
Research Progress on Anti-Inflammatory Effects and Related Mechanisms of Astragalin

Abstract
:
1. Introduction
2. Materials and Methods
3. The In Vitro Study on the Anti-Inflammatory Effect of Astragalin
3.1. The Effect of Astragalin on Macrophage Inflammation-Related Factors and Protein Levels
3.2. The Effect of Astragalin on Inflammation-Related Factors and Protein Levels in Microglia
3.3. The Effect of Astragalin on Inflammation-Related Factors and Protein Levels in Epithelial Cells
3.4. The Effect of Astragalin on Inflammation-Related Factors and Protein Levels in Other Types of Cells
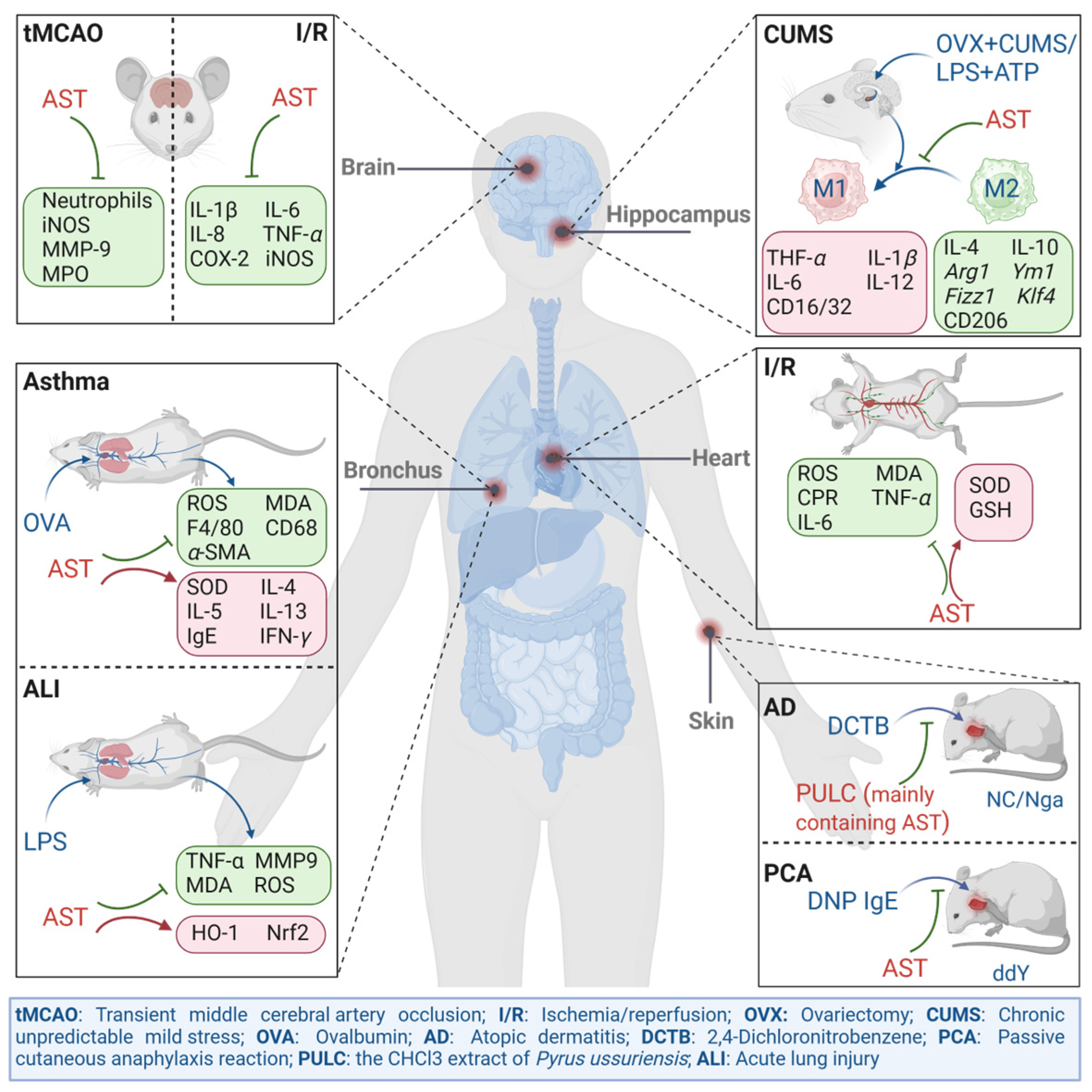
4. The In Vivo Alleviating Effect of Astragalin on Inflammatory Disease Models
4.1. The Effect of Astragalin on Carrageenan-Induced Mouse Foot Swelling Model
4.2. The Effect of Astragalin on the Atopic Dermatitis Model
4.3. The Effect of Astragalin on the Respiratory Inflammation Model
4.4. The Effect of Astragalin on the Ischemia–Reperfusion Injury Model
4.5. The Effect of Astragalin on the Depression Model
5. The Regulatory Effect of Astragalin on Inflammation-Related Pathways
5.1. Astragalin Exerts Anti-Inflammatory Effects by Affecting the NF-κB Signaling Pathway
5.1.1. Astragalin Exerts Anti-Inflammatory Effects by Affecting the TLR4/NF-κB Signaling Pathway
5.1.2. Astragalin Exerts Anti-Inflammatory Effects by Affecting the Notch/HES-1-NF-κB Signaling Pathway
5.1.3. Astragalin Exerts Anti-Inflammatory Effects by Affecting the STAT3/NF-κB Signaling Pathway
5.1.4. Astragalin Exerts Anti-Inflammatory Effects by Affecting the Nrf2/HO-1/NF-κB Signaling Pathway
5.1.5. Astragalin Exerts Anti-Inflammatory Effects by Affecting the NOX2/ROS/NF-κB Signaling Pathway
5.1.6. Astragalin Exerts Anti-Inflammatory Effects by Affecting the SIRT1/NF-κB Signaling Pathway
5.2. Astragalin Exerts Anti-Inflammatory Effects by Affecting the MAPK Signaling Pathway
5.3. Astragalin Exerts Anti-Inflammatory Effects by Affecting the IL-4R/JAK1/STAT6 Signaling Pathway
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviation
References
- Yeung, Y.T.; Aziz, F.; Guerrero-Castilla, A.; Arguelles, S. Signaling pathways in inflammation and anti-inflammatory therapies. Curr. Pharm. Des. 2018, 24, 1449–1484. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.G.; Beart, P.M. Inflammation: Maladies, models, mechanisms and molecules. Br. J. Pharmacol. 2016, 173, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive lipids, inflammation and chronic diseases. Adv. Drug Deliv. Rev. 2020, 159, 133–169. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxid. Med. Cell Longev. 2016, 2016, e5276130. [Google Scholar] [CrossRef] [PubMed]
- Oray, M.; Abu Samra, K.; Ebrahimiadib, N.; Meese, H.; Foster, C.S. Long-term side effects of glucocorticoids. Expert. Opin. Drug Saf. 2016, 15, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Parolini, M. Toxicity of the Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) acetylsalicylic acid, paracetamol, diclofenac, ibuprofen and naproxen towards freshwater invertebrates: A review. Sci. Total Environ. 2020, 740, 140043. [Google Scholar] [CrossRef] [PubMed]
