
Quantitative Shotgun Proteomic Analysis of Bacteria after Overexpression of Recombinant Spider Miniature Spidroin, MaSp1
Abstract
:1. Introduction
2. Results
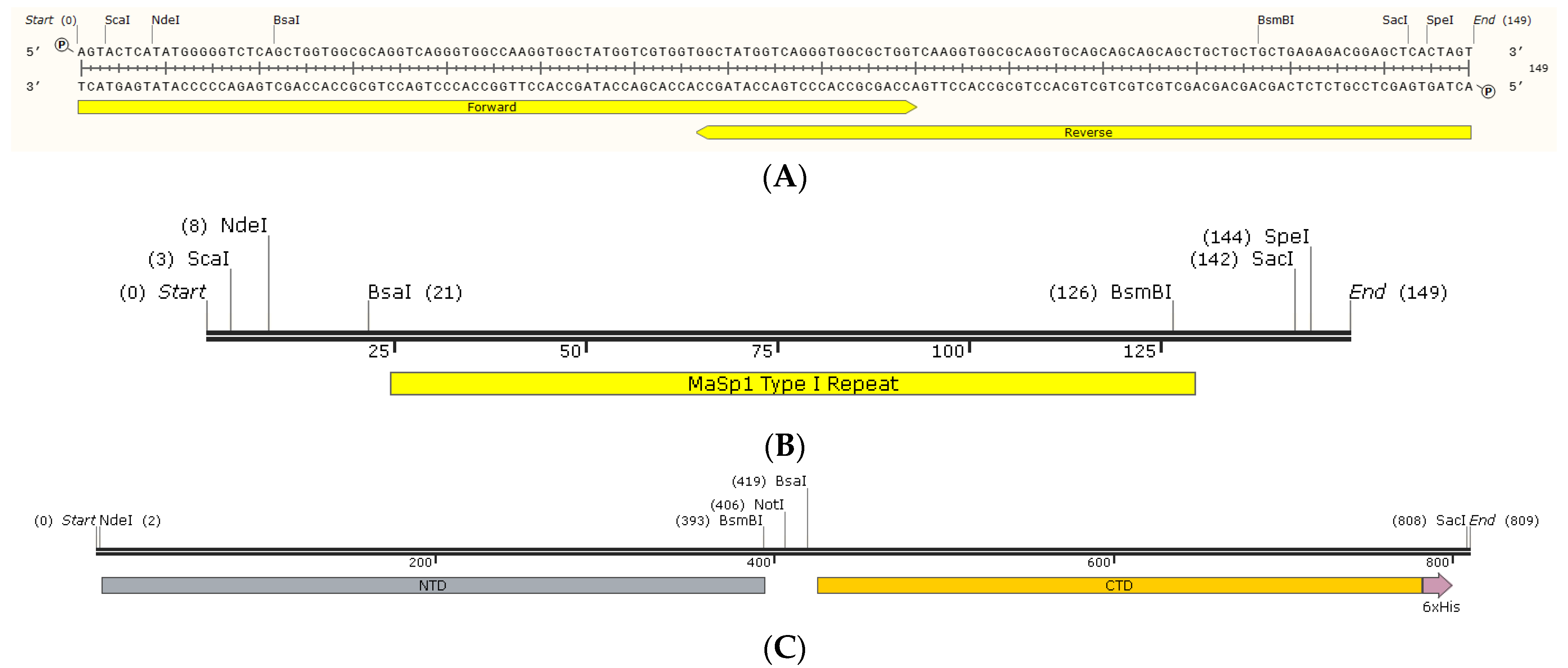
2.1. MaSp1 NTD-nx-CTD Constructs as Miniature Spidroins (Mini-Spidroins)
2.2. Expression of MaSp1 Mini-Spidroins in Bacteria
2.3. Quantitative Shotgun Proteomic Analysis of Bacteria Expressing MaSp1 Mini-Spidroins
3. Discussion
3.1. Experimental Strategy
3.2. Identified Targets
4. Materials and Methods
4.1. Site-Directed Mutagenesis of Parental pET24a Vector
4.2. Plasmid Construct Assembly
4.3. Seamless Cloning Strategy
4.4. Construction of NTD and CTD of MaSp1
4.5. MaSp1 Mini-Spidroin Design
4.6. Recombinant Protein Expression and Western Blot Analysis
4.7. In-Solution Tryptic Digestion
4.8. Mass Spectrometry and Chromatography
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gosline, J.M.; Guerette, P.A.; Ortlepp, C.S.; Savage, K.N. The mechanical design of spider silks: From fibroin sequence to mechanical function. J. Exp. Biol. 1999, 202, 3295–3303. [Google Scholar] [CrossRef] [PubMed]
- Larracas, C.; Hekman, R.; Dyrness, S.; Arata, A.; Williams, C.; Crawford, T.; Vierra, C.A. Comprehensive proteomic analysis of spider dragline silk from black widows: A recipe to build synthetic silk fibers. Int. J. Mol. Sci. 2016, 17, 1537. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Arakawa, K. Molecular mechanisms of the high performance of spider silks revealed through multi-omics analysis. Biophys. Physicobiol. 2023, 20, e200014. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lewis, R.V. Structure of a protein superfiber: Spider dragline silk. Proc. Natl. Acad. Sci. USA 1990, 87, 7120–7124. [Google Scholar] [CrossRef] [PubMed]
- Hinman, M.B.; Lewis, R.V. Isolation of a clone encoding a second dragline silk fibroin. Nephila clavipes dragline silk is a two-protein fiber. J. Biol. Chem. 1992, 267, 19320–19324. [Google Scholar] [CrossRef]
- Chaw, R.C.; Correa-Garhwal, S.M.; Clarke, T.H.; Ayoub, N.A.; Hayashi, C.Y. Proteomic evidence for components of spider silk synthesis from black widow silk glands and fibers. J. Proteome Res. 2015, 14, 4223–4231. [Google Scholar] [CrossRef]
- Ayoub, N.A.; Garb, J.E.; Tinghitella, R.M.; Collin, M.A.; Hayashi, C.Y. Blueprint for a high-performance biomaterial: Full-length spider dragline silk genes. PLoS ONE 2007, 2, e514. [Google Scholar] [CrossRef]
- Lewis, R.V. Spider silk: Ancient ideas for new biomaterials. Chem. Rev. 2006, 106, 3762–3774. [Google Scholar] [CrossRef]
- Beckwitt, R.; Arcidiacono, S. Sequence conservation in the C-terminal region of spider silk proteins (spidroin) from Nephila clavipes (tetragnathidae) and Araneus bicentenarius (araneidae). J. Biol. Chem. 1994, 269, 6661–6663. [Google Scholar] [CrossRef]
