 314–325 | VTPLSFLNVADK | |||
| 283–291 | VSSYILAQR | |||
| 195–207 | VQTVDAFAEQISK | |||
| 101–106 | LGFTDR | |||
| ApxIIIA | A. pleuropneumoniae | 417–421 | LEFSK | |
| 353–359 | LGYDGDK | |||
| 331–341 | LRVADNFNRSK | |||
| 328–332 | LAFLR | |||
| 294–302 | VSSYILAQR | |||
| 259–268 | VTASFALANK | |||
| 124–133 | LDQFLQKHSK | |||
| MbxA | M. bovis | 427–439 | LANNLKFLSELNK | |
| 326–332 | LLAEYQR | |||
| 309–318 | LDEFAKQFRK | |||
| 294–302 | LAFMNAADK | |||
| 203–210 | LQNLNFSK | |||
| RtxA | K. kingae | 348–354 | LLAEYQR | |
| 330–340 | LIDEFAKQFKK | |||
| 316–324 | LAFMNAADK | |||
| 224–232 | LQNLPNFGK | |||
| 194–206 | VQSIEAFSEQLGR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ostolaza, H.; Amuategi, J. Membrane Interaction Characteristics of the RTX Toxins and the Cholesterol-Dependence of Their Cytolytic/Cytotoxic Activity. Int. J. Mol. Sci. 2024, 25, 3131. https://doi.org/10.3390/ijms25063131
AMA Style
Ostolaza H, Amuategi J. Membrane Interaction Characteristics of the RTX Toxins and the Cholesterol-Dependence of Their Cytolytic/Cytotoxic Activity. International Journal of Molecular Sciences. 2024; 25(6):3131. https://doi.org/10.3390/ijms25063131
Chicago/Turabian StyleOstolaza, Helena, and Jone Amuategi. 2024. "Membrane Interaction Characteristics of the RTX Toxins and the Cholesterol-Dependence of Their Cytolytic/Cytotoxic Activity" International Journal of Molecular Sciences 25, no. 6: 3131. https://doi.org/10.3390/ijms25063131
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.