- García-Rayado, G.; Navarro, M.; Lanas, A. NSAID induced gastrointestinal damage and designing GI-sparing NSAIDs. Expert. Rev. Clin. Pharmacol. 2018, 11, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a dietary anti-inflammatory agent: Current therapeutic standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, e125124. [Google Scholar] [CrossRef]
- Hussain, T.; Murtaza, G.; Yang, H.; Kalhoro, M.S.; Kalhoro, D.H. Exploiting anti-inflammation effects of flavonoids in chronic inflammatory diseases. Curr. Pharm. Des. 2020, 26, 2610–2619. [Google Scholar] [CrossRef]
- Czerwińska, M.E.; Świerczewska, A.; Granica, S. Bioactive constituents of Lamium album L. as inhibitors of cytokine secretion in human neutrophils. Molecules 2018, 23, 2770. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liu, D.; Liu, N.; Kuang, Y.; Tai, Q. Astragalin reduces lipopolysaccharide-induced acute lung injury in rats via induction of heme oxygenase-1. Arch. Pharm. Res. 2019, 42, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lv, J.; Li, X.; Lin, Q. The flavonoid astragalin shows anti-tumor activity and inhibits PI3K/AKT signaling in gastric cancer. Chem. Biol. Drug Des. 2021, 98, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Z.; Wang, S.H.; Zhang, R.H.; Lin, J.H.; Tian, Y.H.; Yang, Y.Q.; Liu, J.; Ma, Y.X. Neuroprotective effect of astragalin via activating PI3K/Akt-mTOR-mediated autophagy on APP/PS1 mice. Cell Death Discov. 2023, 9, e15. [Google Scholar] [CrossRef]
- Liu, L.; Wang, D.; Qin, Y.; Xu, M.; Zhou, L.; Xu, W.; Liu, X.; Ye, L.; Yue, S.; Zheng, Q.; et al. Astragalin promotes osteoblastic differentiation in MC3T3-E1 cells and bone formation in vivo. Front. Endocrinol. 2019, 10, e228. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, M.; Jiang, S.; Liang, Z.; Wang, J.; Liu, Z.; Wang, H.D.; Kang, W. Evaluation procoagulant activity and mechanism of astragalin. Molecules 2020, 25, 177. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Ma, Z.; Wang, D.; Niu, Y. Ultrasound-assisted extraction, optimization, isolation, and antioxidant activity analysis of flavonoids from Astragalus membranaceus stems and leaves. Ultrason. Sonochem. 2022, 90, e106190. [Google Scholar] [CrossRef]
- Adesso, S.; Russo, R.; Quaroni, A.; Autore, G.; Marzocco, S. Astragalus membranaceus extract attenuates inflammation and oxidative stress in intestinal epithelial cells via NF-κB activation and Nrf2 response. Int. J. Mol. Sci. 2018, 19, 800. [Google Scholar] [CrossRef]
- Doi, K.; Kojima, T.; Makino, M.; Kimura, Y.; Fujimoto, Y. Studies on the constituents of the leaves of Morus alba L. Chem. Pharm. Bull. 2001, 49, 151–153. [Google Scholar] [CrossRef]
