
VvJAZ13 Positively Regulates Cold Tolerance in Arabidopsis and Grape

College of Horticulture, Gansu Agricultural University, Lanzhou 730070, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(8), 4458; https://doi.org/10.3390/ijms25084458
Submission received: 11 March 2024
/
Revised: 14 April 2024
/
Accepted: 17 April 2024
/
Published: 18 April 2024
(This article belongs to the Section Molecular Plant Sciences)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Cold stress adversely impacts grape growth, development, and yield. Therefore, improving the cold tolerance of grape is an urgent task of grape breeding. The Jasmonic acid (JA) pathway responsive gene JAZ plays a key role in plant response to cold stress. However, the role of JAZ in response to low temperatures in grape is unclear. In this study, VvJAZ13 was cloned from the ‘Pinot Noir’ (Vitis vinefera cv. ‘Pinot Noir’) grape, and the potential interacting protein of VvJAZ13 was screened by yeast two-hybrid (Y2H). The function of VvJAZ13 under low temperature stress was verified by genetic transformation. Subcellular localization showed that the gene was mainly expressed in cytoplasm and the nucleus. Y2H indicated that VvF-box, VvTIFY5A, VvTIFY9, Vvbch1, and VvAGD13 may be potential interacting proteins of VvJAZ13. The results of transient transformation of grape leaves showed that VvJAZ13 improved photosynthetic capacity and reduced cell damage by increasing maximum photosynthetic efficiency of photosystem II (Fv/Fm), reducing relative electrolyte leakage (REL) and malondialdehyde (MDA), and increasing proline content in overexpressed lines (OEs), which played an active role in cold resistance. Through the overexpression of VvJAZ13 in Arabidopsis thaliana and grape calli, the results showed that compared with wild type (WT), transgenic lines had higher antioxidant enzyme activity and proline content, lower REL, MDA, and hydrogen peroxide (H2O2) content, and an improved ability of scavenging reactive oxygen species. In addition, the expression levels of CBF1-2 and ICE1 genes related to cold response were up-regulated in transgenic lines. To sum up, VvJAZ13 is actively involved in the cold tolerance of Arabidopsis and grape, and has the potential to be a candidate gene for improving plant cold tolerance.
1. Introduction
Low temperature stress is one of the main environmental factors affecting plant growth and development. The response to low temperature stress and the breeding of cold tolerant plants have attracted wide attention in recent years [1]. Freezing or extremely low temperature is the key factor affecting plant growth and development and crop yield. Plants have evolved a mechanism to increase their tolerance to freezing during exposure to low but non-freezing temperatures, a phenomenon known as cold acclimation [2]. Grape (Vitis vinifera L.) is a kind of delicious and nutritious fruit, which has a long history of cultivation and is still one of the fruits with the highest yield in the world. Grape is mainly distributed in arid or semi-arid areas with latitude 25–45° north, and have poor adaptability to the low temperature and arid climate in the main planting areas [3]. Therefore, it is of great practical significance to identify and explore new genes of cold resistance in grape and provide resources for breeding new varieties of cold resistance.
Under low temperature stress, transcription factors (TF) also respond to cold signals. Among them, C-repeat Binding Factor (CBFs; also known as dehydration-responsive element-binding proteins or DREBs) transcription factor plays an important role in the response to low temperature stress, and it is identified as the key response factor of plant low temperature stress under cold stress [4,5]. The expression level of the CBF/DREB1-like (C-repeat/dehydration-responsive element binding transcription factors) gene CBF1-4 was increased and maintained at a certain level during low temperature stress. This gene was isolated from the Riparian grape (Vitis vulpina L.) and the Eurasian grape, and the analysis suggested that the cold resistance of grape may be related to this gene [6,7]. Among them, the cold resistance of transgenic grape overexpressing VvCBF4 was significantly improved [8]. In the CBF-dependent pathway, CBF protein recognizes the CRT/DRE (C repeat/dehydration-responsive element) cis element in the promoter region of the COR (cold regulated gene, COR) gene, which in turn activates the transcription of these downstream genes and leads to cold storage and freezing tolerance in plants. ICE1 (Inducer of CBF expression 1) TF is located in the upstream of the CBF gene. Under low temperature stress, ICE1 can activate the expression of the CBF3 gene [9]. The activated CBF binds to the C-repeat/DRE element on the promoter of the CORs gene. For example, heterologous expression of grape VrCBF1 and VrCBF4 can promote the expression of AtCOR6.6, AtCOR15a, and AtCOR47 genes in Arabidopsis, and positively regulate the expression of AtICE1, thus enhancing the cold resistance of the plant [10]. The expression of VvICE1a and VvICE1b improved the cold tolerance of transgenic Arabidopsis and led to the up-regulation of stress-related genes AtRD29A (Stress-inducible promoter) and AtCOR47 [11]. In addition, the cold resistance function of transcription factor genes such as VaERF092 (cold-responsive ethylene response factor), VaAQUILO (AcQUIred tolerance to low temperatures), VbWRKY32 (transcription factors), and VaPAT1 (a microtubule-interacting protein) in grape was also verified [12,13,14,15].
Recent studies have shown that JA is related to leaf senescence and cold resistance [16]. Exogenous JA promotes leaf senescence and expresses genes related to leaf senescence, thus improving the freezing resistance of Arabidopsis thaliana. JA can also improve the cold tolerance of Arabidopsis by regulating cold response genes downstream of the CBF pathway [17]. JAZ is a key gene in the JA pathway. Many negative regulators of the JA signal can be suppressed, such as bHLH (basic helix-loop-helix) subgroup IIId transcription factors [18,19]. During extracellular stimulation or growth and development, JA-Ile enhances the interaction between COI1 (CORONATINE INSENSITIVE 1) and JAZ family proteins, which leads to the ubiquitination and degradation of JAZ by 26S proteasome [20,21]. JAZ proteins belong to a large plant-specific TIFY family. The TIFY family contains a highly conserved ZIM (TIFY) domain. JAZ proteins contain an N-terminal domain and a highly conserved C-terminal Jas domain, which can mediate interactions with COI1 and multiple transcription factors. Among them, the ZIM domain helps JAZ proteins to form dimers and mediates the interaction between JAZ proteins and co-suppressor NINJA (Noval interactor of JAZ). JAZ protein inhibits downstream target genes through Jas domain and negatively regulates the expression of downstream target genes [22,23].
In Arabidopsis, JAZ protein directly interacts with MYB21 and MYB24, which can regulate the model of JA biosynthesis to trigger COI1-dependent JAZ degradation to control MYB21 and MYB24 levels, thereby controlling stamen development [24]. In Arabidopsis, AtJAZ3 targeting proteins TOE2 (TARGET OF EAT 2) and KAN1 (KANADI1) interact directly with the inhibitor TPL (TOPLESS). These JAZ targets seem to be mainly transcription factors related to the hormone regulation of plant development and light signal transduction [25]. Photosensitive pigments and JAZ proteins are involved in the interaction between light and JA signals, thus affecting plant photosynthesis [26]. The coordination of JA signal crosstalk may be one of the important functions of JAZ protein. Recent studies have shown that rice (Oryza) JAZ9 plays a key role in mediating crosstalk between JA and gibberellin (GA). It interacts with DELLA protein (SLENDER RICE 1). Overexpression of OsJAZ9 enhanced the response of rice to GA [27]. In fact, JA is a positive regulator of leaf senescence because JAZ7 protein inhibits dark-induced leaf senescence, while MYC transcription factors promote senescence by activating senescence-related genes and chlorophyll degradation-related genes, indicating that JA stimulates leaf senescence in a COI1-dependent manner. In this pathway, JAZ4, JAZ8, and WRKY57 act as negative regulatory factors, while IAA29, an auxin signal transduction inhibitor, acts as a positive regulatory factor in contrast to JAZ7 [28]. JAZ also inhibits the transcriptional activity of ETHYLENE-INSENSITIVE 3 (EIN3)/EIN3-LIKE (EIL) downstream of APETALA2/ETHYLENE RESPONSE FACTOR (AP2/ERF) domain transcription factor ORA59 (ORA59/ERF1) gene in ethylene pathway and induces its expression, thus resisting the infection of necrotizing pathogens and nutritional pathogens [29].
It is found that the expression of the JAZ gene is regulated by various abiotic stresses such as cold, saline-alkali, drought and so on. In mosses (Racomitrium canescens), overexpression of the JAZ1 gene can enhance salt stress tolerance and inhibit the expression of genes related to abscisic acid (ABA) pathway [30]. In peppermint (Mentha Canadensis) leaves and roots, the expression of McJAZ8 was up-regulated to varying degrees under drought and NaCl treatments, indicating that McJAZ8 was involved in the response of peppermint to severe cold and salt stress [31]. In Arabidopsis, the JAZ gene enhances the drought tolerance of Arabidopsis by regulating redox, plant hormones, and secondary metabolites [32]. In rice, overexpression of OsJAZ1 can enhance the drought resistance of rice, and overexpression of OsJAZ9 can enhance salt and alkali tolerance of rice plants [17]. The expression of the GhJAZ1 gene in cotton (Gossypium hirsutum L.) was up-regulated under low temperature stress, indicating that the gene was involved in the response to low temperature stress [33]. The heterologous expression of soybean (Glycine max) GsJAZ2 gene in Arabidopsis can enhance the tolerance of Arabidopsis to saline-alkali stress [34]. In all, these studies show that the JAZ gene family plays a positive role in plant response to cold stress, and VvJAZ13 may be an important candidate gene to improve stress resistance in grape molecular breeding.
In this study, we identified VvJAZ13 from the ‘Pinot Noir’ grape. The effect of VvJAZ13 on cold stress was examined in transgenic Arabidopsis and grape calli. In addition, we also studied the interaction protein of VvJAZ13 in grape and the physiological and biochemical indexes of Arabidopsis and grape calli overexpressed by VvJAZ13 under cold stress.
2. Results
2.1. Cloning and Phylogenetic Analysis of the VvJAZ13 Gene
According to the previous transcriptome data, in the experiment, we cloned the GSVIVG01015042001 gene from the ‘Pinot Noir’ grape and renamed it VvJAZ13 on the basis of cloning and chromosome mapping of the grape gene (Figures S1 and S2). Based on the phylogenetic tree construction of the grape, Arabidopsis, apple, and tomato JAZ gene family members (Figure 1), these four species can be divided into four subfamilies. The VvJAZ13 gene clustered in III subgroup and is closely related to apple and tomato.
