
PPR596 Is Required for nad2 Intron Splicing and Complex I Biogenesis in Arabidopsis

Key Laboratory of Plant Development and Environmental Adaptation Biology, Ministry of Education, School of Life Sciences, Shandong University, Qingdao 266237, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(6), 3542; https://doi.org/10.3390/ijms25063542
Submission received: 21 February 2024
/
Revised: 15 March 2024
/
Accepted: 16 March 2024
/
Published: 21 March 2024
(This article belongs to the Section Molecular Plant Sciences)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Mitochondria are essential organelles that generate energy via oxidative phosphorylation. Plant mitochondrial genome encodes some of the respiratory complex subunits, and these transcripts require accurate processing, including C-to-U RNA editing and intron splicing. Pentatricopeptide repeats (PPR) proteins are involved in various organellar RNA processing events. PPR596, a P-type PPR protein, was previously identified to function in the C-to-U editing of mitochondrial rps3 transcripts in Arabidopsis. Here, we demonstrate that PPR596 functions in the cis-splicing of nad2 intron 3 in mitochondria. Loss of the PPR596 function affects the editing at rps3eU1344SS, impairs nad2 intron 3 splicing and reduces the mitochondrial complex I’s assembly and activity, while inducing alternative oxidase (AOX) gene expression. This defect in nad2 intron splicing provides a plausible explanation for the slow growth of the ppr595 mutants. Although a few P-type PPR proteins are involved in RNA C-to-U editing, our results suggest that the primary function of PPR596 is intron splicing.
1. Introduction
Mitochondria are the major cellular organelles important for energy production and the synthesis of essential metabolites. Mitochondria evolved 1.5 billion years ago via the symbiosis of an early anaerobic eukaryote cell engulfing an α-protobacterium. During evolution, most mitochondrial genes either shifted to the nucleus or were lost [1]. Animal mitochondrial genomes are concise, with 37 genes encoding merely 13 proteins, 22 tRNAs, and 2 rRNAs [2]. Conversely, plant mitochondrial genomes, notably larger and structurally diverse, contain around 60 genes that primarily code for essential components like rRNAs, tRNAs, and subunits of respiratory complexes (CI to CV). Despite their compact size, mitochondria require numerous nuclear-encoded proteins for DNA maintenance, transcription, translation, and post-transcriptional processes like RNA editing and intron splicing [3,4].
Intron splicing is an essential step of post-transcriptional processing, and plants use post-transcriptional processes to regulate mitochondrial gene expression [5]. Many mitochondrial genes contain introns that must be removed and neighboring exons joined to code for functional proteins. Arabidopsis thaliana mitochondria have been found to harbor twenty-three introns, including four in NADH dehydrogenase subunit1 (nad1), nad2, nad5, and nad7, three in nad4, and one in each of the cytochrome c oxidase subunit2 (cox2), cytochrome c maturation subunit F C-terminus (ccmFc), ribosomal protein large subunit2 (rpl2), and ribosomal protein small subunit3 (rps3). Plant mitochondrial genomes contain cis- and trans-introns. Thus, the trans-introns in nad1, nad2, and nad5 transcripts require trans-splicing [6]. Regardless of intron configuration, introns are classified into group I and group II based on their structure and splicing mechanism. Group I introns comprise 10 conserved paired regions, P1–P10, in the core secondary structure. The splicing of group I introns is facilitated by two-step phosphoryl transfer reactions that use exogenous guanosine as a cofactor [7,8]. Only one group I intron has been discovered in the cox1 gene of multiple plant species [9]. Almost all mitochondrial introns are group II introns. Group II introns are characterized by six stem-loop structures (e.g., domains I-VI), and the splicing mechanism resembles that of nuclear pre-mRNA splicing, involving two transesterification steps and the intron being released as a lariat [10]. In bacteria, the group II intron domain IV encodes a maturase, which is crucial for facilitating the splicing of its cognate group II intron. In higher plants, only one maturase, MatR encoded by nad1 intron 4, is found in the mitochondrial genome [11]. Meanwhile, four maturases, named nMAT1 to nMAT4, are encoded by the nuclear genome in Arabidopsis. Higher plants such as angiosperms, however, have evolved numerous nucleus-encoded factors to facilitate intron splicing, including CRM (Chloroplast RNA splicing, ribosome maturation (CRM), Plant Organellar RNA Recognition (PORR) domain protein, Regulator of Chromosome Condensation (RCC), and pentatricopeptide repeat (PPR) protein [12].
PPR proteins are a prominent group of proteins in land plants, with over 450 members in Arabidopsis thaliana predominantly localized in chloroplasts or mitochondria [13,14]. Typically characterized by 2–30 tandem repeats of a 35-amino-acid motif, each PPR motif folds into a pair of antiparallel helices [15,16]. They are classified into the P and PLS subfamilies based on motif structures, with the latter further divided into PLS, E, and DYW subclasses based on the presence of E or DYW motifs in the C-terminal sequences [17]. Members of the P subfamily participate in various organellar post-transcriptional processes, including RNA stabilization and splicing, whereas PLS subfamily proteins are involved in RNA editing [16,18]. PPR proteins bind to their cognate nucleotides in a target-specific manner. Each PPR repeat of a PPR protein binds to a single RNA nucleotide in a modular pattern. The amino acids at the fifth and thirty-fifth positions determine the specificity of the PPR protein [16,19,20,21]. PPR proteins play a critical role in mitochondrial intron splicing, and mutations in the genes encoding these proteins lead to defects in splicing, impacting mitochondrial respiratory complex assembly and causing growth and developmental abnormalities [22,23,24,25,26,27]. Various PPR proteins contribute to the splicing of introns in multiple genes, underscoring their significance in organelle biogenesis [23,28].
PPR596 encodes a mitochondrion-localized P-type PPR protein. It was previously reported to be involved in RNA C-to-U editing at the rps3eU1344SS site of the rps3 transcript in Arabidopsis thaliana [29]. In this study, we identified that PPR596 functions in intron splicing, probably the primary function that contributed to the observed slow growth phenotype in its mutants. The loss of PPR596 function results in defective cis-splicing of nad2 intron 3, leading to a significant reduction in the assembly and activity of complex I, increases the expression of alternative oxidase genes, and ultimately results in the slow growth phenotype in Arabidopsis thaliana.
2. Results
2.1. PPR596 Encodes a Mitochondrion-Localized PPR Protein
AT1G80270 encodes a mitochondrion-localized P-type PPR protein consisting of 596 amino acids and was previously named PPR596 [29]. Unlike most of the PPR genes that are intronless, PPR596 contains three introns, with two introns in the coding region and one in the 5′-untranslated region (Figure 1A). This PPR protein contains eight P-type PPR repeats, according to the PPRCODE prediction software (http://yinlab.hzau.edu.cn/pprcode) (accessed on 20 August 2023) (Figure 1B).