- Oliveira, A.M.; Nascimento, M.F.; Ferreira, M.R.; Moura, D.F.; Souza, T.G.; Silva, G.C.; Ramos, E.H.; Paiva, P.M.; Medeiros, P.L.; Silva, T.G.; et al. Evaluation of acute toxicity, genotoxicity and inhibitory effect on acute inflammation of an ethanol extract of Morus alba L. (Moraceae) in mice. J. Ethnopharmacol. 2016, 194, 162–168. [Google Scholar] [CrossRef]
- He, X.; Yang, W.; Ye, M.; Wang, Q.; Guo, D. Differentiation of Cuscuta chinensis and Cuscuta australis by HPLC-DAD-MS analysis and HPLC-UV quantitation. Planta Med. 2011, 77, 1950–1957. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Yang, H.; Jung, D.H.; Hwang, J.T.; Ko, B.S. Ameliorating effects of Cuscuta chinensis Lamak extract on hind-limb ischemia, and angiogenic or inflammatory associated factors in ovariectomized mice. Mol. Med. Rep. 2019, 19, 3321–3329. [Google Scholar] [CrossRef] [PubMed]
- Nhiem, N.X.; Tai, B.H.; Quang, T.H.; Kiem, P.V.; Minh, C.V.; Nam, N.H.; Kim, J.H.; Im, L.R.; Lee, Y.M.; Kim, Y.H. A new ursane-type triterpenoid glycoside from Centella asiatica leaves modulates the production of nitric oxide and secretion of TNF-α in activated RAW 264.7 cells. Bioorg. Med. Chem. Lett. 2011, 21, 1777–1781. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Choi, H.K.; N’deh, K.P.U.; Choi, Y.J.; Fan, M.; Kim, E.K.; Chung, K.H.; An, A.J.H. Inhibitory effect of Centella asiatica extract on DNCB-induced atopic dermatitis in HaCaT cells and BALB/c mice. Nutrients 2020, 12, 411. [Google Scholar] [CrossRef] [PubMed]
- Hering, A.; Stefanowicz-Hajduk, J.; Hałasa, R.; Olech, M.; Nowak, R.; Kosiński, P.; Ochocka, J.R. Polyphenolic characterization, antioxidant, antihyaluronidase and antimicrobial activity of young leaves and stem extracts from Rubus caesius L. Molecules 2022, 27, 6181. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Lee, M.S.; Chang, E.; Kim, C.T.; Kim, Y. Mulberry (Morus alba L.) fruit extract ameliorates inflammation via regulating microRNA-21/132/143 expression and increases the skeletal muscle mitochondrial content and AMPK/SIRT activities. Antioxidants 2021, 10, 1453. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Doh, E.J.; Lee, G. Quantitative comparison of the marker compounds in different medicinal parts of Morus alba L. using high-performance liquid chromatography-diode array detector with chemometric analysis. Molecules 2020, 25, 5592. [Google Scholar] [CrossRef]
- Riaz, A.; Rasul, A.; Hussain, G.; Zahoor, M.K.; Jabeen, F.; Subhani, Z.; Younis, T.; Ali, M.; Sarfraz, I.; Selamoglu, Z. Astragalin: A bioactive phytochemical with potential therapeutic activities. Adv. Pharmacol. Sci. 2018, 2018, e9794625. [Google Scholar] [CrossRef]
- Chen, J.; Zhong, K.; Qin, S.; Jing, Y.; Liu, S.; Li, D.; Peng, C. Astragalin: A food-origin flavonoid with therapeutic effect for multiple diseases. Front. Pharmacol. 2023, 14, e1265960. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, M.; Wang, L.; Chen, W. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, e173090. [Google Scholar] [CrossRef]