- Zhu, H.; Rising, A.; Johansson, J.; Zhang, X.; Lin, Y.; Zhang, L.; Yi, T.; Mi, J.; Meng, Q. Tensile properties of synthetic pyriform spider silk fibers depend on the number of repetitive units as well as the presence of N- and C-terminal domains. Int. J. Biol. Macromol. 2020, 154, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Fan, J.S.; Lai, C.C.; Raghuvamsi, P.V.; Chee, P.X.; Anand, G.S.; Yang, D. Structural basis of oligomerization of N-terminal domain of spider aciniform silk protein. Int. J. Mol. Sci. 2020, 21, 4466. [Google Scholar] [CrossRef]
- Askarieh, G.; Hedhammar, M.; Nordling, K.; Saenz, A.; Casals, C.; Rising, A.; Johansson, J.; Knight, S.D. Self-assembly of spider silk proteins is controlled by a pH-sensitive relay. Nature 2010, 465, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Hagn, F.; Thamm, C.; Scheibel, T.; Kessler, H. pH-dependent dimerization and salt-dependent stabilization of the N-terminal domain of spider dragline silk--implications for fiber formation. Angew. Chem. Int. Ed. Engl. 2011, 50, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Kronqvist, N.; Otikovs, M.; Chmyrov, V.; Chen, G.; Andersson, M.; Nordling, K.; Landreh, M.; Sarr, M.; Jornvall, H.; Wennmalm, S.; et al. Sequential pH-driven dimerization and stabilization of the N-terminal domain enables rapid spider silk formation. Nat. Commun. 2014, 5, 3254. [Google Scholar] [CrossRef] [PubMed]
- Sponner, A.; Vater, W.; Rommerskirch, W.; Vollrath, F.; Unger, E.; Grosse, F.; Weisshart, K. The conserved C-termini contribute to the properties of spider silk fibroins. Biochem. Biophys. Res. Commun. 2005, 338, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Copeland, C.G.; Bell, B.E.; Christensen, C.D.; Lewis, R.V. Development of a process for the spinning of synthetic spider silk. ACS Biomater. Sci. Eng. 2015, 1, 577–584. [Google Scholar] [CrossRef]
- Hsia, Y.; Gnesa, E.; Pacheco, R.; Kohler, K.; Jeffery, F.; Vierra, C. Synthetic spider silk production on a laboratory scale. J. Vis. Exp. 2012, 65, e4191. [Google Scholar]
- Teule, F.; Cooper, A.R.; Furin, W.A.; Bittencourt, D.; Rech, E.L.; Brooks, A.; Lewis, R.V. A protocol for the production of recombinant spider silk-like proteins for artificial fiber spinning. Nat. Protoc. 2009, 4, 341–355. [Google Scholar] [CrossRef]
- Vendrely, C.; Scheibel, T. Biotechnological production of spider-silk proteins enables new applications. Macromol. Biosci. 2007, 7, 401–409. [Google Scholar] [CrossRef]
- Heidebrecht, A.; Scheibel, T. Recombinant production of spider silk proteins. Adv. Appl. Microbiol. 2013, 82, 115–153. [Google Scholar]
- Fahnestock, S.R.; Bedzyk, L.A. Production of synthetic spider dragline silk protein in Pichia pastoris. Appl. Microbiol. Biotechnol. 1997, 47, 33–39. [Google Scholar] [CrossRef]
- Huemmerich, D.; Scheibel, T.; Vollrath, F.; Cohen, S.; Gat, U.; Ittah, S. Novel assembly properties of recombinant spider dragline silk proteins. Curr. Biol. 2004, 14, 2070–2074. [Google Scholar] [CrossRef]