TargetP [30], MitoFates (aist.go.jp) (accessed on 21 April 2023), and ExPASy portal (https://www.expasy.org) (accessed on 21 April 2023) collectively indicate the presence of 61 amino acid mitochondrial targeting sequences at the N-terminal region of PPR596 (Figure S1). Experimental evidence confirmed the mitochondria localization of PPR596 [29]. Additionally, the in silico 3D structure prediction results obtained from the AlphaFold server [31] suggest that PPR596 has a typical PPR helical fold (Figure S2), including an inner basic core that serves as the RNA-binding surface. This finding is consistent with previous studies on plant PPR10 proteins [32].
2.2. Characterization of the ppr596 Mutants
The T-DNA insertion line (SAIL_367_A06) was obtained from the Arabidopsis Biological Resource Center (ABRC; Columbus, OH, USA). Homozygous plants were identified by PCR analysis (Figure 1D). The T-DNA insertion was confirmed by sequencing and was found to be located 207 nt upstream of the stop codon of PPR596 (Figure 1A). The expression of wild-type PPR596 was not detected in homozygous mutants, whereas the transcript levels of Actin2 (At3g18780) were not affected (Figure 1C).
The ppr596 mutants displayed a noticeably slower growth phenotype than the wild-type plants. The ppr596 seed germination was significantly delayed, and after two weeks, the mutant plants were visibly smaller than their wild-type counterparts (Figure 2C). Based on soil-based phenotypic analysis, plant growth was retarded throughout the germination stage. The ppr596 mutant plants had a delay of approximately 30 days at the onset of flowering compared with the wild-type plants (Figure 2B,D). Under the light microscope, mature seeds of the mutant plants were smaller and wrinkled, unlike mature wild-type seeds (Figure 2A). Mutant plants expressing the open reading frame of AT1G80270 under the control of the 35S promoter and flag-tag at the C terminal rescued the slow growth phenotype and seed development. They grew similarly to wild-type plants, indicating that the lack of PPR596 is the cause of the slow growth phenotype.
2.3. Expression of nad2 Is Reduced in ppr596
PPR proteins are RNA-binding proteins involved in RNA metabolism including RNA maturation, splicing, and C-to-U RNA editing in mitochondria and chloroplasts. PPR596 is a P-type PPR protein that has previously been reported to partially decrease the editing of mitochondrial ribosomal protein S3 (RPS3) transcripts [29]. PPR596, POCO1 [33] and PPME [34], have been frequently cited as exceptional P-type PPR proteins for their role in RNA-editing [16,35,36,37]. However, most P-type PPR proteins are primarily associated with RNA stabilization and splicing in organelles. To further investigate the possible functions of PPR596, all mature transcripts of mitochondrial protein-coding genes were examined by semi-quantitative RT-PCR in the ppr596 mutant, WT, and complementation lines using the primers listed in Table S1. Notably, we observed a significant reduction in the abundance of mature nad2 transcripts in the ppr596 mutant compared with the WT and complementation lines (Figure 3). Additionally, the expression levels of various mitochondrial transcripts, including nad1, nad4, nad6, nad7, atp4, ccmFn2, cox1, cox2, cox3, Rps4, rps12, and MAT-R, were higher in the ppr596 mutant than in WT and complemented plants.
2.4. PPR596 Is Required for Splicing of the nad2 Intron 3
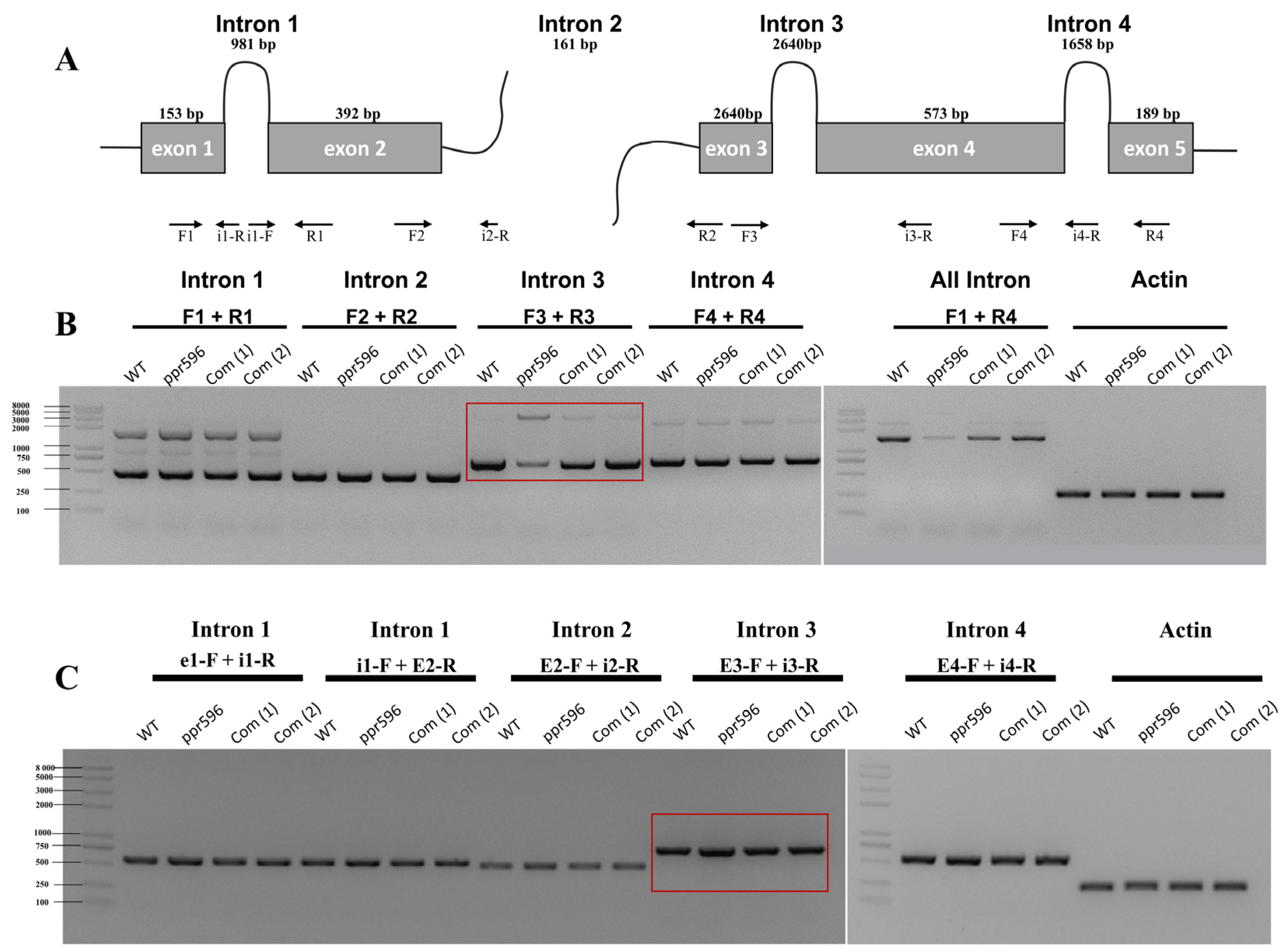
RT-PCR analysis of transcripts from mitochondrial protein-coding genes revealed a significant reduction in the accumulation of mature nad2 transcripts in the ppr596 mutant (Figure 3). The nad2 transcript contains three cis introns and one trans intron (Figure 4A), and the maturation of nad2 mRNA requires the proper splicing of these introns. To address whether the loss of PPR596 affects the splicing of these introns, we performed RT-PCR analysis in the ppr596, WT, and complemented plants using the primers listed in Table S1. The primer positions are shown in Figure 4A. RT-PCR analysis indicated a reduction in the splicing efficiency of nad2 intron 3 in ppr596 compared to the WT and complementation lines (Figure 4B). Defective splicing of nad2 intron 3 was confirmed by the accumulation of unspliced intron 3 in the mutant plants (Figure 4C).