2.2. Transient Expression of VvJAZ13 in Tobacco
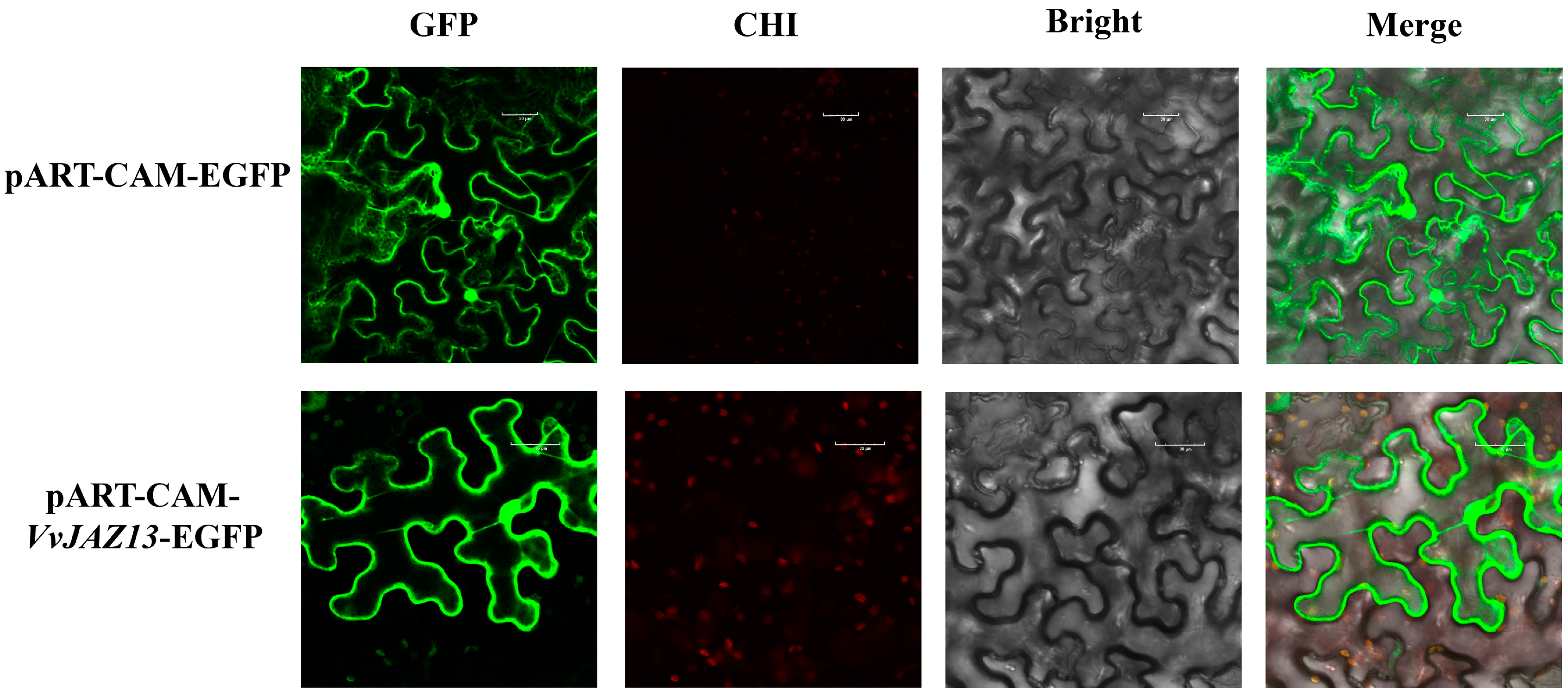
We constructed the pART-CAM-VvJAZ13-EGFP vector. The empty plasmids of pART-CAM-VvJAZ13-EGFP and pART-CAM-EGFP were transformed into Agrobacterium tumefaciens GV3101 and injected into tobacco leaves. The results showed that VvJAZ13 was localized in the cytoplasm and nucleus (Figure 2).
2.3. The Interaction Proteins Screening of VvJAZ13
According to the CDS sequence of VvJAZ13, pGBKT7-VvJAZ13 primers were designed and amplified (Table S1). The pGBKT7-VvJAZ13 recombinant plasmid was introduced into yeast receptor cells Y2H gold (Figure S3), which were coated on SD/-Trp and SD/-Trp/-Leu/-His plates, respectively, and was found not to grow on SD/-Trp/-Leu/-His plates, suggesting that no self-activation was detected for VvJAZ13 (Figure S4).
Using pGBKT7-VvJAZ13 as bait protein, the monoclonal strain was screened on an SD/-Trp/-Leu/-His/-Ade plate by Mating double hybridization. Through blast comparison in NCBI, it was found that most of the genes were related to abiotic stress. For example, VvF-box (core subunit of E3 ubiquitin ligase SCF complex), VvTIFY5A (JA signal regulatory protein), VvTIFY9 (JA signal regulatory protein), Vvbch1 (β-carotene hydroxylase), VvAGD13 (ADP-ribose factor GTP enzyme activating protein AGD13) (Figure S5).
The target gene fragments of the same size were obtained by PCR amplification with pGADT7-VvF-box, pGADT7-VvTIFY5A, pGADT7-VvTIFY9, pGADT7-Vvbch1, and pGADT7-VvAGD13 primers (Table S1 and Figure S6). The recombinant plasmids BD-VvJAZ13/AD-VvF-box, BD-VvJAZ13/AD-VvTIFY5A, BD-VvJAZ13/AD-VvTIFY9, BD-VvJAZ13/AD-Vvbch1, and BD-VvJAZ13/AD-VvAGD13 were co-transformed into Y2Hgold yeast competent cells and verified. BD/AD, BD-VvJAZ13/AD, BD/AD-VvF-box, BD/AD-VvTIFY5A, BD/AD-VvTIFY9, BD/AD-Vvbch1, and BD/AD-VvAGD1 co-transduction was used as control. They were spread on DDO, DDO + X, TDO, TDO + X, QDO, and QDO + X plates, respectively. It was found that BD-VvJAZ1/AD-VvF-box, BD-VvJAZ1/AD-VvTIFY5A, and BD-VvJAZ1/AD-VvTIFY9 grew and turned blue in QDO + X, indicating that there was an interaction between VvJAZ13 and VvF-box, VvTIFY5A, and VvTIFY9. BD-VvJAZ1/AD-Vvbch1 and BD-VvJAZ1/AD-VvAGD13 did not grow in QDO but grew and turned blue in TDO + X, indicating that there was a weak interaction between VvJAZ13 and Vvbch1 and VvAGD13 (Figure 3).
2.4. The Instantaneous Transformation of VvJAZ13 Improved the Cold TOLERANCE of Grape Leaves
Gene VvJAZ13 was overexpressed in grape leaves by the Agrobacterium-mediated transient transformation system to characterize the gene function under cold stress. The results showed that the fluorescence intensity of grape leaves overexpressing VvJAZ13-OE was significantly higher than that of WT grape leaves under cold-stress treatment (Figure 4A).
Under non-stress treatment, the transcriptional level of VvJAZ13-OE grape leaves was higher than that of WT grape leaves, indicating that VvJAZ13 was successfully transformed into grape leaves. Under cold-stress treatment, the expression of VvJAZ13 in overexpressed grape leaves was significantly higher than that in WT grape leaves (Figure 4B). Thereafter, the VvJAZ13-OE and WT leaves were exposed to 4 °C for 24 h. Consistent with fluorescence intensity, REL and MDA contents in VvJAZ13-OE grape leaves decreased by 20.37% and 28.57% compared with WT leaves, respectively, while proline contents increased by 8.82% compared with WT leaves (Figure 4C–E). Compared with WT leaves, the Fv/Fm and SPAD values of VvJAZ13-OE leaves increased by 20.83% and 24.26%, respectively (Figure 4F,G).
Subsequently, the transcript abundances of VvCBF1, VvCBF2, VvCBF3, ICE1a, ICE1b, and ICE1c were examined. The results showed that VvJAZ13 overexpression promoted the expression of VvCBF and VvICE1 genes, especially the VvCBF3 gene under cold stress, which was 38.02% higher than that of WT. These results indicated that overexpression of VvJAZ13 improved the cold resistance of the grape leaves (Figure S7).