- Menassa, R.; Zhu, H.; Karatzas, C.N.; Lazaris, A.; Richman, A.; Brandle, J. Spider dragline silk proteins in transgenic tobacco leaves: Accumulation and field production. Plant Biotechnol. J. 2004, 2, 431–438. [Google Scholar] [CrossRef]
- Lazaris, A.; Arcidiacono, S.; Huang, Y.; Zhou, J.F.; Duguay, F.; Chretien, N.; Welsh, E.A.; Soares, J.W.; Karatzas, C.N. Spider silk fibers spun from soluble recombinant silk produced in mammalian cells. Science 2002, 295, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Fahnestock, S.R.; Irwin, S.L. Synthetic spider dragline silk proteins and their production in Escherichia coli. Appl. Microbiol. Biotechnol. 1997, 47, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Dong, Q.; Yu, Y.; Niu, B.; Ji, D.; Li, M.; Huang, Y.; Chen, X.; Tan, A. Mass spider silk production through targeted gene replacement in Bombyx mori. Proc. Natl. Acad. Sci. USA 2018, 115, 8757–8762. [Google Scholar] [CrossRef]
- Schmuck, B.; Greco, G.; Barth, A.; Pugno, N.; Johansson, J.; Rising, A. High-yield production of a super-soluble miniature spidroin for biomimetic high-performance materials. Mater. Today 2021, 50, 16–23. [Google Scholar] [CrossRef]
- Xia, X.X.; Qian, Z.G.; Ki, C.S.; Park, Y.H.; Kaplan, D.L.; Lee, S.Y. Native-sized recombinant spider silk protein produced in metabolically engineered Escherichia coli results in a strong fiber. Proc. Natl. Acad. Sci. USA 2010, 107, 14059–14063. [Google Scholar] [CrossRef]
- Hu, X.; Kohler, K.; Falick, A.M.; Moore, A.M.; Jones, P.R.; Sparkman, O.D.; Vierra, C. Egg case protein-1. A new class of silk proteins with fibroin-like properties from the spider Latrodectus hesperus. J. Biol. Chem. 2005, 280, 21220–21230. [Google Scholar] [CrossRef]
- Burnand, B.; Kernan, W.N.; Feinstein, A.R. Indexes and boundaries for “quantitative significance” in statistical decisions. J. Clin. Epidemiol. 1990, 43, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Ping, L.; Zhang, H.; Zhai, L.; Dammer, E.B.; Duong, D.M.; Li, N.; Yan, Z.; Wu, J.; Xu, P. Quantitative proteomics reveals significant changes in cell shape and an energy shift after IPTG induction via an optimized silac approach for Escherichia coli. J. Proteome Res. 2013, 12, 5978–5988. [Google Scholar] [CrossRef]
- Wei, Y.; Lee, J.M.; Richmond, C.; Blattner, F.R.; Rafalski, J.A.; LaRossa, R.A. High-density microarray-mediated gene expression profiling of Escherichia coli. J. Bacteriol. 2001, 183, 545–556. [Google Scholar] [CrossRef]
- Chevallier, V.; Andersen, M.R.; Malphettes, L. Oxidative stress-alleviating strategies to improve recombinant protein production in Cho cells. Biotechnol. Bioeng. 2020, 117, 1172–1186. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. Alkyl hydroperoxide reductase is the primary scavenger of endogenous hydrogen peroxide in Escherichia coli. J. Bacteriol. 2001, 183, 7173–7181. [Google Scholar] [CrossRef]