To verify this result by an independent approach, we conducted quantitative RT-PCR using the splicing efficiency of the nad4 and nad5 introns as controls. qRT-PCR results showed a (5.6-fold) decrease in the splicing efficiency of nad2 intron 3 in mutant plants compared with the wild-type (Figure 5).
2.5. Biogenesis of Respiratory Complex I Was Affected in ppr596
nad2 encodes a subunit of the mitochondrial complex I (CI) in the electron transfer chain. Nad2 is the core CI subunit located in the matrix arm [38]. To investigate the effect of defective splicing on the assembly of complex I, we analyzed the mitochondrial complex profile by blue native polyacrylamide gel electrophoresis (BN-PAGE), in-gel activity assay, and immunoblotting. Mitochondria were isolated from ppr596, WT, and complementation line seedlings grown in the dark as described [39]. Mitochondrial membrane complexes were solubilized with n-dodecyl-β-D-maltoside (β-DM), separated by BN-PAGE, and subjected to in-gel complex activity assay [40] and immunoblotting assays. Coomassie Brilliant Blue (CBB) staining of the gels revealed a significant reduction in CI in the ppr596 mutant compared to WT and complemented plants (Figure 6A). The in-gel activity of CI indicated that the CI activity was severely reduced in ppr596 and partially restored in the complementation line (Figure 6B). The reduction in CI assembly was further confirmed by immunoblotting using antibodies against the CI carbonic anhydrase-like subunit 2 (CA2), one of the first subunits to be incorporated in CI (Figure 6C) [41]. These results strongly support the crucial role of PPR596 in the assembly of CI.
To address the effect of reduced partial C-to-U editing of rps3 and defective nad2 splicing on the abundance of mitochondrial proteins, immunoblotting was performed using antibodies against L16, COX1, NAD9, and CA2. The abundance of mitochondria-encoded proteins L16 and COX1 was normal in PPR596 mutants. However, in agreement with the dramatic reduction in CI, the accumulation of CA2 and NAD9 was severely decreased in the ppr596 mutant (Figure 6D).
It has been reported that the dysfunction of mitochondrial CI activates the alternative respiratory pathway as an adaptive response to mitochondrial stress [42,43,44]. To further assess the induction of alternative respiratory pathways, we measured the expression levels of various alternative oxidase (AOX) genes and rotenone-insensitive NAD(P)H dehydrogenase (NDs) in mutant plants. qRT-PCR analysis indicated that the relative accumulation of these genes transcripts was higher in mutant plants than in WT plants (Figure 7).
2.6. Expression Pattern and Conservation of PPR596 in Angiosperms
PPR596 expression was evaluated by using the publicly available database The Arabidopsis Information Resource (TAIR), which indicates that the PPR596 gene (AT1G80270) is widely expressed throughout various developmental stages of Arabidopsis thaliana. Its expression is notably prominent during seed germination in apical root tissues, young developing leaves, certain flower stages (particularly in carpels), and the shoot apex (Figure S3). To gain insights into its evolutionary relationship, homologous proteins of PPR596 were identified from the UniProt database using the Basic Local Alignment Search Tool (BLAST), and a phylogenetic tree was constructed. The analysis revealed that PPR596 is closely related to putative orthologs in dicots, especially with Brassica napus and Nicotiana tabacum. It formed a distinct cluster of monocots (Triticum aestivum, Hordeum vulgare, Oryza sativa, and Zea mays), suggesting that PPR596 homologs in these species may have evolved from a common ancestor (Supplemental Figure S4A). Detailed sequence alignment indicated that PPR596 is highly conserved among angiosperms (Figure S4B).
3. Discussion
Mitochondrial biogenesis relies on the correct expression of the mitochondrial genome. Before translation, mitochondrial transcripts undergo RNA maturation, including RNA C-to-U editing, cis- and trans-splicing of introns, and maturation of 3′- and 5′-ends. Most introns in mitochondrial transcripts are group II and require nucleus-encoded splicing factors such as maturases, chloroplast RNA splicing and ribosome maturation (CRM) domain-containing proteins, and PPR proteins [12]. PPR proteins are RNA-binding proteins involved in organellar RNA maturation events such as RNA stabilization and splicing. The P-type of the PPR protein family has predominantly been reported for its role in intron splicing, except for PPR596, PPME, and POCO1, which were reported to function in the C-to-U RNA editing [29,33,34]. PPR596 is a previously identified P-type PPR protein linked to reduced C-to-U-editing of mitochondrial rps3 transcripts [29]. However, whether PPR596 is directly or indirectly involved in C-to-U editing is not addressed. We suspect the slight reduction in the C-to-U editing at the rps3eU1344SS site of the rps3 transcript may not be the primary cause for the severely slow growth phenotype in ppr596.
This study demonstrates that PPR596 is required for the splicing of nad2 intron 3, which is essential to the expression of Nad2 protein, a core subunit of mitochondrial CI. qRT-PCR analysis showed that the splicing efficiency of nad2 intron 3 was severely decreased in the ppr596 mutant, while other nad2 transcript introns also displayed slightly reduced splicing efficiency (Figure 5). Conventional RT-PCR analysis further confirmed the severe decrease in the splicing efficiency of nad2 intron 3 in the ppr596 mutant (Figure 4). Many P-type PPR proteins, such as MISF26, MISF68, MISF74, ABO5, MISF2, and OTP43, play crucial roles in mitochondrial intron splicing [22,23,25,27]. Our findings suggest that the lack of PPR596 primarily affects the splicing of nad2 intron 3, with the observed reduction in splicing efficiency of other nad2 transcript introns possibly being a secondary consequence of impaired nad2 intron 3 splicing. Previous research has demonstrated interdependence between the splicing efficiency of different introns [45], supported by the observations in rug3, where reduced splicing efficiency of nad2 intron 3 affects the splicing effect of intron 2 [46]. This evidence strongly supports the requirement of PPR596 for the efficient splicing of nad2 intron 3 in Arabidopsis thaliana.