2.5. Overexpression of VvJAZ13 Increased Cold Tolerance in Transgenic Arabidopsis
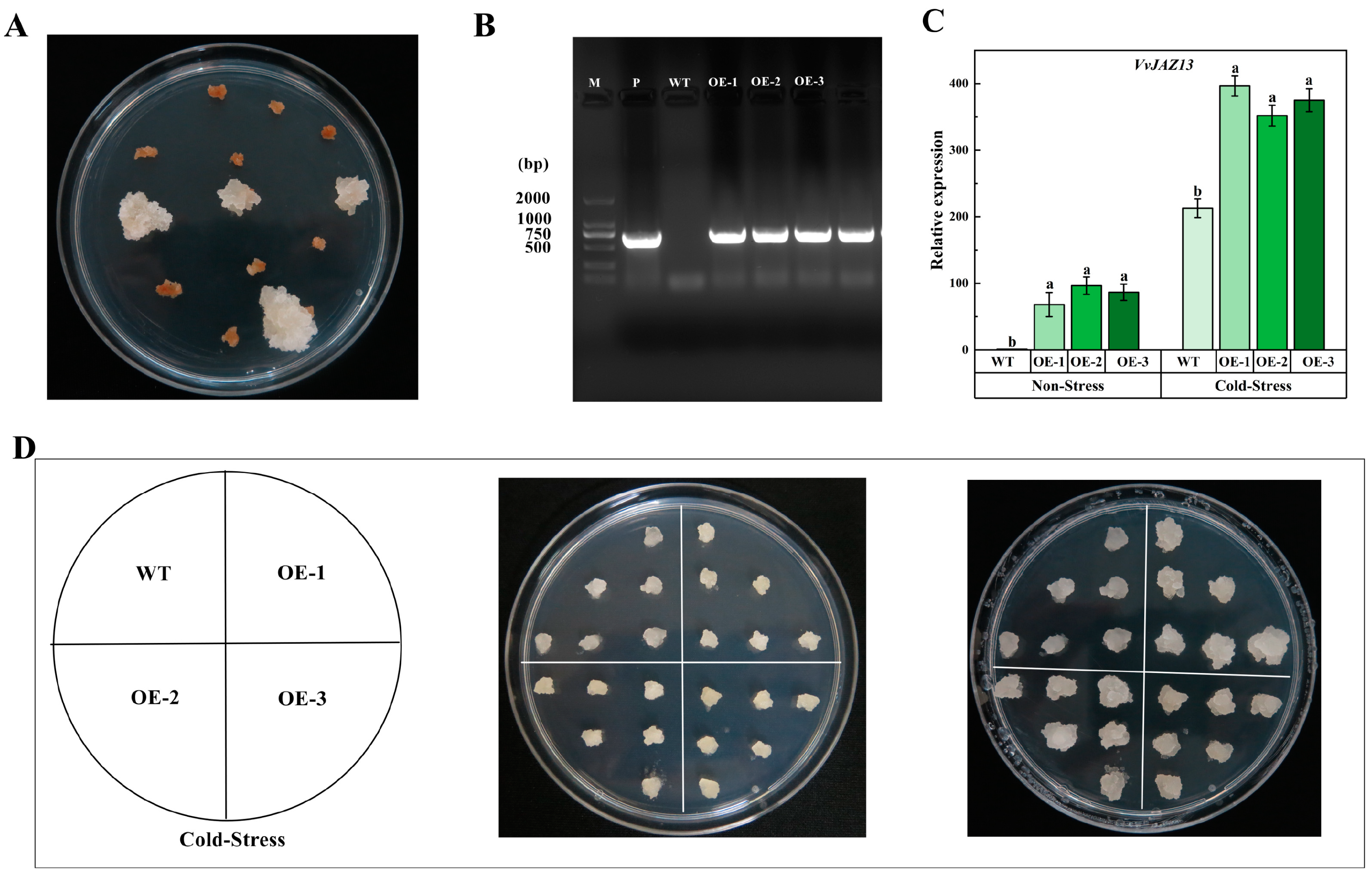
The transformed Arabidopsis seeds were screened on the MS medium containing Kana, and the Arabidopsis lines with overexpression of VvJAZ13 were obtained (Figure S8A,B). Three transgenic lines OE-1, OE-2, and OE-3 were selected for functional verification (Figure S8C).
The results showed that cold stress of WT was more serious than that of OEs. One week after recovery, the survival rate of OEs was significantly higher than that of WT (Figure 5A). qRT-PCR analysis of Arabidopsis showed that the expression of the VvJAZ13 gene was significantly up-regulated under cold stress treatment (Figure 5B).
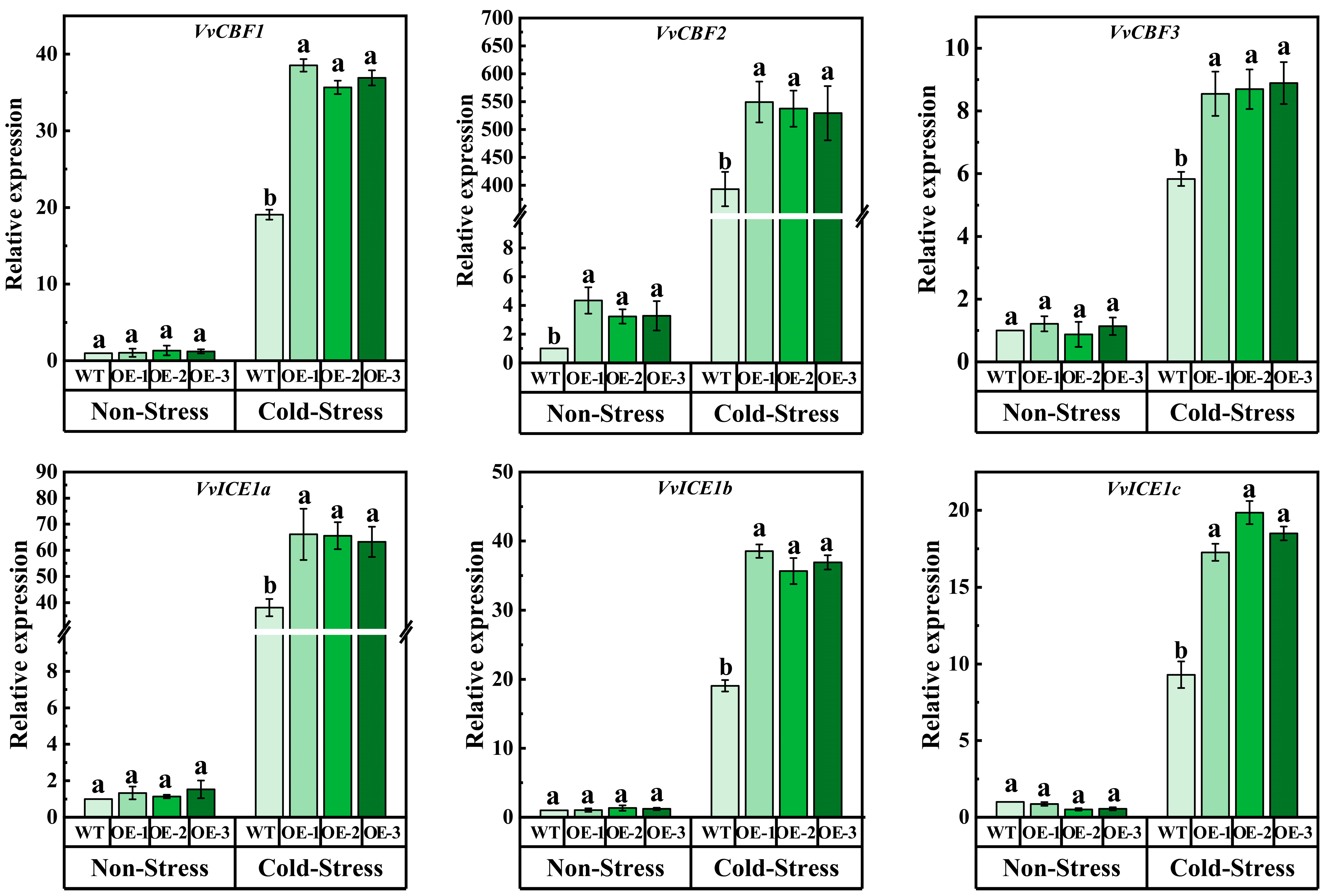
Analysis of the changes of cold response genes AtCBF and AtICE1 in transgenic Arabidopsis is shown in Figure 5C. The results showed that there was no significant difference in the expression of AtCBF1, AtCBF2, AtCBF3, and AtICE1 between transgenic VvJAZ13 Arabidopsis and WT before stress. However, under cold stress, the expression of AtCBF1, AtCBF2, and AtCBF3 in transgenic Arabidopsis was significantly higher than that in WT; especially, the expression of AtCBF3 in OEs increased by 181.11%, 262.22%, and 215.56%, respectively, compared with WT (Figure 5C). The results indicated that overexpression of VvJAZ13 can lead to upregulation of AtCBF1-3 and AtICE1 gene expression in Arabidopsis under cold stress conditions.
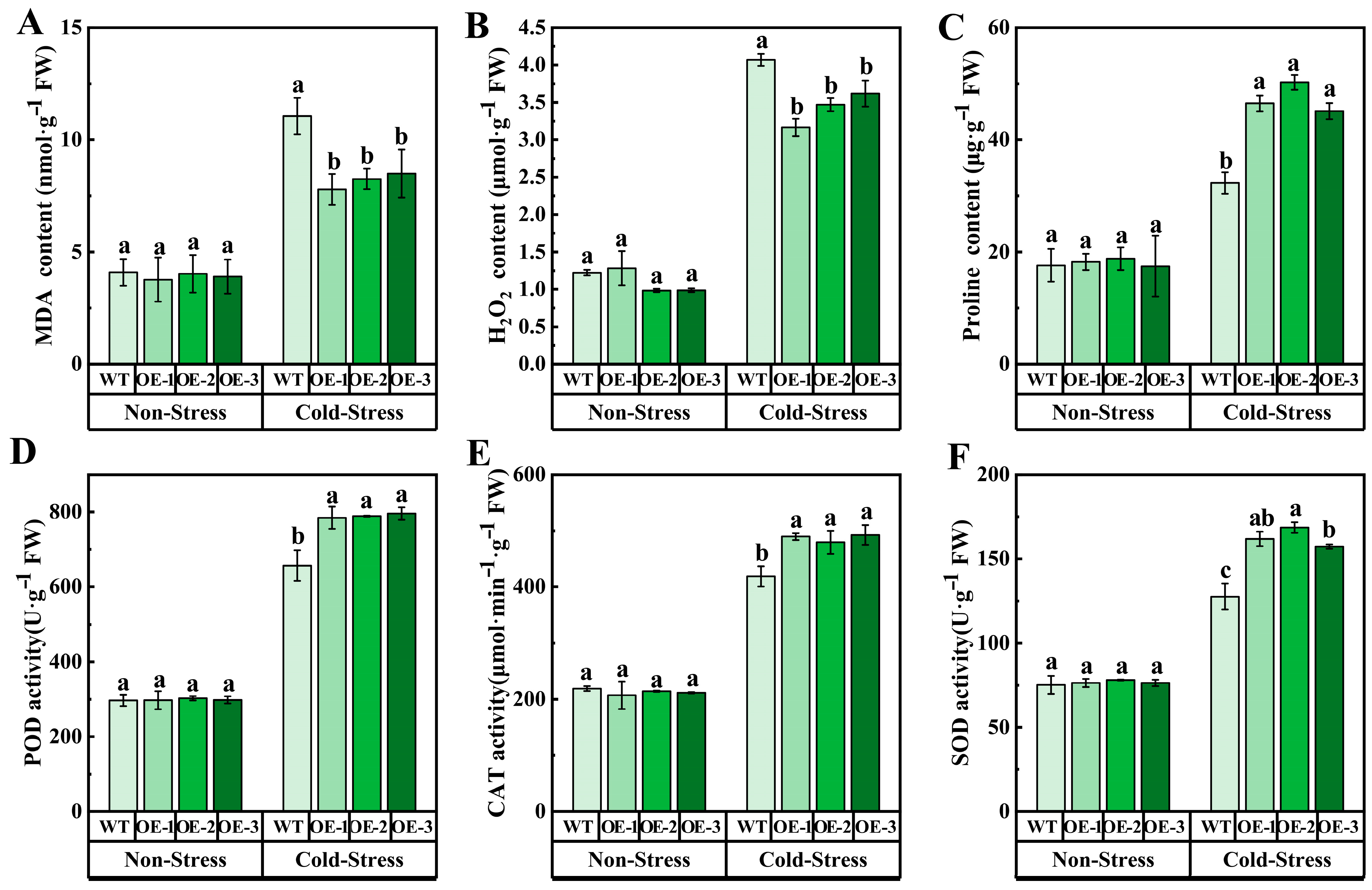
At the same time, under cold stress, the REL and MDA content in OE in OE were lower than those in WT, indicating that the damage to cell membrane caused by cold stress in transgenic Arabidopsis was reduced. In addition, compared with WT, the H2O2 content of transgenic Arabidopsis decreased under cold stress, while the activity of antioxidant enzymes (CAT, SOD, POD) and proline content increased, of which proline content increased by 31.58%, 32.70%, and 24.55%, respectively, compared with WT (Figure 6). To sum up, overexpression of VvJAZ13 increased the ability of scavenging reactive oxygen species (ROS), reduced the degree of membrane lipid peroxidation and cell dehydration, and increased the expression of cold response genes to protect Arabidopsis from cold stress.