- Dubbs, J.M.; Mongkolsuk, S. Peroxiredoxins in bacterial antioxidant defense. Subcell. Biochem. 2007, 44, 143–193. [Google Scholar]
- Schneider, B.L.; Hernandez, V.J.; Reitzer, L. Putrescine catabolism is a metabolic response to several stresses in Escherichia coli. Mol. Microbiol. 2013, 88, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, Y.; Zhan, W.; Wood, T.K.; Wang, X. Resistance to oxidative stress by inner membrane protein ElaB is regulated by OxyR and RpoS. Microb. Biotechnol. 2019, 12, 392–404. [Google Scholar] [CrossRef]
- Connor, A.; Wigham, C.; Bai, Y.; Rai, M.; Nassif, S.; Koffas, M.; Zha, R.H. Novel insights into construct toxicity, strain optimization, and primary sequence design for producing recombinant silk fibroin and elastin-like peptide in E. coli. Metab. Eng. Commun. 2023, 16, e00219. [Google Scholar] [CrossRef] [PubMed]
- Goemans, C.V.; Vertommen, D.; Agrebi, R.; Collet, J.F. Cnox is a chaperedoxin: A holdase that protects its substrates from irreversible oxidation. Mol. Cell 2018, 70, 614–627.e7. [Google Scholar] [CrossRef]
- Jones, P.G.; Inouye, M. The cold-shock response—A hot topic. Mol. Microbiol. 1994, 11, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-X.; Qian, Z.-G.; Zhong, J.-J.; Xia, X.-X. Hyper-production of large proteins of spider dragline silk masp2 by Escherichia coli via synthetic biology approach. Process Biochem. 2016, 51, 484–490. [Google Scholar] [CrossRef]
- Phadtare, S.; Severinov, K. Comparative analysis of changes in gene expression due to RNA melting activities of translation initiation factor IF1 and a cold shock protein of the CspA family. Genes. Cells 2009, 14, 1227–1239. [Google Scholar] [CrossRef]
- Clarke, T.H.; Garb, J.E.; Hayashi, C.Y.; Arensburger, P.; Ayoub, N.A. Spider transcriptomes identify ancient large-scale gene duplication event potentially important in silk gland evolution. Genome Biol. Evol. 2015, 7, 1856–1870. [Google Scholar] [CrossRef]
- Hernandez, S.B.; Cava, F. Environmental roles of microbial amino acid racemases. Environ. Microbiol. 2016, 18, 1673–1685. [Google Scholar] [CrossRef]
- Scior, A.; Preissler, S.; Kochh, M.; Deuerling, E. Directed PCR-free engineering of highly repetitive DNA sequences. BMC Biotechnol. 2011, 11, 87. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Repeats | Amino Acid Sequence |
|---|---|
| 2× | MQANTPWSSKANADAFINSFISAASNTGSFSQDQMEDMSLIGNTLMAAMDNMGGRITPSKLQALDMAFASSVAEIAASEGGDLGVTTNAIADALTSAFYQTTGVVNSRFISEIRSLIGMFAQASANDVYAAGGAGQGGQGGYGRGGYGQGGAGQGGAGAAAAAAAAGGAGQGGQGGYGRGGYGQGGAGQGGAGAAAAAAAAASGPGQIYYGPQSVAAPAAAAASALSAPATSARISSHASALLSNGPTNPASISNVISNAVSQISSSNPGASACDVLVQALLELVTALLTIIGSSNIGSVNYDSSGQYAQVVTQSVQNAFAHHHHHH |