The plant mitochondrial respiratory chain comprises five multi-subunit protein complexes (CI-CV). CI is a large L-shaped complex composed of two arms: a membrane arm embedded in the mitochondrial inner membrane, and a matrix arm protruding into the matrix [47]. In angiosperms, CI comprises approximately 50 subunits encoded by nuclear and mitochondrial genomes [48]. Nad2, a core subunit of CI, is incorporated into the membrane arm during the early CI assembly. A reduction in Nad2 leads to decreased CI assembly. Mutants with defective nad2 splicing exhibit low CI abundance and reduced complex I activity [23,25,46,49,50]. Similarly, our BN-PAGE analysis and in-gel activity assays of CI revealed a severe reduction in CI abundance in the ppr596 mutant compared to WT and complementation lines. Immunoblot analysis with anti-CA2 further confirmed the reduced CI assembly in ppr596. Complex I serves as the entry point for the electron transfer chain. The dysfunction of mitochondrial CI triggers the activation of alternative respiratory pathways in Arabidopsis thaliana [25,27,51]. Correspondingly, the expression of transcripts corresponding to various alternative oxidases (AOX) and rotenone-insensitive NAD(P)H dehydrogenase (NDs) was upregulated in ppr596, suggesting the induction of an alternative respiratory pathway (Figure 7). These findings strongly suggest the essential role of PPR596 in the nad2 intron 3 splicing and biogenesis of respiratory complex I.
Defective splicing of nad2 intron 3 and reduced complex I assembly are the primary cause of the observed slow growth phenotype in ppr596. Despite reduced partial C-to-U editing in the rps3 transcript, ppr596 can produce normal proteins, as in WT plants [29]. The normal accumulation of mitochondrion-encoded proteins, COX1 and L16, in the ppr596 mutant mitochondria suggests that the partially reduced C-to-U editing in rps3 transcripts does not strongly affect mitochondrial translation. Arabidopsis mutants defective in nad2 splicing and complex I assembly showed similar slow growth phenotypes [23,46,49]. This evidence suggests that the primary function of PPR596 is in the efficient splicing of nad2 intron 3, and a lack of function leads to reduced complex I assembly, resulting in the slow growth phenotype.
PPR proteins function as RNA-binding proteins by recognizing specific sequences in target RNA. The PPR code, defined by combining the fifth and last amino acids in each PPR motif, facilitates nucleotide recognition [52,53]. However, the code prediction of PPR 596 failed using the PPRCODE web server (Figure S1B) [19]. The difficulty in determining the sequence specificity of PPR proteins may be attributed to their other distinctive features [54]. The precise mechanism through which PPR596 is involved in C-to-U editing and nad2 intron 3 splicing remains unknown. To date, two PPR proteins, ABO5 [22] and MISF26 [23], along with several other splicing factors like ABO6, CFM9, mCSF1, RUG3, mTERF15 and Organelle Zinc (OZ2) [46,50,55,56,57,58], have been implicated in the splicing of nad2 intron 3. Exploring the interaction between PPR596 and these splicing factors could provide valuable insights into the complexities of the plant mitochondrial spliceosome.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Arabidopsis ppr596 mutant (SAIL_367_A06) seeds were obtained from the Arabidopsis Biological Resource Center (ABRC, Columbus, OH, USA) in the Col-0 background. Before sowing, the seeds were surface sterilized with 75% ethanol with 0.05% Triton X-100 solution for 10 min, followed by 3 min sterilization with 3% sodium hypochlorite (NaClO) solution. Immediately thereafter, the seeds were washed five times with sterile water. The seeds were planted on Murashige and Skoog (MS) medium (pH 5.7) with 3% sucrose and 0.8% agar, incubated at 4 °C for three days, and then grown under long-day conditions (16 h light/8 h dark photoperiod) at 22 °C in a growth chamber. After germination on MS medium for 2 weeks, the plants were transferred to soil.
4.2. Plants Genotyping
4.3. Plant Complementation
The coding sequence of PPR596 was amplified by PCR from the cDNA of wild-type Arabidopsis using the primers listed in Table S1. The PCR product was successfully cloned into pENTR/D-TOPO (Invitrogen, Carlsbad, CA, USA), confirmed by Sanger sequencing, and transferred into the binary vector pGWB11 through Gateway site-specific recombination. Subsequently, the construct was introduced into ppr596 homozygous mutant plants via Agrobacterium tumefaciens-mediated transformation using the floral dip method [60].
4.4. RNA Extraction RT-PCR and qRT-PCR
Total RNA was extracted from two-week-old seedlings of the WT, complemented lines, and one-month-old seedlings of the mutant plants using RNeasy Plant Mini Kit (Vazyme Biotech, Nanjing, China). RNA was treated with RNase-free DNase I to remove potential DNA contamination (New England Biolabs, Rowley, MA, USA). cDNA was synthesized using the Transcript FirstStrand cDNA Synthesis SuperMix (TransGen Biotech, Beijing, China). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using a LightCycler 96 (Roche, Basel, Switzerland) with three biological replicates. The Arabidopsis actin gene AtActin was utilized as a reference. For the functional analysis of PPR596, RT-PCR and qRT-PCR were performed using the primers listed in Table S1.
4.5. Mitochondria Isolation
Crude mitochondria were isolated from two-week-old seedlings of WT, complemented lines, and one-month-old seedlings of mutant plants grown in the dark on Murashige and Skoog (MS) medium with 3% sucrose and 0.8% agar, using a previously described protocol [39]. Fresh seedlings of the samples were homogenized with the help of a pestle and mortar on ice in extraction buffer containing 0.3 M sucrose, 5 mM tetrasodium pyrophosphate, 10 mM KH2PO4, pH 7.5, 2 mM EDTA, 1% (w/v) polyvinylpyrrolidone 40, 1% (w/v) BSA, 5 mM cysteine, and 20 mM ascorbic acid.
The homogenate was centrifuged at 3000× g for 5 min at 4 °C, and the resulting supernatant was centrifuged at 18,000× g for 30 min at the same temperature. The pellets were resuspended in a wash buffer consisting of 0.3 M sucrose, 1 mM EGTA, 10 mM MOPS-KOH, and pH 7.2. Protein concentration was determined using the Bradford method [61].
4.6. SDS-PAGE
4.7. BN-PAGE
Mitochondrial proteins were solubilized using β-DM and separated by blue native polyacrylamide gel electrophoresis (BN-PAGE) [64]. BN-PAGE was performed at 4 °C in a vertical apparatus using blue cathode buffer (0.02% Coomassie Blue G-250) as the running buffer. Separation gels were created using linear gradients of 3–12% or 4–16% polyacrylamide (Invitrogen, Carlsbad, CA, USA).
4.8. In-Gel Staining
The in-gel complex activity assay was performed as previously described [40]. The complex I assay used 100 mM Tris-HCl, pH 7.5, 768 mM glycine, 0.1 mM NADH, and 0.04% nitrotetrazolium blue (NTB) (w/v).
4.9. Immunoblotting
The assembly of complex I was detected by immunoblotting. BN-PAGE gels were first treated with a denaturation buffer (1% SDS, 50 mM Tris-HCl, and 0.05% β-mercaptoethanol) for 30 min. For the immunoblotting analysis, mitochondrial proteins were transferred onto PVDF membranes (0.45 mm; Millipore, Burlington, MA, USA). The PVDF membranes were incubated with CA2 primary antibody against CA2, an early assembly factor of complex I as described [65]. For the immunodetection of mitochondrial-encoded proteins, proteins from the SDS page were transferred onto PVDF membranes (0.45 mm; Millipore, Burlington, MA, USA). The PVDF membranes were incubated with the primary antibodies L16, COX1, and CA2. Signal detection was carried out by ECL reagents (Thermo Fisher Scientific, Waltham, MA, USA) after incubation with the horseradish peroxidase (HRP)-conjugated secondary antibody.