2.6. Overexpression of VvJAZ13 Increased Cold Tolerance in Transgenic Grape Calli
After screening on the medium containing 10 mg·L−1 kanamycin, new growth of resistant calli was observed (Figure 7A). The new calli were transferred to the same screening medium again. After 30 days of culture, DNA was extracted and PCR was performed. The results showed that they were transformed successfully (OE-1, OE-2, OE-3) (Figure 7B). After cold stress at 4 °C for 10 days, it was found that OE grew faster than WT calli (Figure 7D). The results of qRT-PCR showed that compared with WT, the expression of VvJAZ13 was significantly up-regulated in transgenic calli under cold stress (Figure 7C).
In order to further study the mechanism of potential cold tolerance of VvJAZ13 overexpression, the transcriptional abundance of cold-related genes (VvCBF1, VvCBF2, VvCBF3, VvICE1a, VvICE1b, and VvICE1c) was determined. Under non-stress treatment, the expression levels of 6 genes were similar between WT and OE. However, under cold treatment, the expression of these genes in VvJAZ13 overexpression plants was significantly higher than that in WT plants, especially that in VvCBF2 was significantly higher than that in WT, which was 39.71%, and 34.63% higher than that in WT, respectively (Figure 8).
In order to study the effect of VvJAZ13 overexpression on grape calli under low temperature treatment, the physiological indexes of WT and OEs were further determined. The results showed that there was no significant difference in MDA and H2O2 contents between WT and OEs under non-stress, but the MDA and H2O2 contents of WT were significantly higher than those of OEs under cold stress. In order to alleviate the high level of ROS, the antioxidant mechanisms such as SOD, POD, and CAT were activated and the content of proline increased [35]. Under non-stress conditions, there was no significant difference in SOD, POD, CAT, activity and proline content between transgenic calli and WT, but the antioxidant enzyme activity and proline content of transgenic calli after cold treatment were higher than those of WT. The activity of POD increased by 19.45%, 20.12%, and 21.17%, and the content of proline increased by 43.90%, 55.54%, and 39.69% compared with WT (Figure 9). These results suggest that overexpression of VvJAZ13 enhances cold tolerance of grape calli by inhibiting severe cell membrane damage and activating antioxidant mechanism.
3. Discussion
Cold stress will adversely affect the growth and development of plants. Most temperate plants acquire cold tolerance through a process called cold adaptation. Cold stress is an important environmental restriction, which greatly affects plant growth and productivity and limits plant geographical distribution [1]. About 51% to 82% of crop yield losses are caused by extreme cold stress [36]. Under cold stress, plants will undergo a series of damage, including the destruction of cell membrane and cell structural stability, the decrease in photosynthesis and metabolism, and the excessive production of ROS [37,38,39].
In tea (Camellia sinensis), the CsJAZ1-1 fluorescence signal is only located in the nucleus, while CsJAZ1-2 and CsJAZ1-3 are widely distributed in the cytoplasm and nucleus. When these plasmids were transformed into Arabidopsis protoplasts, similar results were obtained. In short, the results showed that the subcellular localization of the three CsJAZ1 subtypes was determined by the N-terminal and Jas domain [40]. In this study, the transient transformation of tobacco leaves showed that VvJAZ13 was widely distributed in the cytoplasm and nucleus, which was the same as the mapping of CsJAZ1-2 and CsJAZ1-3 genes in tea.
JAZ proteins are a family widely involved in plant metabolism, defense and development [22]. Recently, the role of the JAZ protein family in the molecular mechanism related to the response to low temperature stress has been studied. For example, the interaction between JAZ1 and JAZ4 in Arabidopsis inhibits the transcription factors ICE1 and ICE2, thus inhibiting the ICE1-CBF/DREB1 signal pathway [16]. The JAZ signaling pathway leads to the release of bHLH and MYB TFs, which promote anthocyanin accumulation in Arabidopsis thaliana, thus improving its cold tolerance [41]. Plants have formed a complex regulatory mechanism against cold stress. In particular, the CBF cold response pathway plays a vital role in enhancing cold tolerance by directly activating multiple cold response genes [42,43]. In Arabidopsis, AtCBF1-AtCBF3 has been identified, and the target gene of CBF has been reported [44]. As a cold response gene, the CBF gene can be induced rapidly by low temperature. Chilling is mediated by a variety of transcription factors. For example, CBF expression inducer 1 (ICE1) can activate the expression of CBF, thereby enhancing the tolerance of plants to low temperature stress [45]. In our study, the relative expression levels of AtCBF1, AtCBF2, AtCBF3, and AtICE1 in transgenic Arabidopsis were up-regulated under cold stress. At the same time, the transcriptional abundance of VvCBF1, VvCBF2, VvCBF3, VvICE1a, VvICE1b, and VvICE1c genes in VvJAZ13 transgenic grape calli also increased. In addition, the expression of VvCBF and VvICE1 genes (VvCBF1, VvCBF2, VvCBF3) in VvJAZ13 transient overexpression transgenic grape leaves was also upregulated under cold stress. Therefore, we speculate that VvJAZ13 may respond to cold stress by directly regulating CBF signaling transduction in grape.
In kiwifruit (Actinidia arguta), the overexpression of AaBAM3.1 lines showed increased freezing tolerance with higher chlorophyll content and chlorophyll fluorescence [46]. We speculate that low temperature may inhibit the synthesis of chlorophyll. Overexpressed VvJAZ13 had a slightly lower sensitivity to inhibition, increased Fv/Fm and SPAD, and a higher efficiency of chlorophyll synthesis, which maintained the ability of the photosynthetic system, thus enhanced the cold tolerance of grape leaves.
The level of REL is considered to be the analysis of plant cell membrane damage under abiotic stress [47]. Meanwhile, MDA, as the final product of lipid peroxidation, is widely used as a marker of ROS-mediated plant damage [48]. In this study, VvJAZ13 overexpression significantly decreased REL in Arabidopsis and grape leaves. The content of MDA also decreased significantly in Arabidopsis and grape calli with VvJAZ13 overexpression. It is suggested that the overexpression of VvJAZ13 can prevent cell membrane damage caused by cold stress and respond to low temperature. Under low temperature stress, oxidative damage caused by excessive ROS accumulation is harmful to cell function and biological processes [45]. Under normal growth environment, there is a dynamic balance between ROS production and scavenging. However, when plants are subjected to cold stress, ROS production increases significantly, while antioxidant defense systems reduces the effects of oxidative stress on plants [49,50,51]. Proline accumulation is one of the important physiological responses of plants to resist low temperature stress, and plays an important role in plant cold resistance [52]. In our study, VvJAZ13 overexpression in Arabidopsis could significantly increase the activity and proline content of POD, SOD, CAT, and proline compared with WT under cold treatment. Similarly, under cold stress, we found that compared with WT calli, the overexpression of VvJAZ13 increased the activities of SOD, POD, CAT, and the content of proline. Therefore, we speculate that VvJAZ13 can reduce the accumulation of ROS by increasing the activity of antioxidant enzymes, so as to reduce cell membrane damage, reduce cell death, and enhance the cold resistance of grape. These results indicated that VvJAZ13 overexpression could improve the antioxidant capacity of Arabidopsis and grape, and thus enhance the cold resistance.
Plants have evolved complex regulatory mechanisms to cope with the effects of the external environment. The ubiquitin proteasome pathway involved in F-box is one of the key biological regulatory pathways, and a few studies have shown that F-box protein plays an important role in responding to abiotic stresses such as drought, salt, and temperature stress in plants. For example, the expression of F-box protein 7 in Arabidopsis was induced by low temperature stress, and the protein biosynthesis of fbp7 mutants was inhibited under low temperature stress [53]. The interaction between the F-box family gene ShPP2-1 (phloem protein 2-1) and ACR11A (ACT DOMAIN REPEAT) regulates the cold tolerance of tomato (Solanum lycopersicum) [54]. Overexpression of rice F-box protein MAIF1 (miRNAs regulated and abiotic stress induced F-box gene) reduces abiotic stress tolerance and promotes root growth, and may play a negative role in response to abiotic stress by regulating root growth [55]. TIFY proteins are a class of transcription factors unique to plants, characterized by highly conserved TIFY domains, which play important regulatory roles in plant growth and development and stress tolerance [56]. Overexpression of AtTIFY10a, 10b, and GsTIFY10a in Arabidopsis (as its wild soybean homolog) and OsJAZ8 in rice significantly regulated plant response to alkaline and salt stress, respectively [57,58]. The TIFY gene family has multiple regulatory roles in cell signal transduction and regulation of plant response to stress, so it may be a valuable stress response gene resource. The β-carotene hydroxylase gene bch1 can catalyze β-cryptoxanthin to synthesize zeaxanthin, which plays an important role in plant resistance to abiotic stress [59]. In this study, a yeast double hybrid showed that VvF-box, VvTIFY5A, VvTIFY9, and Vvbch1 had potential interaction with VvJAZ13 and played an important role in cold resistance of grape. However, we have not verified its interaction, and the interaction mechanism between grape and VvJAZ13 in cold resistance is not clear, and subsequent experiments are needed to study its molecular mechanism.