| 4× | MQANTPWSSKANADAFINSFISAASNTGSFSQDQMEDMSLIGNTLMAAMDNMGGRITPSKLQALDMAFASSVAEIAASEGGDLGVTTNAIADALTSAFYQTTGVVNSRFISEIRSLIGMFAQASANDVYAAGGAGQGGQGGYGRGGYGQGGAGQGGAGAAAAAAAAGGAGQGGQGGYGRGGYGQGGAGQGGAGAAAAAAAAGGAGQGGQGGYGRGGYGQGGAGQGGAGAAAAAAAAGGAGQGGQGGYGRGGYGQGGAGQGGAGAAAAAAAAASGPGQIYYGPQSVAAPAAAAASALSAPATSARISSHASALLSNGPTNPASISNVISNAVSQISSSNPGASACDVLVQALLELVTALLTIIGSSNIGSVNYDSSGQYAQVVTQSVQNAFAHHHHHH |
| Gene | Protein | Acc. # | FCR | p-Value | Function |
|---|---|---|---|---|---|
| lacA | Galactoside O-acetyltransferase * | P07464 | +OLR | 7.89 × 10−19 | Lactose metabolism |
| lacZ | Beta-galactosidase * | P00722 | +OLR +270.33 | 1.00 × 10−20 | Lactose metabolism |
| N/A | MaSp1 NTD-2x-CTD | N/A | +OLR +12.09 | 1.00 × 10−20 | Target protein |
| ybgl | GTP cyclohydrolase 1 type 2 homolog | P0AFP6 | +3.57 | 1.26 × 10−3 | O.S. resistance |
| melA | Alpha-galactosidase * | P06720 | +2.30 | 3.59 × 10−5 | Lactose metabolism |
| pnp | Polyribonucleotide nucleotidyltransferase | P05055 | +1.94 | 3.08 × 10−14 | mRNA degradation |
| slyD | FKBP-type peptidyl-prolyl cis-trans isomerase SlyD | P0A9K9 | +1.52 | 2.52 × 10−3 | Chaperone |
| ytfQ | Galactofuranose-binding protein YtfQ | P39325 | −1.64 | 1.04 × 10−8 | Sugar transport |
| gatB | PTS system galactitol-specific EIIB component | P37188 | −1.82 | 1.87 × 10−4 | Sugar transport |
| pliG | Inhibitor of g-type lysozyme | P76002 | −3.33 | 4.28 × 10−4 | Inhibitor |
| mdoG | Glucans biosynthesis protein G | P33136 | −4.20 | 1.06 × 10−3 | Cell wall synthesis |
| galS | HTH-type transcriptional regulator GalS | P25748 | −6.50 | 1.47 × 10−11 | Repressor of mgl operon |
| mglA | Gal./methyl galactoside import ATP-binding protein * | P0AAG8 | −12.00 | 1.53 × 10−3 | Lactose metabolism |
| mglB | D-galactose-binding periplasmic protein * | P0AEE5 | −OLR | 1.39 × 10−9 | Lactose metabolism |
| Gene | Protein | Acc. # | p-Value | FCR | Function |
|---|---|---|---|---|---|
| lacA | Galactoside O-acetyltransferase * | P07464 | 1.03 × 10−14 | +OLR | Lactose metabolism |
| qorB | Quinone oxidoreductase 2 | P39315 | 8.38 × 10−4 | +OLR | Electron transport in respiratory chain |
| cnoX | Chaperedoxin | P77395 | 8.38 × 10−4 | +OLR | Protein deaggregation |
| dadX | Alanine racemase, catabolic | P29012 | 5.79 × 10−3 | +OLR | Isomerizes L- to D-alanine |
| yiaF | Uncharacterized protein YiaF | P0ADK0 | 1.60 × 10−3 | +OLR | Uncharacterized |
| panB | 3-methyl-2-oxobutanoate hydroxymethyltransferase | P31057 | 1.60 × 10−3 | +OLR | Pantothenate biosynthesis |
| yajG | Uncharacterized lipoprotein YajG | P0ADA5 | 5.79 × 10−3 | +OLR | Uncharacterized |
| N/A | MaSp1 NT4XCT | N/A | 1.00 × 10−20 | +OLR | Target protein |
| lacZ | Beta-galactosidase* | P00722 | 1.00 × 10−20 | +OLR | Lactose metabolism |
| yfcZ | UPF0381 protein YfcZ | P0AD33 | 9.29 × 10−4 | +14.00 | Uncharacterized |