4.10. Phylogenetic Analysis
Homologous protein sequences of PPR596 were retrieved from the UniProt database using the Basic Local Alignment Search Tool (BLAST). Alignment of these sequences from various plant species was conducted using Geneious 9.2.2 software. A phylogenetic tree was constructed using the Maximum Likelihood method and JTT matrix-based model in MEGA11. The analysis was performed using 1000 replicates [66].
Supplementary Materials
The supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25063542/s1.
Author Contributions
A.S. and B.-C.T. designed the research. A.S. performed most of the experiments. B.C., Y.W. and S.-K.C. participated in vector construction. A.S. and B.-C.T. analyzed the data and wrote the article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (project Nos. 32230075 to B.-C.T. and 32101640 to Y.W.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martin, W.; Herrmann, R.G. Gene transfer from organelles to the nucleus: How much, what happens, and why? Plant Physiol. 1998, 118, 9–17. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef]
- Hammani, K.; Giege, P. RNA metabolism in plant mitochondria. Trends Plant Sci. 2014, 19, 380–389. [Google Scholar] [CrossRef] [PubMed]
- de Longevialle, A.F.; Small, I.D.; Lurin, C. Nuclearly encoded splicing factors implicated in RNA splicing in higher plant organelles. Mol. Plant 2010, 3, 691–705. [Google Scholar] [CrossRef]
- Binder, S.; Brennicke, A. Gene expression in plant mitochondria: Transcriptional and post–transcriptional control. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 181–189. [Google Scholar] [CrossRef]
- Malek, O.; Knoop, V. Trans-splicing group II introns in plant mitochondria: The complete set of cis-arranged homologs in ferns, fern allies, and a hornwort. RNA 1998, 4, 1599–1609. [Google Scholar] [CrossRef]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Cech, T.R. Self-splicing of group I introns. Annu. Rev. Biochem. 1990, 59, 543–568. [Google Scholar] [CrossRef]
- Cho, Y.; Adams, K.; Qiu, Y.; Kuhlman, P.; Vaughn, J.; Palmer, J. A highly invasive group I intron in the mitochondrial cox1 gene. In Plant Mitochondria: Gene Function; Backhuys Publisher: Leiden, The Netherlands, 1998; pp. 19–23. [Google Scholar]
- Lambowitz, A.M.; Zimmerly, S. Mobile group II introns. Annu. Rev. Genet. 2004, 38, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Wahleithner, J.A.; MacFarlane, J.L.; Wolstenholme, D.R. A sequence encoding a maturase-related protein in a group II intron of a plant mitochondrial nad1 gene. Proc. Natl. Acad. Sci. USA 1990, 87, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.G.; Colas des Francs-Small, C.; Ostersetzer-Biran, O. Group II intron splicing factors in plant mitochondria. Front. Plant Sci. 2014, 5, 35. [Google Scholar] [CrossRef]
- Schmitz-Linneweber, C.; Small, I. Pentatricopeptide repeat proteins: A socket set for organelle gene expression. Trends Plant Sci. 2008, 13, 663–670. [Google Scholar] [CrossRef]
- Colcombet, J.; Lopez-Obando, M.; Heurtevin, L.; Bernard, C.; Martin, K.; Berthome, R.; Lurin, C. Systematic study of subcellular localization of Arabidopsis PPR proteins confirms a massive targeting to organelles. RNA Biol. 2013, 10, 1557–1575. [Google Scholar] [CrossRef]
- Yin, P.; Li, Q.; Yan, C.; Liu, Y.; Liu, J.; Yu, F.; Wang, Z.; Long, J.; He, J.; Wang, H.W.; et al. Structural basis for the modular recognition of single-stranded RNA by PPR proteins. Nature 2013, 504, 168–171. [Google Scholar] [CrossRef]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Lurin, C.; Andreés, C.; Aubourg, S.; Bellaoui, M.; Bitton, F.; Bruyère, C.; Caboche, M.; Debast, C.; Gualberto, J.; Hoffmann, B. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 2004, 16, 2089–2103. [Google Scholar] [CrossRef]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Hartel, B.; Brennicke, A. RNA editing in plants and its evolution. Annu. Rev. Genet. 2013, 47, 335–352. [Google Scholar] [CrossRef]
- Yan, J.; Yao, Y.; Hong, S.; Yang, Y.; Shen, C.; Zhang, Q.; Zhang, D.; Zou, T.; Yin, P. Delineation of pentatricopeptide repeat codes for target RNA prediction. Nucleic Acids Res. 2019, 47, 3728–3738. [Google Scholar] [CrossRef]
- Coquille, S.; Filipovska, A.; Chia, T.; Rajappa, L.; Lingford, J.P.; Razif, M.F.; Thore, S.; Rackham, O. An artificial PPR scaffold for programmable RNA recognition. Nat. Commun. 2014, 5, 5729. [Google Scholar] [CrossRef]
- Shen, C.; Wang, X.; Liu, Y.; Li, Q.; Yang, Z.; Yan, N.; Zou, T.; Yin, P. Specific RNA recognition by designer pentatricopeptide repeat protein. Mol. Plant 2015, 8, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, J.; Chen, Z.; Ren, X.; Hong, X.; Gong, Z. ABA overly-sensitive 5 (ABO5), encoding a pentatricopeptide repeat protein required for cis-splicing of mitochondrial nad2 intron 3, is involved in the abscisic acid response in Arabidopsis. Plant J. 2010, 63, 749–765. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Aube, F.; Quadrado, M.; Dargel-Graffin, C.; Mireau, H. Three new pentatricopeptide repeat proteins facilitate the splicing of mitochondrial transcripts and complex I biogenesis in Arabidopsis. J. Exp. Bot. 2018, 69, 5131–5140. [Google Scholar] [CrossRef]
- Weißenberger, S.; Soll, J.; Carrie, C. The PPR protein SLOW GROWTH 4 is involved in editing of nad4 and affects the splicing of nad2 intron 1. Plant Mol. Biol. 2017, 93, 355–368. [Google Scholar] [CrossRef]
- Nguyen, T.-T.; Best, C.; Shevtsov, S.; Zmudjak, M.; Quadrado, M.; Mizrahi, R.; Zer, H.; Mireau, H.; Ostersetzer-Biran, O. MISF2 Encodes an essential mitochondrial splicing cofactor required for nad2 mRNA processing and embryo development in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 2670. [Google Scholar] [CrossRef] [PubMed]
- Koprivova, A.; des Francs-Small, C.C.; Calder, G.; Mugford, S.T.; Tanz, S.; Lee, B.R.; Zechmann, B.; Small, I.; Kopriva, S. Identification of a pentatricopeptide repeat protein implicated in splicing of intron 1 of mitochondrial nad7 transcripts. J. Biol. Chem. 2010, 285, 32192–32199. [Google Scholar] [CrossRef] [PubMed]