4. Materials and Methods
4.1. Test Material and Treatment
‘Pinot Noir’ grape plantlets were preserved in the plant tissue culture room of the Horticulture College of Gansu Agricultural University (Lanzhou, China). Tobacco (Nicotiana benthamiana) and Arabidopsis (ecotype Columbia) seeds were purchased from Nanjing Fengshuo Horticulture Ltd., Company (Nanjing, China). ‘Pinot Noir’ grape plantlets were subcultured in a GS solid medium (improved B5 solid medium) and cycled under 28 °C/25 °C and 12,000 lx light (LED plant lamp) for 16 h/dark for 8 h. After 35 days of culture, half of the samples were subjected to cold stress at 4 °C in a low-temperature incubator for 3 days.
‘Pinot Noir’ grape calli was preserved in a dark incubator at 28 °C. The culture conditions of Arabidopsis was 28 °C/25 °C, light for 16 h/dark for 8 h, and light intensity of 12,000 lx (LED plant lamp).
Tobacco was cultured in an incubator with 22 °C/18 °C, light intensity 10,000 lx (LED plant lamp), light for 16 h/dark for 8 h.
4.2. Chromosome Mapping and Evolutionary Analysis of VvJAZ13
The protein sequences of JAZ genes were obtained from the plant genome database phytozome v13.1: Home (https://phytozome-next.jgi.doe.gov/, accessed on 11 October 2023). Phylogenetic tree was constructed in MEGA 7.0 using the Neighbor Joining Algorithm (NJ) and set up with 1000 bootstrap repetitions [60]. The chromosome location was visualized by the software TBtools (version 1.108).
4.3. RNA Extraction and qRT-PCR
Total RNA was extracted from grape, Arabidopsis plant leaves, and grape calli by the RNAprep Pure Plant Kit (TIANGEN, Beijing, China). All primers were designed on an online website and synthesized by Sangon Biotech Co., Ltd. (Shanghai, China) (Table S1). The GAPDH gene (GenBank No. CB973647) was used as an internal control gene. Total reaction volume was 20 μL, including 7 μL ddH2O, 1 μL cDNA, 2 μL upstream and downstream primers, and 10 µL TaKaRa SYBR Premix Ex Taq. II (TaKaRa Biotechnology, Lanzhou, China). The cycling parameters were 95 °C for 30 s, 40 cycles of 95 °C for 5 s, and 60 °C for 34 s. Melting curve analysis was performed after the PCR cycle, with a program that included 95 °C for 15 s, 60 °C for 60 s, and 95 °C for 15 s [61].
4.4. Gene Cloning and Subcellular Location
The CDS of the VvJAZ13 (GSVIVG01015042001) gene was amplified by the homologous recombination technique. Primers were designed according to the CDS sequence in ‘Pinot Noir’(PN40024) 12Xv1 genome accession. Primers were synthesized by Sangon Biotech (Shanghai, China) Co., Ltd. (Table S1). Subcellular localisation of VvJAZ13 was performed with reference to Gou et al. [62]. The specific steps were as follows: the positive Agrobacterium containing the candidate gene was activated and cultured, and then expanded and centrifuged to collect the bacterial cells after activation, and then resuspended in a resuspension solution (10 m mol·L−1 MES, 10 mmol·L−1 MgCl2, 150 μmol·L−1 Acetosyringone) adjust to OD600 = 0.75, using the disposable syringe prepared in advance to accurately inject the bacteria liquid from the back of the prepared tobacco leaves, so that all the bacteria liquid could penetrate into the leaf flesh of the tobacco leaves. The injected tobacco was cultured in the dark for 24 h, and then GFP fluorescence was detected by a laser confocal scanning microscope (Olympus FV1000 Viewer, Tokyo, Japan) to observe the expression of VvJAZ13.
4.5. Instantaneous Transformation of Grape Leaves
The Agrobacterium tumefaciens solution was put into a 10 mL centrifuge tube and centrifuged for 5 min at 6000 rpm. The supernatant was removed and the heavy suspension was added to OD600 = 0.4–0.6. The suspension was activated for 3 h and infected grape leaves. The transient expression of grape leaves mediated by Agrobacterium tumefaciens refers to the method of Li et al. [63]. The transfected leaves were cultured at 25 °C for 24 h, then some of them were cultured at 4 °C and 16 h/8 h photoperiod for 24 h, and some of them were cultured at 25 °C and 16 h/8 h photoperiod for 24 h. The chlorophyll fluorescence imaging system (IMAPING-PAM, Walz, Rohrdorf, Germany) was used for visualization and determination of Fv/Fm. The SPAD value was determined by a chlorophyll content tester (SPAD-502Plus, Konica Minolta, Tokyo, Japan).
4.6. Arabidopsis Thaliana Transformation and Treatment
Arabidopsis was transformed by the Agrobacterium tumefaciens-mediated flower soaking method to obtain T0 generation transgenic plants [64], The T1 generations of positive transgenic seeds were selected on MS + 3% sucrose medium containing 50 mg·L−1 kanamycin, and the live seedlings were transplanted in nutritious soil to identify transgenic lines and harvest seeds. The same method was repeated to screen T2 and T3 generations. Finally, the T3 generation plants of three Arabidopsis lines (OE-1, OE-2 and OE-3) overexpressing VvJAZ13 were treated with cold stress.
Three stable T3 transgenic lines were screened from Arabidopsis. Some 3-week Arabidopsis plants with good growth and consistent growth, including transgenic plants (OEs) and wild type (WT) plants, were selected and placed in the plant low temperature incubator. It was cooled gradually from 24 °C to 4 °C, with a decrease of 1 °C per hour, and then cryogenically preserved for 48 h at 4 °C. This was followed by gradual cooling from 4 °C to −5 °C at a rate of 1 °C per hour, and samples were collected after 3 h at −5 °C. Then the contents of MDA, H2O2, proline and antioxidant (Peroxidase POD, Superoxide dismutase SOD, Catalase CAT) enzyme activity were determined. The transcriptional abundance of cold response genes AtCBF (AtCBF1, AtCBF2, AtCBF3) and AtICE1 were determined. It was then cooled gradually from −5 °C to −8 °C, with a decrease of 1 °C per hour. After 12 h of treatment at −8 °C, the phenotype was observed. After cold treatment, the survival of WT and OE plants was observed at room temperature for a week.
4.7. Grape Calli Transformation and Treatment
Calli was induced from the grape ovary of ‘Pinot Noir’ by basic medium NN69 (2.5 μmol·L−1 2,4-D + 2.5 μmol·L−1 NOA + 5 μmol·L−1 CPPU, culture in dark incubator at 25 °C). After successful induction, calli was subcultured on B5 medium (0.5 mg·L−1 NAA + 1 mg·L−1 KT).
The new grape calli was transferred to B5 solid medium and cultured for 10 days in a dark incubator at 25 °C to the calli to be infected. Agrobacterium solution was absorbed 200 μL and added to 50 mL LB liquid medium (containing 50 mg·L−1 Kanamycin and 50 mg·L−1 Rifampicin), cultured at 220 rpm at 28 °C to OD600 = 0.4–0.8. The bacterial liquid 5000 rpm was centrifuged for 10 min, then the supernatant was discarded and re-suspended in a 20 mL B5 liquid medium. The resuscitated bacterial liquid 5000 rpm was centrifuged for 10 min, and the supernatant was re-suspended in a B5 liquid medium (containing 50 mg·L−1 Acetosyringone). OD600 = 0.4 was adjusted and incubated at 28 °C for 3 h to obtain Agrobacterium tumefaciens suspension infection liquid.
The recombinant plasmids VvJAZ13 were transformed into ‘Pinot Noir’ calli following the Agrobacterium-mediated procedure according to the method proposed by Liang et al. [61]. The grape calli was transferred to a re-suspended Agrobacterium tumefaciens solution to shake for 8 min, then the calli was filtered with aseptic filter paper and cultured on a solid medium of 1/2 B5 + 1.5% sucrose for 2 days. After culture, the calli were transferred to a B5 solid selective medium (containing 10 mg·L−1 Kanamycin, 150 mg·L−1 Cefamycin) and cultured in the dark environment of 28 °C for 3–4 weeks. Then the screening medium was changed until new calli were formed, and then positive calli were identified at DNA level.
WT and transgenic grape calli were treated with cold stress at 4 °C for 10 days under dark conditions and then observed for phenotypic changes. After that, we determined the content of MDA, H2O2, and proline and the activity of antioxidant enzymes (POD, SOD, CAT). The transcriptional abundance of cold response genes VvCBF (VvCBF1, VvCBF2, VvCBF3) and VvICE1 (VvICE1a, VvICE1b, VvICE1c) were determined.
4.8. Y2H Protein Binding Assay
The CDS sequence of VvJAZ13 was cloned into pGBKT7 and fused to the DNA-binding domain to generate a bait construct. The primer sequences used are listed in supplementary Table S1. Y2H competent cells were prepared by a yeast peptone dextrose adenine (YPDA) liquid media. The recombinant plasmid was transformed into a Y2H Gold yeast strain and cultured on SD/-Trp medium at 30 °C for 3 days. The gene VvJAZ13 self-activation was detected. The potential interacting proteins were screened from the cold treatment-related AD library constructed and preserved by the Laboratory of Physiology and Biotechnology (Gansu Agricultural University of China). Screened potential protein sequences were cloned into pGADT7 and fused to activation domain sequences to generate prey construct. Yeast transformation and point-to-point verification of interaction proteins refer to the methods of Ren et al. [65].