| wzzB | Chain length determinant protein | P76372 | 5.22 × 10−3 | +11.00 | Lipopolysaccharide biosynthesis |
| kdsB | 3-deoxy-manno-octulosonate cytidylyltransferase | P04951 | 9.18 × 10−3 | +10.00 | Lipopolysaccharide biosynthesis |
| efeO | Iron uptake system component EfeO | P0AB24 | 9.18 × 10−3 | +10.00 | Iron uptake |
| metQ | D-methionine-binding lipoprotein MetQ | P28635 | 1.68 × 10−9 | +9.80 | Methionine transport system |
| ydgJ | Uncharacterized oxidoreductase YdgJ | P77376 | 1.60 × 10−2 | +9.00 | Uncharacterized |
| osmC | Peroxiredoxin OsmC | P0C0L2 | 7.66 × 10−4 | +8.50 | Hydroperoxide metabolism |
| cdd | Cytidine deaminase | P0ABF6 | 1.18 × 10−4 | +7.67 | Pyrimidine salvage pathway |
| ycdY | Chaperone protein YcdY | P75915 | 2.24 × 10−3 | +7.50 | Molybdoenzyme chaperone |
| cyoB | Cytochrome bo(3) ubiquinol oxidase subunit | P0ABI8 | 4.95 × 10−7 | +6.83 | Terminal enzyme in respiratory chain |
| speB | Agmatinase | P60651 | 8.45 × 10−6 | +6.60 | Putrescine biosynthesis |
| ushA | Protein UshA | P07024 | 2.42 × 10−4 | +6.00 | Nucleotidase |
| yicC | UPF0701 protein YicC | P23839 | 3.99 × 10−4 | +5.75 | sRNA degradation |
| ybgl | GTP cyclohydrolase 1 type 2 homolog | P0AFP6 | 2.71 × 10−5 | +5.50 | Oxidative stress resistance |
| yfcE | Phosphodiesterase YfcE | P67095 | 1.68 × 10−4 | +5.40 | Phosphodiesterase |
| nagA | N-acetylglucosamine-6-phosphate deacetylase | P0AF18 | 4.38 × 10−4 | +5.00 | Peptidoglycan synthesis |
| acrB | Multidrug efflux pump subunit AcrB | P31224 | 6.89 × 10−3 | +5.00 | Transporter |
| gpr | L-glyceraldehyde 3-phosphate reductase | Q46851 | 4.80 × 10−5 | +4.86 | Oxidoreductase activity |
| btsT | Pyruvate/proton symporter BtsT | P39396 | 4.80 × 10−5 | +4.86 | Pyruvate transport |
| iscX | Protein IscX | P0C0L9 | 2.75 × 10−3 | +4.75 | Iron–sulfur cluster assembly |
| gor | Glutathione reductase | P06715 | 1.76 × 10−3 | +4.00 | Oxidative stress prevention |
| fmt | Methionyl-tRNA formyltransferase | P23882 | 8.04 × 10−5 | +3.90 | Initiation of protein synthesis |
| gnsA | Protein GnsA | P0AC92 | 1.92 × 10−4 | +3.89 | Membrane lipid synthesis |
| maeA | NAD-dependent malic enzyme | P26616 | 6.69 × 10−3 | +3.80 | Oxidation–reduction |
| eptA | Phosphoethanolamine transferase EptA | P30845 | 1.03 × 10−2 | +3.60 | Membrane synthesis |
| grcA | Autonomous glycyl radical cofactor | P68066 | 6.75 × 10−4 | +3.56 | Glucose fermentation |
| zntA | Zinc/cadmium/lead-transporting P-type ATPase | P37617 | 1.56 × 10−2 | +3.40 | Metal ion transporter |
| melA | Alpha-galactosidase * | P06720 | 1.67 × 10−7 | +3.37 | Lactose metabolism |
| mtlD | Mannitol-1-phosphate 5-dehydrogenase | P09424 | 5.98 × 10−4 | +3.27 | Oxidoreductase |
| iscA | Iron-binding protein IscA | P0AAC8 | 5.55 × 10−4 | +3.17 | Iron-binding protein |
| rihC | Non-specific ribonucleoside hydrolase RihC | P22564 | 1.42 × 10−2 | +3.17 | Ribonucleoside cleavage |