- De Longevialle, A.F.; Meyer, E.H.; Andrés, C.; Taylor, N.L.; Lurin, C.; Millar, A.H.; Small, I.D. The pentatricopeptide repeat gene OTP43 is required for trans-splicing of the mitochondrial nad1 intron 1 in Arabidopsis thaliana. Plant Cell 2007, 19, 3256–3265. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, J.; Zhong, S.; Gu, H.; He, S.; Qu, L.J. Novel DYW-type pentatricopeptide repeat (PPR) protein BLX controls mitochondrial RNA editing and splicing essential for early seed development of Arabidopsis. J. Genet. Genom. 2018, 45, 155–168. [Google Scholar] [CrossRef]
- Doniwa, Y.; Ueda, M.; Ueta, M.; Wada, A.; Kadowaki, K.; Tsutsumi, N. The involvement of a PPR protein of the P subfamily in partial RNA editing of an Arabidopsis mitochondrial transcript. Gene 2010, 454, 39–46. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; Von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Gully, B.S.; Cowieson, N.; Stanley, W.A.; Shearston, K.; Small, I.D.; Barkan, A.; Bond, C.S. The solution structure of the pentatricopeptide repeat protein PPR10 upon binding atpH RNA. Nucleic Acids Res. 2015, 43, 1918–1926. [Google Scholar] [CrossRef]
- Emami, H.; Kempken, F. PRECOCIOUS 1 (POCO 1), a mitochondrial pentatricopeptide repeat protein affects flowering time in Arabidopsis thaliana. Plant J. 2019, 100, 265–278. [Google Scholar] [CrossRef]
- Leu, K.-C.; Hsieh, M.-H.; Wang, H.-J.; Hsieh, H.-L.; Jauh, G.-Y. Distinct role of Arabidopsis mitochondrial P-type pentatricopeptide repeat protein-modulating editing protein, PPME, in nad1 RNA editing. RNA Biol. 2016, 13, 593–604. [Google Scholar] [CrossRef]
- Fujii, S.; Small, I. The evolution of RNA editing and pentatricopeptide repeat genes. New Phytol. 2011, 191, 37–47. [Google Scholar] [CrossRef]
- Huang, C.; Liu, D.; Li, Z.-A.; Molloy, D.P.; Luo, Z.-F.; Su, Y.; Li, H.-O.; Liu, Q.; Wang, R.-Z.; Xiao, L.-T. The PPR protein RARE1-mediated editing of chloroplast accD transcripts is required for fatty acid biosynthesis and heat tolerance in Arabidopsis. Plant Commun. 2023, 4, 100461. [Google Scholar] [CrossRef]
- Guo, J.; Zhao, Y.; Chu, Y.; Li, Y.; Song, Y.; Pan, Q.; Qiu, Z.; Yu, B.; Zhu, Y. Screening Candidate Genes at the Co Locus Conferring to the Columnar Growth Habit in Apple (Malus× Domestica Borkh.). Genes 2023, 14, 964. [Google Scholar] [CrossRef]
- Ghifari, A.S.; Saha, S.; Murcha, M.W. The biogenesis and regulation of the plant oxidative phosphorylation system. Plant Physiol. 2023, 192, 728. [Google Scholar] [CrossRef]
- Li, X.J.; Zhang, Y.F.; Hou, M.; Sun, F.; Shen, Y.; Xiu, Z.H.; Wang, X.; Chen, Z.L.; Sun, S.S.; Small, I.; et al. Small kernel 1 encodes a pentatricopeptide repeat protein required for mitochondrial nad7 transcript editing and seed development in maize (Zea mays) and rice (Oryza sativa). Plant J. 2014, 79, 797–809. [Google Scholar] [CrossRef]
- Wittig, I.; Karas, M.; Schagger, H. High resolution clear native electrophoresis for in-gel functional assays and fluorescence studies of membrane protein complexes. Mol. Cell. Proteom. 2007, 6, 1215–1225. [Google Scholar] [CrossRef]
- Meyer, E.H.; Solheim, C.; Tanz, S.K.; Bonnard, G.; Millar, A.H. Insights into the composition and assembly of the membrane arm of plant complex I through analysis of subcomplexes in Arabidopsis mutant lines. J. Biol. Chem. 2011, 286, 26081–26092. [Google Scholar] [CrossRef]
- Ostersetzer-Biran, O. Respiratory complex I and embryo development. J. Exp. Bot. 2016, 67, 1205–1207. [Google Scholar] [CrossRef] [PubMed]
- Kühn, K.; Obata, T.; Feher, K.; Bock, R.; Fernie, A.R.; Meyer, E.H. Complete mitochondrial complex I deficiency induces an up-regulation of respiratory fluxes that is abolished by traces of functional complex I. Plant Physiol. 2015, 168, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Fromm, S.; Göing, J.; Lorenz, C.; Peterhänsel, C.; Braun, H.-P. Depletion of the “gamma-type carbonic anhydrase-like” subunits of complex I affects central mitochondrial metabolism in Arabidopsis thaliana. Biochim. Biophys. Acta-Bioenerg. 2016, 1857, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Elina, H.; Brown, G.G. Extensive mis-splicing of a bi-partite plant mitochondrial group II intron. Nucleic Acids Res. 2010, 38, 996–1008. [Google Scholar] [CrossRef] [PubMed]
- Kühn, K.; Carrie, C.; Giraud, E.; Wang, Y.; Meyer, E.H.; Narsai, R.; des Francs-Small, C.C.; Zhang, B.; Murcha, M.W.; Whelan, J. The RCC1 family protein RUG3 is required for splicing of nad2 and complex I biogenesis in mitochondria of Arabidopsis thaliana. Plant J. 2011, 67, 1067–1080. [Google Scholar] [CrossRef]
- Meyer, E.H.; Welchen, E.; Carrie, C. Assembly of the complexes of the oxidative phosphorylation system in land plant mitochondria. Annu. Rev. Plant Biol. 2019, 70, 23–50. [Google Scholar] [CrossRef]
- Lee, C.P.; Taylor, N.L.; Millar, A.H. Recent advances in the composition and heterogeneity of the Arabidopsis mitochondrial proteome. Front. Plant Sci. 2013, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, F.; Li, N.; Shi, D.Q.; Yang, W.C. Pentatricopeptide repeat protein MID1 modulates nad2 intron 1 splicing and Arabidopsis development. Sci. Rep. 2020, 10, 2008. [Google Scholar] [CrossRef]
- Hsu, Y.-W.; Wang, H.-J.; Hsieh, M.-H.; Hsieh, H.-L.; Jauh, G.-Y. Arabidopsis mTERF15 is required for mitochondrial nad2 intron 3 splicing and functional complex I activity. PLoS ONE 2014, 9, e112360. [Google Scholar] [CrossRef]
- Best, C.; Zmudjak, M.; Ostersetzer-Biran, O. The PPR-related splicing cofactor MSP1/EMB1025 protein, encoded by At4g20090, encode an essential protein that is required for the splicing of nad1 intron 1 and for the biogenesis of complex I in Arabidopsis mitochondria. bioRxiv 2019, 615856. [Google Scholar] [CrossRef]
- Barkan, A.; Rojas, M.; Fujii, S.; Yap, A.; Chong, Y.S.; Bond, C.S.; Small, I. A combinatorial amino acid code for RNA recognition by pentatricopeptide repeat proteins. PLoS Genet. 2012, 8, e1002910. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Gutmann, B.; Zhong, X.; Ye, Y.; Fisher, M.F.; Bai, F.; Castleden, I.; Song, Y.; Song, B.; Huang, J. Redefining the structural motifs that determine RNA binding and RNA editing by pentatricopeptide repeat proteins in land plants. Plant J. 2016, 85, 532–547. [Google Scholar] [CrossRef]