4.9. Determination of Physiological Indexes of Arabidopsis and Grape Calli
The REL was measured using a model DDS307A (Leica Instruments Shanghai Co., Ltd., Shanghai, China). For subsequent data processing, refer to our previous methods [66]. H2O2, MDA, proline, and antioxidant enzyme activities (POD, SOD, CAT) were measured using a commercial ELISA kit purchased from Suzhou Comin Biotechnology Co., Ltd. (Suzhou, China) following the manufacturer’s instructions. Reaction mixtures were measured using a Multimode Reader (Spark® Multifunctional micro-plate detector, TECAN, Pudong, Shanghai, China).
4.10. Statistical Analysis
All data were determined in three independent biological replicates for each experiment. Data analysis is based on our previous method. Data analysis was performed by one-way analysis of variance (ANOVA) using SPSS 22.0 (SPSS, Inc., Chicago, IL, USA). Duncan’s multiple range test examined whether differences were significant (p < 0.05). The experimental data were processed by the 2−ΔΔCT method.
5. Conclusions
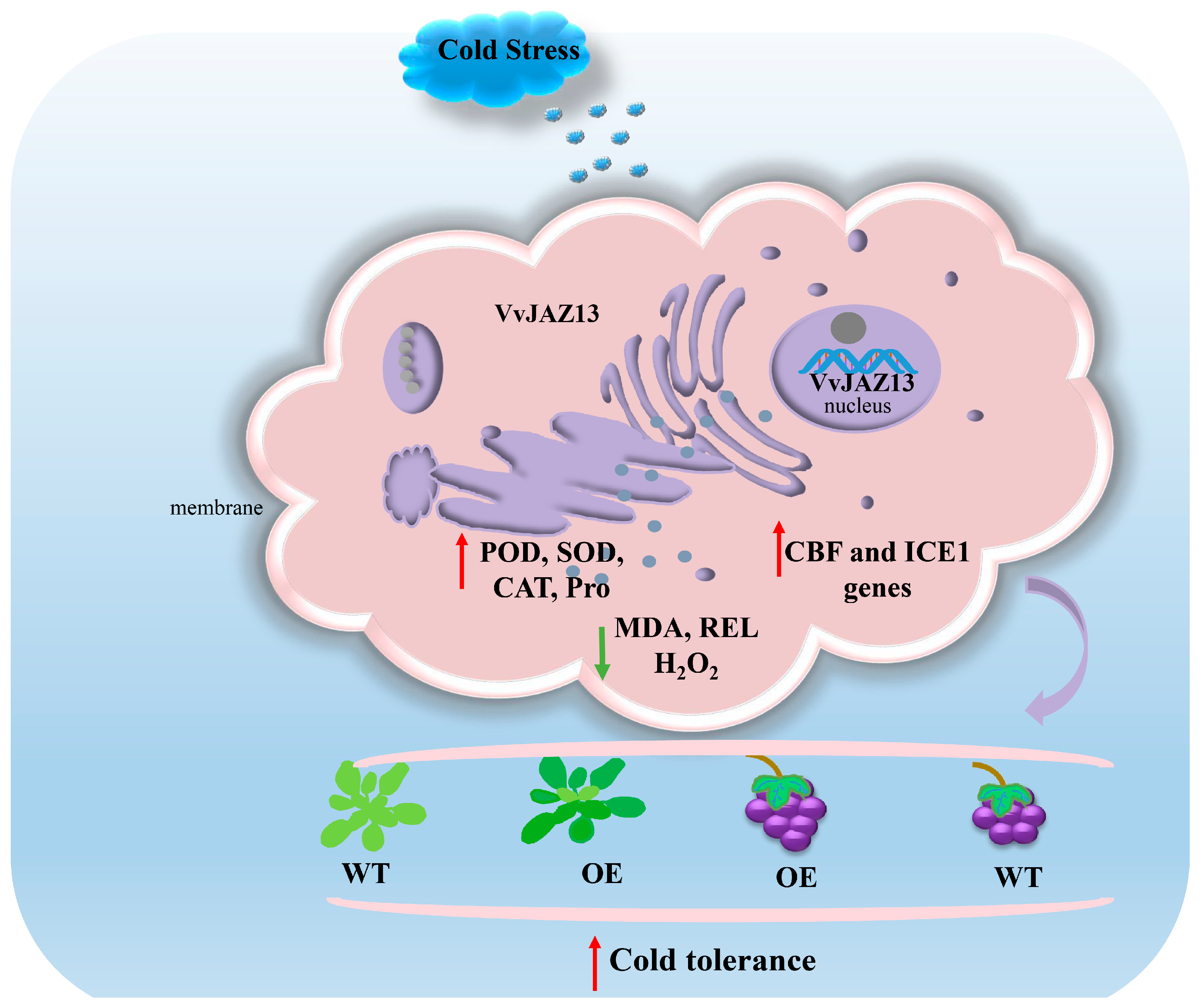
In conclusion, overexpression of VvJAZ13 can increase the antioxidant enzyme activity and proline content of Arabidopsis and grape, decrease REL and MDA content, and increase the transcriptional abundance of cold response genes CBF1-3 and ICE1, under cold stress, thereby improving the antioxidant capacity of transgenic plants and reducing membrane lipid peroxidation and accumulation of reactive oxygen species. Therefore, the overexpression of VvJAZ13 can improve the cold resistance of Arabidopsis and grape (Figure 10). This study laid a theoretical foundation for further understanding the biological function and molecular mechanism of VvJAZ13 under stress, and also provided a reference for further study of grape breeding under stress.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms25084458/s1.
Author Contributions
Conceived and designed the experiments: J.M.; Conducted experimentation and drafting: L.C., H.G. and M.L.; Analyzed the data: J.M. and L.C.; Wrote the paper: J.M. and L.C.; Revised the paper: J.M. and L.C.; Retrieve and data analysis: L.C. and S.L.; Revised the manuscript and technical input: J.M., L.G. and J.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Key Project of the Natural Science Foundation of Gansu Province (22JR5RA831) and the FuXi Foundation of the Gansu Agricultural University (No. Ganfx-03J02).
Data Availability Statement
Data is contained within the article and Supplementary Materials.
Acknowledgments
We thank Guobin Zhang (Gansu Agricultural University, China) for providing the chlorophyll fluorimeter.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant cold-stress responses. Trends Plant Sci. 2022, 27, 922–935. [Google Scholar] [CrossRef]
- Miura, K.; Furumoto, T. Cold Signaling and Cold Response in Plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef]
- Fennell, A. Freezing Tolerance and Injury in Grapevines. J. Crop Improv. 2004, 10, 201–235. [Google Scholar] [CrossRef]
- Carlow, C.E.; Faultless, J.T.; Lee, C.; Siddiqua, M.; Edge, A.; Nassuth, A. Nuclear localization and transactivation by Vitis CBF transcription factors are regulated by combinations of conserved amino acid domains. Plant Physiol. Bioch. 2017, 118, 306–319. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Xiao, H.; Siddiqua, M.; Braybrook, S.; Nassuth, A. Three grape CBF/DREB1 genes respond to low temperature, drought and abscisic acid. Plant Cell Environ. 2010, 29, 1410–1421. [Google Scholar] [CrossRef]
- Xiao, H.; Tattersall, E.A.R.; Siddiqua, M.K.; Cramer, G.R.; Nassuth, A. CBF4 is a unique member of the CBF transcription factor family of Vitis vinifera and Vitis riparia. Plant Cell Environ. 2008, 31, 1–10. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.H.; Hong, X.; Agarwal, M.; Zhu, J.K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef]
- Tillett, R.L.; Wheatley, M.D.; Tattersall, E.A.R.; Schlauch, K.A.; Cramer, G.R.; Cushman, J.C. The Vitis vinifera C-repeat binding protein 4 (VvCBF4) transcriptional factor enhances freezing tolerance in wine grape. Plant Biotechnol. J. 2012, 10, 105–124. [Google Scholar] [CrossRef]
- Siddiqua, M.; Nassuth, A. Vitis CBF1 and Vitis CBF4 differ in their effect on Arabidopsis abiotic stress tolerance, development and gene expression. Plant Cell Environ. 2011, 34, 1345–1359. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Zhu, W.; Wang, N.; Xin, H.P.; Li, S.H. Characterization of two VvICE1 genes isolated from ‘Muscat Hamburg’ grapevine and their effect on the tolerance to abiotic stresses. Sci. Hortic. 2014, 165, 266–273. [Google Scholar] [CrossRef]
- Sun, X.M.; Matus, J.T.; Wong, D.C.J.; Wang, Z.M.; Chai, F.M.; Zhang, L.L.; Fang, T.; Zhao, L.; Wang, Y.; Han, Y.P.; et al. The GARP/MYB-related grape transcription factor AQUILO improves cold tolerance and promotes the accumulation of raffinose family oligosaccharides. J. Exp. Bot. 2018, 69, 1749–1764. [Google Scholar] [CrossRef]
- Sun, X.M.; Zhang, L.L.; Wong, D.C.J.; Wang, Y.; Zhu, Z.F.; Xu, G.Z.; Wang, Q.F.; Li, S.H.; Liang, Z.C.; Xin, H.P. The ethylene response factor VaERF092 from Amur grape regulates the transcription factor VaWRKY33, improving cold tolerance. Plant J. 2019, 99, 988–1002. [Google Scholar] [CrossRef]
- Wang, M.Q.; Huang, Q.X.; Lin, P.; Zeng, Q.H.; Li, Y.; Liu, Q.L.; Zhang, L.; Pan, Y.Z.; Jiang, B.B.; Zhang, F. The overexpression of a transcription factor gene VbWRKY32 enhances the cold tolerance in Verbena bonariensis. Front. Plant Sci. 2020, 10, 1746. [Google Scholar] [CrossRef]
- Wang, Z.; Wong, D.C.J.; Wang, Y.X.; Guang, Z.; Ren, C.; Liu, Y.F.; Kuang, Y.F.; Fan, P.G.; Li, S.H.; Xin, H.P.; et al. GRAS-domain transcription factor PAT1 regulates jasmonic acid biosynthesis in grape cold stress response. Plant Physiol. 2021, 186, 1660–1678. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate regulates the inducer of CBF expression–c-repeat binding factor/DRE binding factor1 cascade and freezing tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, Y.; Han, X.; Wang, H.; Pan, J.; Yu, D. Jasmonate regulates leaf senescence and tolerance to cold stress: Crosstalk with other phytohormones. J. Exp. Bot. 2017, 68, 1361–1369. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signaling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Ghorbel, M.; Brini, F.; Sharma, A.; Landi, M. Role of jasmonic acid in plants: The molecular point of view. Plant Cell Rep. 2021, 40, 1471–1494. [Google Scholar] [CrossRef]
- Chung, H.S.; Cooke, T.F.; Depew, C.L.; Patel, L.C.; Ogawa, N.; Kobayashi, Y.; Howe, G.A. Alternative splicing expands the repertoire of dominant JAZ repressors of jasmonate signaling. Plant J. 2010, 63, 613–622. [Google Scholar] [CrossRef]
- Chung, H.S.; Howe, G.A. A critical role for the TIFY motif in repression of Jasmonate signaling by a stabilized splice variant of the JASMO-NATE ZIM-domain protein JAZ10 in Arabidopsis. Plant Cell 2009, 21, 131–145. [Google Scholar] [CrossRef]
- Howe, G.A.; Major, I.T.; Koo, A.J. Modularity in Jasmonate signaling for multistress resilience. Annu. Rev. Plant Biol. 2018, 69, 387–415. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosythesis, perception, signal tansduction and action in plant stress response, growth and development. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Song, S.; Qi, T.; Huang, H.; Ren, Q.; Wu, D.; Chang, C.; Peng, W.; Liu, Y.; Peng, J.; Xie, D. The Jasmonate-ZIM domain proteins interact with the R2R3-MYB transcription factors MYB21 and MYB24 to affect jasmonate-regulated stamen development in Arabidopsis. Plant Cell 2011, 23, 1000–1013. [Google Scholar] [CrossRef]
- Pascal, B.; Anne-Ruxandra, C.; Benoit, C.; Matija, D.; Joseph, R.E.; David, E.H.; Frederick, P.R.; Marc, V.; Mary, G.; Padmavathi, B.; et al. Evidence for Network Evolution in an Arabidopsis Interactome Map. Science 2011, 333, 601–607. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. The interplay between light and jasmonate signalling during defence and development. J. Exp. Bot. 2011, 62, 4087–4100. [Google Scholar] [CrossRef]
- Um, T.Y.; Lee, H.Y.; Lee, S.; Chang, S.H.; Chung, P.J.; Oh, K.B.; Kim, J.K.; Jang, G.; Choi, Y.D. Jasmonate Zim-Domain protein 9 interacts with slender rice 1 to mediate the antagonistic interaction between Jasmonic and Gibberellic Acid signals in rice. Front. Plant Sci. 2018, 9, 1866. [Google Scholar] [CrossRef]