| yhhX | Uncharacterized oxidoreductase YhhX | P46853 | 8.68 × 10−3 | +3.14 | Uncharacterized |
| gpml | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase | P37689 | 1.55 × 10−7 | +3.03 | Glycolysis |
| lpp | Major outer membrane lipoprotein Lpp | P69776 | 9.98 × 10−8 | +3.03 | Outer membrane lipoprotein |
| yfcD | Uncharacterized Nudix hydrolase YfcD | P65556 | 1.28 × 10−2 | +3.03 | Uncharacterized |
| cspA | Cold shock protein CspA | P0A9X9 | 1.19 × 10−4 | +3.00 | RNA chaperone |
| carA | Carbamoyl-phosphate synthase small chain | P0A6F1 | 1.87 × 10−4 | +3.00 | Arginine and pyrimidine biosynthesis |
| dcrB | Inner membrane lipoprotein DcrB | P0AEE1 | 7.85 × 10−3 | +3.00 | Membrane fluidity and homeostasis |
| hslO | 33 kDa chaperonin | P0A6Y5 | 1.89 × 10−3 | +3.00 | Oxidative stress chaperone |
| trxC | Thioredoxin 2 | P0AGG4 | 1.28 × 10−2 | +3.00 | Oxidative stress response |
| malM | Maltose operon periplasmic protein | P03841 | 1.89 × 10−3 | +3.00 | Maltose transport |
| spy | Periplasmic chaperone Spy | P77754 | 1.45 × 10−2 | −3.25 | Chaperone |
| galS | HTH-type transcriptional regulator GalS | P25748 | 1.62 × 10−7 | −3.60 | Repression of mgl operon |
| dnaX | DNA polymerase III subunit tau | P06710 | 6.16 × 10−3 | −4.33 | DNA replication |
| ppk | Polyphosphate kinase | P0A7B1 | 1.25 × 10−2 | −5.00 | Polyphosphate synthesis |
| mglA | Galactose/methyl galactoside import ATP-binding protein MglA * | P0AAG8 | 6.91 × 10−3 | −5.50 | Galactose import |
| pliG | Inhibitor of g-type lysozyme | P76002 | 2.98 × 10−6 | −6.00 | Inhibits g-type lysozyme |
| hrpA | ATP-dependent RNA helicase HrpA | P43329 | 9.59 × 10−4 | −12.00 | RNA helicase |
| mglB | D-galactose-binding periplasmic protein * | P0AEE5 | 2.11 × 10−11 | −OLR | Lactose metabolism |
| rnpA | Ribonuclease P protein component | P0A7Y8 | 5.44 × 10−3 | −OLR | tRNA processing |
| mntR | Transcriptional regulator MntR | P0A9F1 | 5.44 × 10−3 | −OLR | Manganese transport regulation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randene, K.; Mendoza, J.A.H.; Ysit, M.; Vierra, C. Quantitative Shotgun Proteomic Analysis of Bacteria after Overexpression of Recombinant Spider Miniature Spidroin, MaSp1. Int. J. Mol. Sci. 2024, 25, 3556. https://doi.org/10.3390/ijms25063556
Randene K, Mendoza JAH, Ysit M, Vierra C. Quantitative Shotgun Proteomic Analysis of Bacteria after Overexpression of Recombinant Spider Miniature Spidroin, MaSp1. International Journal of Molecular Sciences. 2024; 25(6):3556. https://doi.org/10.3390/ijms25063556
Chicago/Turabian StyleRandene, Kathryn, J Alexander Hoang Mendoza, Michael Ysit, and Craig Vierra. 2024. "Quantitative Shotgun Proteomic Analysis of Bacteria after Overexpression of Recombinant Spider Miniature Spidroin, MaSp1" International Journal of Molecular Sciences 25, no. 6: 3556. https://doi.org/10.3390/ijms25063556