- Rojas, M.; Ruwe, H.; Miranda, R.G.; Zoschke, R.; Hase, N.; Schmitz-Linneweber, C.; Barkan, A. Unexpected functional versatility of the pentatricopeptide repeat proteins PGR3, PPR5 and PPR10. Nucleic Acids Res. 2018, 46, 10448–10459. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Duan, Y.; Hua, D.; Fan, G.; Wang, L.; Liu, Y.; Chen, Z.; Han, L.; Qu, L.-J.; Gong, Z. DEXH box RNA helicase–mediated mitochondrial reactive oxygen species production in Arabidopsis mediates crosstalk between abscisic acid and auxin signaling. Plant Cell 2012, 24, 1815–1833. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Park, S.J.; Park, Y.-I.; Kang, H. CFM9, a mitochondrial CRM protein, is crucial for mitochondrial intron splicing, mitochondria function and Arabidopsis growth and stress responses. Plant Cell Physiol. 2019, 60, 2538–2548. [Google Scholar] [CrossRef] [PubMed]
- Zmudjak, M.; Colas des Francs-Small, C.; Keren, I.; Shaya, F.; Belausov, E.; Small, I.; Ostersetzer-Biran, O. m CSF 1, a nucleus-encoded CRM protein required for the processing of many mitochondrial introns, is involved in the biogenesis of respiratory complexes I and IV in A rabidopsis. New Phytol. 2013, 199, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Bentolila, S.; Gipson, A.B.; Kehl, A.J.; Hamm, L.N.; Hayes, M.L.; Mulligan, R.M.; Hanson, M.R. A RanBP2-type zinc finger protein functions in intron splicing in Arabidopsis mitochondria and is involved in the biogenesis of respiratory complex I. Nucleic Acids Res. 2021, 49, 3490–3506. [Google Scholar] [CrossRef]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Brunelle, J.L.; Green, R. One-dimensional SDS-polyacrylamide gel electrophoresis (1D SDS-PAGE). In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 541, pp. 151–159. [Google Scholar]
- Neuhoff, V.; Arold, N.; Taube, D.; Ehrhardt, W. Improved staining of proteins in polyacrylamide gels including isoelectric focusing gels with clear background at nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Electrophoresis 1988, 9, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Wang, X.; Bonnard, G.; Shen, Y.; Xiu, Z.; Li, X.; Gao, D.; Zhang, Z.; Tan, B.C. Empty pericarp7 encodes a mitochondrial E–subgroup pentatricopeptide repeat protein that is required for ccmFN editing, mitochondrial function and seed development in maize. Plant J. 2015, 84, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.; Sun, F.; Shen, Y.; Zhang, X.; Jiang, R.; Bonnard, G.; Zhang, J.; Tan, B.C. EMPTY PERICARP 16 is required for mitochondrial nad2 intron 4 cis-splicing, complex I assembly and seed development in maize. Plant J. 2016, 85, 507–519. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Gene topology, characterization, and complementation of the homozygous SAIL_367_A06 (ppr596) mutant. (A) Gene structure showing introns and exons. Blue indicates the coding region (exon), white indicates the untranslated regions, and oblique lines in the structure represent introns of the gene. The triangle indicates the T-DNA insertion site. The T-DNA insertion site in the coding region of SAIL_367_A06 (ppr596) is located at 1584bp from the start codon and 207bp upstream of the stop codon. Arrows indicate the positions of the primers used for genotyping. (B) Schematic representation of the PPR596 protein, showing mitochondrial targeting peptide (MTP) in a gray box, and blue boxes represent PPR domains in the protein. (C) Expression analysis of PPR596 in WT, ppr596 mutant, and complementation lines by RT-PCR. Actin expression was used as a loading control. (D) Genotyping of the plants for identifying homozygous ppr596 mutant plants using primers indicated in the gene structure.
Figure 1.
Gene topology, characterization, and complementation of the homozygous SAIL_367_A06 (ppr596) mutant. (A) Gene structure showing introns and exons. Blue indicates the coding region (exon), white indicates the untranslated regions, and oblique lines in the structure represent introns of the gene. The triangle indicates the T-DNA insertion site. The T-DNA insertion site in the coding region of SAIL_367_A06 (ppr596) is located at 1584bp from the start codon and 207bp upstream of the stop codon. Arrows indicate the positions of the primers used for genotyping. (B) Schematic representation of the PPR596 protein, showing mitochondrial targeting peptide (MTP) in a gray box, and blue boxes represent PPR domains in the protein. (C) Expression analysis of PPR596 in WT, ppr596 mutant, and complementation lines by RT-PCR. Actin expression was used as a loading control. (D) Genotyping of the plants for identifying homozygous ppr596 mutant plants using primers indicated in the gene structure.

Figure 2.
Phenotypic analysis of ppr596 mutant plants. (A) Morphology of WT, ppr596 homozygous mutant, and complementation line seeds under a light microscope. (B) Four-week-old seedlings of the WT, ppr596 homozygous mutant, and complementation lines (ppr596/ppr596: PPR596-FLAG) in pots. The four-week-old homozygous ppr596 mutant (ppr596/ppr596) was very small compared to WT plants. (C) Two-week-old seedlings grown on MS medium. (D) 45-day-old plants of WT, ppr596 homozygous mutant, and complementation lines (ppr596/ppr596: PPR596-FLAG) in pots. The homozygous ppr596 mutant exhibited slower growth and delayed maturation compared to both WT and complementation plants (first from the right, 75 days old). The bar in (B–D) indicates 2 cm.
Figure 2.
Phenotypic analysis of ppr596 mutant plants. (A) Morphology of WT, ppr596 homozygous mutant, and complementation line seeds under a light microscope. (B) Four-week-old seedlings of the WT, ppr596 homozygous mutant, and complementation lines (ppr596/ppr596: PPR596-FLAG) in pots. The four-week-old homozygous ppr596 mutant (ppr596/ppr596) was very small compared to WT plants. (C) Two-week-old seedlings grown on MS medium. (D) 45-day-old plants of WT, ppr596 homozygous mutant, and complementation lines (ppr596/ppr596: PPR596-FLAG) in pots. The homozygous ppr596 mutant exhibited slower growth and delayed maturation compared to both WT and complementation plants (first from the right, 75 days old). The bar in (B–D) indicates 2 cm.

Figure 3.
Analysis of mitochondrial gene transcripts in the ppr596 mutant. RT-PCR analysis of 33 mitochondrial protein-coding genes in the ppr596 mutant with WT and complemented plants. Actin expression was used as the loading control. Transcripts of nad2 (highlighted in red box) were severely decreased in the ppr596 mutant. The expression of some genes increased in the ppr596 mutant, as indicated by the asterisk.