- Jiang, Y.; Liang, G.; Yang, S.; Yu, D. Arabidopsis WRKY57 functions as a node of convergence for jasmonic acid- and auxin-mediated signaling in jasmonic acid-induced leaf senescence. Plant Cell 2014, 26, 230–245. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Z.; An, F.; Hao, D.; Li, P.; Song, J.; Yi, C.; Guo, H. Jasmonate-Activated MYC2 Represses ethylene insensitive3 activity to antagonize ethylene-promoted apical hook formation in Arabidopsis. Plant Cell 2014, 26, 1105–1117. [Google Scholar] [CrossRef]
- Zhu, D.; Cai, C.; Luo, X.; Bai, X.; Deyholos, M.K.; Chen, Q.; Chen, C.; Ji, W.; Zhu, Y.M. Over-expression of a novel JAZ family gene from Glycine soja, increases salt and alkali stress tolerance. Biochem. Biophys. Res. Commun. 2012, 426, 273–379. [Google Scholar] [CrossRef]
- Zhang, G.L.; Tang, W.L.; Kang, Q.; Shen, M.Y.; Xu, J.T.; Ping, D.D.; Hou, K.; Wu, W.; Xu, D.B. Identification and functional analysis of jasmonic acid signaling repressor McJAZ8 gene in Mentha canadensis. China J. Chin. Mater. Medica 2024, 49, 691–701. (In Chinese) [Google Scholar] [CrossRef]
- Shiduku, T.; Yumi, H.S.; Daisuke, T.; Shoko, Y.; Kazuya, A.; Kenji, G. Jasmonate induction of the monoterpene linalool confers resistance to rice bacterial blight and its biosynthesis is regulated by JAZ protein in rice. Plant Cell Environ. 2014, 37, 451–461. [Google Scholar] [CrossRef]
- Cai, X.; Zhen, J.B.; Jiang, Z.X.; Liu, L.L.; Liu, D.; Zhang, J.H.; Tan, H.Y.; Zhang, X.Y.; Chi, J. Cloning and Expression analysis of cold response gene GhJAZ1 from Gossypium hirsutum L. Acta Agric. Boreali-Sin. 2018, 33, 7–13. (In Chinese) [Google Scholar]
- Zhao, C.; Pan, X.; Yu, Y.; Zhu, Y.M.; Kong, F.J.; Sun, X.; Wang, F.F. Overexpression of a TIFY family gene, GsJAZ2, exhibits enhanced tolerance to alkaline stress in soybean. Mol. Breed. 2020, 40, 33. [Google Scholar] [CrossRef]
- He, X.Y.; Sambe, M.A.N.; Zhuo, C.L.; Tu, Q.H.; Guo, Z.F. A temperature induced lipocalin gene from Medicago falcata (MfTIL1) confers tolerance to cold and oxidative stress. Plant Mol. Biol. 2015, 87, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Hwarari, D.; Guan, Y.L.; Ahmad, B.; Movahedi, A.; Min, T.; Hao, Z.D.; Lu, Y.; Chen, J.H.; Yang, L.M. ICE-CBF-COR signaling cascade and its regulation in plants responding to cold stress. Int. J. Mol. Sci. 2022, 23, 1549. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Wang, X.F.; Zhang, X.W.; You, C.X.; Hao, Y.J. Apple B-box protein BBX37 regulates jasmonic acid mediated cold tolerance through the JAZ-BBX37-ICE1-CBF pathway and undergoes MIEL1-mediated ubiquitination and degradation. New Phytol. 2020, 229, 2707–2729. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Guo, Z. Effect of ABA and its biosynthesis inhibitor on chilling resistance and antioxidant enzymes activity. Acta Pratacult. Sin. 2005, 14, 94. [Google Scholar]
- Oquist, G.; Huner, N.P. Photosynthesis of overwintering evergreen plants. Annu. Rev Plant Biol. 2003, 54, 329–355. [Google Scholar] [CrossRef]
- Zhu, J.; Yan, X.; Liu, S.; Xia, X.B.; An, Y.L.; Xu, Q.S.; Zhao, S.Q.; Liu, L.; Guo, R.; Zhang, Z.L.; et al. Alternative splicing of CsJAZ1 negatively regulates flavan-3-ol biosynthesis in tea plants. Plant J. 2022, 110, 243–261. [Google Scholar] [CrossRef]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-Domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.R.; Zhang, N.B.; Jiao, Y.T.; Li, R.M.; Xiao, D.M.; Wang, Z.P. The grapevine basic helix-loop-helix (bHLH) transcription factor positively modulates CBF-pathway and confers tolerance to cold-stress in Arabidopsis. Mol. Biol. Rep. 2014, 41, 5329–5342. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.T.; Ding, Y.L.; Yang, S.H. Molecular regulation of CBF signaling in cold acclimation. Trends Plant Sci. 2018, 23, 623–637. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, S.J.; Zarka, D.G.; Stockinger, E.J.; Salazar, M.P.; Houghton, J.M.; Thomashow, M.F. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J. 1998, 16, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Kidokoro, S.; Yoneda, K.; Takasaki, H.; Takahashi, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Different cold-signaling pathways function in the responses to rapid and gradual decreases in temperature. Plant Cell 2017, 29, 760–774. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.H.; Hu, C.G.; Qi, X.J.; Chen, J.Y.; Zhong, Y.P.; Muhammad, A.; Lin, M.M.; Fang, J.B. The AaCBF4-AaBAM3.1 module enhances freezing tolerance of kiwifruit (Actinidia arguta). Hortic. Res. 2021, 8, 97. [Google Scholar] [CrossRef] [PubMed]
- Baiji, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Lv, Y.; Yang, M.; Hu, D.; Yang, Z.; Ma, S.; Li, X.; Xiong, L. The OsMYB30 transcription factor suppresses cold tolerance by interacting with a JAZ protein and suppressing β-amylase expression. Plant Physiol. 2017, 173, 1475–1491. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.Y.; Qing, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sucu, S.; Yagci, A.; Yildirim, K. Changes in morphological, physiological traits and enzyme activity of grafted and ungrafted grapevine rootstocks under drought stress. Erwerbs-Obstbau 2018, 60, 127–136. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Villalobos, L.I.A.; Nill, C.; Marrocco, K.; Schwechheimer, C. The evolutionarily conserved Arabidopsis thaliana F-box protein AtFBP7 is required for efficient translation during temperature stress. Gene 2007, 392, 106–116. [Google Scholar] [CrossRef]
- Song, J.W.; Shang, L.L.; Chen, S.W.; Lu, Y.G.; Zhang, Y.Y.; Yang, B.O.; Ye, Z.B.; Zhang, J.H. Interactions between ShPP2-1, an F-box family gene, and ACR11A regulate cold tolerance of tomato. Hortic. Res. 2021, 8, 148. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.S.; Chen, X.Y.; Yang, K.; Sun, Z.X.; Fu, Y.P.; Zhang, Y.M.; Fang, R.X. Overexpression of an F-box protein gene reduces abiotic stress tolerance and promotes root growth in rice. Mol. Plant 2011, 4, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, B.; Grunewald, W.; Bateman, A.; Kohchi, T.; Gheysen, G. The tify family previously known as ZIM. Trends Plant Sci. 2007, 12, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Li, R.; Liu, X.; Sun, M.; Wu, J.; Zhang, N.; Zhu, Y.M.; Wu, K.Q. The positive regulatory roles of the TIFY10 proteins in plant responses to alkaline stress. PLoS ONE 2014, 9, e111984. [Google Scholar] [CrossRef]
- Peethambaran, P.K.; Glenz, R.; Höninger, S.; Islam, S.M.S.; Hummel, S.; Harter, K.; Kolukisaoglu, Ü.; Meynard, D.; Guiderdoni, E.; Nick, P.; et al. Salt-inducible expression of OsJAZ8 improves resilience against salt-stress. BMC Plant Biol. 2018, 18, 311. [Google Scholar] [CrossRef] [PubMed]
- Arango, J.; Jourdan, M.; Geoffriau, E.; Beyer, P.; Welsch, R. Carotene hydroxylase activity determines the levels of both α-Carotene and total carotenoids in orange carrots. Plant Cell 2014, 26, 2223–2233. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGAX: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Liang, G.P.; Hou, Y.; Wang, H.; Wang, P.; Mao, J.; Hong, B. VaBAM1 weakens cold tolerance by interacting with the negative regulator VaSR1 to suppress β-amylase expression. Int. J. Biol. Macromol. 2023, 225, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- Gou, H.M.; Nai, G.; Lu, S.X.; Hong, B.H.; Mao, J. Genome-wide identification and expression analysis of PIN gene family under phytohormone and abiotic stresses in Vitis vinifera L. Physiol. Mol. Biol. Plants 2022, 28, 1905–1919. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, X.; Wang, X.F.; Xi, Z.M. An AP2/ERF transcription factor VvERF63 positively regulates cold tolerance in Arabidopsis and grape leaves. Environ. Exp. Bot. 2023, 205, 105124. [Google Scholar] [CrossRef]
- Lu, S.X.; Wang, P.; Nai, G.; Li, Y.M.; Su, Y.L.; Liang, G.P.; Chen, B.H.; Mao, J. Insight into VvGH3 genes evolutional relationship from monocotyledons and dicotyledons reveals that VvGH3-9 negatively regulates the drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2022, 172, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.X.; Zhang, P.; Dai, Y.; Liu, X.; Lu, S.X.; Guo, L.L.; Gou, H.M.; Mao, J. Evolution of the 14-3-3 gene family in monocotyledons and dicotyledons and validation of MdGRF13 function in transgenic Arabidopsis thaliana. Plant Cell Rep. 2023, 42, 1345–1364. [Google Scholar] [CrossRef]
- Ma, X.; Chen, C.; Yang, M.; Dong, X.; Lv, W.; Meng, Q. Cold-regulated protein (SlCOR413IM1) confers chilling stress tolerance in tomato plants. Plant Physiol. Biochem. 2018, 124, 29–39. [Google Scholar] [CrossRef]
Figure 1.
Evolutionary analysis of the VvJAZ13 gene in grape.

Figure 2.
Subcellular localization of VvJAZ13 protein. The GFP fluorescence image, Chloroplast auto fluorescence (CHI) image, bright field (Bright) image, and the merged GFP, CHI, and Bright images are shown. Scare bar: 30 μm.
Figure 2.
Subcellular localization of VvJAZ13 protein. The GFP fluorescence image, Chloroplast auto fluorescence (CHI) image, bright field (Bright) image, and the merged GFP, CHI, and Bright images are shown. Scare bar: 30 μm.

Figure 3.
Verification of interaction between VvJAZ13 protein and interaction protein in the yeast two-hybrid system. AD and BD represent pGBKT7 and pGADT7, respectively. DDO indicates SD/-Trp, TDO indicates SD/-Trp/-Leu/-His, QDO indicates SD/-Trp/-Leu/-His/-Ade, X indicates X-α-gal.
Figure 3.
Verification of interaction between VvJAZ13 protein and interaction protein in the yeast two-hybrid system. AD and BD represent pGBKT7 and pGADT7, respectively. DDO indicates SD/-Trp, TDO indicates SD/-Trp/-Leu/-His, QDO indicates SD/-Trp/-Leu/-His/-Ade, X indicates X-α-gal.

Figure 4.
Transient transformation of VvJAZ13 in grape leaves. (A) Fluorescence intensity of WT and VvJAZ13-OEs grape leaves, (B) Expression of VvJAZ13 in WT and VvJAZ13-OEs grape leaves, (C–E) REL, MDA, and proline contents of WT and VvJAZ13-OEs grape leaves (F) Fv/Fm of WT and VvJAZ13-OEs grape leaves, (G) SPAD value of WT and VvJAZ13-OEs grape leaves. Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).
Figure 4.
Transient transformation of VvJAZ13 in grape leaves. (A) Fluorescence intensity of WT and VvJAZ13-OEs grape leaves, (B) Expression of VvJAZ13 in WT and VvJAZ13-OEs grape leaves, (C–E) REL, MDA, and proline contents of WT and VvJAZ13-OEs grape leaves (F) Fv/Fm of WT and VvJAZ13-OEs grape leaves, (G) SPAD value of WT and VvJAZ13-OEs grape leaves. Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).

Figure 5.
Phenotypic aspect and gene expression levels of VvJAZ13 in Arabidopsis transgenic lines under cold stress. (A) Phenotypes of WT and VvJAZ13-OEs Arabidopsis, (B) Expression levels of the VvJAZ13 gene in WT and VvJAZ13-OEs Arabidopsis, (C) Expression levels of the cold response genes AtCBF1, AtCBF2, AtCBF3, and AtICE1 in WT and VvJAZ13-OEs Arabidopsis. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).
Figure 5.
Phenotypic aspect and gene expression levels of VvJAZ13 in Arabidopsis transgenic lines under cold stress. (A) Phenotypes of WT and VvJAZ13-OEs Arabidopsis, (B) Expression levels of the VvJAZ13 gene in WT and VvJAZ13-OEs Arabidopsis, (C) Expression levels of the cold response genes AtCBF1, AtCBF2, AtCBF3, and AtICE1 in WT and VvJAZ13-OEs Arabidopsis. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).

Figure 6.
Analysis of membrane lipid peroxidation and antioxidant enzyme activities of Arabidopsis transgenic lines of VvJAZ13 under cold stress. (A) REL of WT and OEs Arabidopsis, (B) MDA content of WT and OEs Arabidopsis, (C) H2O2 content of WT and OEs Arabidopsis, (D–F) POD, SOD, and CAT activities of WT and OEs Arabidopsis, (G) Proline content of WT and OEs Arabidopsis. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).
Figure 6.
Analysis of membrane lipid peroxidation and antioxidant enzyme activities of Arabidopsis transgenic lines of VvJAZ13 under cold stress. (A) REL of WT and OEs Arabidopsis, (B) MDA content of WT and OEs Arabidopsis, (C) H2O2 content of WT and OEs Arabidopsis, (D–F) POD, SOD, and CAT activities of WT and OEs Arabidopsis, (G) Proline content of WT and OEs Arabidopsis. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).

Figure 7.
VvJAZ13 identification of transgenic grape calli. (A) Screening of transgenic calli in a B5 medium containing Kana resistance, (B) Positive detection of grape calli, (C) Expression levels of the VvJAZ13 gene in WT and VvJAZ13-OEs grape calli, (D) Phenotypes of WT and VvJAZ13-OEs grape calli. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).
Figure 7.
VvJAZ13 identification of transgenic grape calli. (A) Screening of transgenic calli in a B5 medium containing Kana resistance, (B) Positive detection of grape calli, (C) Expression levels of the VvJAZ13 gene in WT and VvJAZ13-OEs grape calli, (D) Phenotypes of WT and VvJAZ13-OEs grape calli. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).

Figure 8.
Expression levels of VvCBF1, VvCBF2, VvCBF3, VvICE1a, VvICE1b, and VvICE1c in WT and VvPAL10-OEs grape calli. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).
Figure 8.
Expression levels of VvCBF1, VvCBF2, VvCBF3, VvICE1a, VvICE1b, and VvICE1c in WT and VvPAL10-OEs grape calli. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).

Figure 9.
Analysis of membrane lipid peroxidation and antioxidant enzyme activities of grape calli transgenic lines of VvJAZ13. (A) The content of MDA in WT and VvPAL10-OEs grape calli, (B) The content of H2O2 in WT and VvPAL10-OEs grape calli, (C) The content of Proline in WT and VvPAL10-OEs grape calli, (D–F) POD, SOD, and CAT enzymes activities were determined in WT and VvPAL10-OEs grape calli. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).
Figure 9.
Analysis of membrane lipid peroxidation and antioxidant enzyme activities of grape calli transgenic lines of VvJAZ13. (A) The content of MDA in WT and VvPAL10-OEs grape calli, (B) The content of H2O2 in WT and VvPAL10-OEs grape calli, (C) The content of Proline in WT and VvPAL10-OEs grape calli, (D–F) POD, SOD, and CAT enzymes activities were determined in WT and VvPAL10-OEs grape calli. The bar represents the value of the standard error (three biological replicates). Based on Duncan’s multiple range test, lowercase letters represent significant differences (p < 0.05).

Figure 10.
Systematic model of overexpression of VvJAZ13 gene to improve cold tolerance of Arabidopsis and grape. Green and red arrows represent down and up-regulated expression levels in WT vs OE.
Figure 10.
Systematic model of overexpression of VvJAZ13 gene to improve cold tolerance of Arabidopsis and grape. Green and red arrows represent down and up-regulated expression levels in WT vs OE.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Che, L.; Lu, S.; Gou, H.; Li, M.; Guo, L.; Yang, J.; Mao, J. VvJAZ13 Positively Regulates Cold Tolerance in Arabidopsis and Grape. Int. J. Mol. Sci. 2024, 25, 4458. https://doi.org/10.3390/ijms25084458
AMA Style
Che L, Lu S, Gou H, Li M, Guo L, Yang J, Mao J. VvJAZ13 Positively Regulates Cold Tolerance in Arabidopsis and Grape. International Journal of Molecular Sciences. 2024; 25(8):4458. https://doi.org/10.3390/ijms25084458
Chicago/Turabian StyleChe, Lili, Shixiong Lu, Huimin Gou, Min Li, Lili Guo, Juanbo Yang, and Juan Mao. 2024. "VvJAZ13 Positively Regulates Cold Tolerance in Arabidopsis and Grape" International Journal of Molecular Sciences 25, no. 8: 4458. https://doi.org/10.3390/ijms25084458
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.