Figure 3.
Analysis of mitochondrial gene transcripts in the ppr596 mutant. RT-PCR analysis of 33 mitochondrial protein-coding genes in the ppr596 mutant with WT and complemented plants. Actin expression was used as the loading control. Transcripts of nad2 (highlighted in red box) were severely decreased in the ppr596 mutant. The expression of some genes increased in the ppr596 mutant, as indicated by the asterisk.

Figure 4.
PPR596 is involved in the splicing of nad2 intron3. Splicing events of nad2 were examined in WT, ppr596, and complementation lines by RT-PCR. Actin expression was used as a loading control. (A) The structure of the nad2 pre-mRNA consists of two precursor RNA transcripts. The first precursor RNA consists of two exons and one intron, while the second consists of three exons and two introns. The maturation of nad2 mRNA requires proper splicing of three cis (introns 1, 3, and 4) and one trans (intron 2) intron splicing. Arrows indicate the positions of primers used to detect unspliced introns. (B) RT-PCR analysis revealed a significant decrease in the splicing efficiency of nad2 intron 3 in the ppr596 mutant, as highlighted by the red box. (C) RT-PCR analysis showing the accumulation of unspliced intron 3 in the nad2 transcript of the ppr596 mutant, with unspliced nad2 intron 3 highlighted by red box.
Figure 4.
PPR596 is involved in the splicing of nad2 intron3. Splicing events of nad2 were examined in WT, ppr596, and complementation lines by RT-PCR. Actin expression was used as a loading control. (A) The structure of the nad2 pre-mRNA consists of two precursor RNA transcripts. The first precursor RNA consists of two exons and one intron, while the second consists of three exons and two introns. The maturation of nad2 mRNA requires proper splicing of three cis (introns 1, 3, and 4) and one trans (intron 2) intron splicing. Arrows indicate the positions of primers used to detect unspliced introns. (B) RT-PCR analysis revealed a significant decrease in the splicing efficiency of nad2 intron 3 in the ppr596 mutant, as highlighted by the red box. (C) RT-PCR analysis showing the accumulation of unspliced intron 3 in the nad2 transcript of the ppr596 mutant, with unspliced nad2 intron 3 highlighted by red box.

Figure 5.
Confirmation of nad2 intron 3 splicing efficiency through quantitative RT-PCR. The nad4 and nad5 introns’ splicing was used as control. The relative accumulation of mRNA and pre-RNA transcripts was analyzed in both WT and ppr596 mutant plants. The histogram shows the splicing efficiencies, as indicated by log2 ratios of pre-RNA to mature RNA abundance in the ppr596 mutant compared to the WT plants. Asterisks indicate altered splicing of nad2 intron 3.
Figure 5.
Confirmation of nad2 intron 3 splicing efficiency through quantitative RT-PCR. The nad4 and nad5 introns’ splicing was used as control. The relative accumulation of mRNA and pre-RNA transcripts was analyzed in both WT and ppr596 mutant plants. The histogram shows the splicing efficiencies, as indicated by log2 ratios of pre-RNA to mature RNA abundance in the ppr596 mutant compared to the WT plants. Asterisks indicate altered splicing of nad2 intron 3.

Figure 6.
The loss of function of PPR596 affects the assembly of mitochondrial complex I. Mitochondrial proteins were isolated from 14-day seedlings of WT and complemented line, as well as one-month-old seedlings of ppr596, all grown in the dark. The proteins were solubilized with n-Dodecyl β-D-maltoside (β-DM), and mitochondrial membrane complexes were separated via BN-PAGE. (A) Coomassie Brilliant Blue (CCB) staining following electrophoresis confirms equal protein loading. The band representing complex I was reduced in the BN gel of the ppr596 mutant. (B) The in-gel activity assay of complex I indicated the severe decrease in complex I in the ppr596 mutant, with partial complementation observed in the complemented line. (C) Immunoblot analysis of BN-page using primary antibody for CA2, an early assembly factor of complex I. (D) Mitochondrion-encoded proteins L16 and COX1 were detected in WT, ppr596 mutant, and complementation line. Crude total mitochondrial proteins from WT, ppr596, and complementation line were separated by SDS page followed by transformation to the polyvinylidene difluoride (PVDF) membrane. Immunoblotting analyses used primary antibodies for L16, COX1, and CA2. Coomassie Brilliant Blue (CBB) stained gels were used as the sample loading control.
Figure 6.
The loss of function of PPR596 affects the assembly of mitochondrial complex I. Mitochondrial proteins were isolated from 14-day seedlings of WT and complemented line, as well as one-month-old seedlings of ppr596, all grown in the dark. The proteins were solubilized with n-Dodecyl β-D-maltoside (β-DM), and mitochondrial membrane complexes were separated via BN-PAGE. (A) Coomassie Brilliant Blue (CCB) staining following electrophoresis confirms equal protein loading. The band representing complex I was reduced in the BN gel of the ppr596 mutant. (B) The in-gel activity assay of complex I indicated the severe decrease in complex I in the ppr596 mutant, with partial complementation observed in the complemented line. (C) Immunoblot analysis of BN-page using primary antibody for CA2, an early assembly factor of complex I. (D) Mitochondrion-encoded proteins L16 and COX1 were detected in WT, ppr596 mutant, and complementation line. Crude total mitochondrial proteins from WT, ppr596, and complementation line were separated by SDS page followed by transformation to the polyvinylidene difluoride (PVDF) membrane. Immunoblotting analyses used primary antibodies for L16, COX1, and CA2. Coomassie Brilliant Blue (CBB) stained gels were used as the sample loading control.

Figure 7.
The alternative respiratory pathway is induced in the pp596 mutant. Quantitative RT-PCR results show the relative accumulation of alternative oxidase (AOX) and NADH dehydrogenase (NDA1, NDA2, NDB1) transcript in ppr596 mutant plants.
Figure 7.
The alternative respiratory pathway is induced in the pp596 mutant. Quantitative RT-PCR results show the relative accumulation of alternative oxidase (AOX) and NADH dehydrogenase (NDA1, NDA2, NDB1) transcript in ppr596 mutant plants.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sayyed, A.; Chen, B.; Wang, Y.; Cao, S.-K.; Tan, B.-C. PPR596 Is Required for nad2 Intron Splicing and Complex I Biogenesis in Arabidopsis. Int. J. Mol. Sci. 2024, 25, 3542. https://doi.org/10.3390/ijms25063542
AMA Style
Sayyed A, Chen B, Wang Y, Cao S-K, Tan B-C. PPR596 Is Required for nad2 Intron Splicing and Complex I Biogenesis in Arabidopsis. International Journal of Molecular Sciences. 2024; 25(6):3542. https://doi.org/10.3390/ijms25063542
Chicago/Turabian StyleSayyed, Aqib, Baoyin Chen, Yong Wang, Shi-Kai Cao, and Bao-Cai Tan. 2024. "PPR596 Is Required for nad2 Intron Splicing and Complex I Biogenesis in Arabidopsis" International Journal of Molecular Sciences 25, no. 6: 3542. https://doi.org/10.3390/ijms25063542
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.