- Lee, H.B.; Kim, E.K.; Park, S.J.; Bang, S.G.; Kim, T.G.; Chung, D.W. Isolation and anti-inflammatory effect of astragalin synthesized by enzymatic hydrolysis of tea seed extract. J. Sci. Food Agric. 2011, 91, 2315–2321. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Nam, Y.H.; Jung, Y.S.; Oh, S.M.; Nguyen, T.N.; Lee, M.H.; Kim, D.O.; Kang, T.H.; Lee, D.Y.; Baek, N.I. Aurones and flavonols from Coreopsis lanceolata L. flowers and their anti-oxidant, pro-inflammatory inhibition effects, and recovery effects on alloxan-induced pancreatic islets in zebrafish. Molecules 2021, 26, 6098. [Google Scholar] [CrossRef]
- Rao, Y.K.; Fang, S.H.; Tzeng, Y.M. Antiinflammatory activities of flavonoids and a triterpene caffeate isolated from Bauhinia variegate. Phytother. Res. 2008, 22, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, V.; Gupta, S.; Sharma, N.; Tiwari, H.; Bhardwaj, S.; Dutt, P.; Satti, N.; Nargotra, A.; Bhagat, A.; Ahmed, Z. Kaempferol-3-O-β-D-glucuronate exhibit potential anti-inflammatory effect in LPS stimulated RAW 264.7 cells and mice model. Int. Immunopharmacol. 2018, 57, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, S.H. Inhibitory effect of astragalin on expression of lipopolysaccharide-induced inflammatory mediators through NF-κB in macrophages. Arch. Pharm. Res. 2011, 34, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Atay Balkan, İ.; Gören, A.C.; Kırmızıbekmez, H.; Yeşilada, E. Evaluation of the in vitro anti-inflammatory activity of Nerium oleander L. flower extracts and activity-guided isolation of the active constituents. Rec. Nat. Prod. 2018, 12, 128–141. [Google Scholar] [CrossRef]
- Borst, K.; Dumas, A.A.; Prinz, M. Microglia: Immune and non-immune functions. Immunity 2021, 54, 2194–2208. [Google Scholar] [CrossRef]
- Kim, E.H.; Shim, Y.Y.; Lee, H.I.; Lee, S.; Reaney, M.J.T.; Chung, M.J. Astragalin and isoquercitrin isolated from Aster scaber suppress LPS-induced neuroinflammatory responses in microglia and mice. Foods 2022, 11, 1505. [Google Scholar] [CrossRef]
- Lim, H.J.; Prajapati, R.; Seong, S.H.; Jung, H.A.; Choi, J.S. Antioxidant and antineuroinflammatory mechanisms of kaempferol-3-O-β-D-glucuronate on lipopolysaccharide-stimulated BV2 microglial cells through the Nrf2/HO-1 signaling cascade and MAPK/NF-κB pathway. ACS Omega 2023, 8, 6538–6549. [Google Scholar] [CrossRef]
- Reddel, R.R.; Ke, Y.; Gerwin, B.I.; McMenamin, M.G.; Lechner, J.F.; Su, R.T.; Brash, D.E.; Park, J.B.; Rhim, J.S.; Harris, C.C. Transformation of human bronchial epithelial cells by infection with SV40 or adenovirus-12 SV40 hybrid virus, or transfection via strontium phosphate coprecipitation with a plasmid containing SV40 early region genes. Cancer Res. 1988, 48, 1904–1909. [Google Scholar]
- Albright, C.D.; Jones, R.T.; Hudson, E.A.; Fontana, J.A.; Trump, B.F.; Resau, J.H. Transformed human bronchial epithelial cells (BEAS-2B) alter the growth and morphology of normal human bronchial epithelial cells in vitro. Cell Biol. Toxicol. 1990, 6, 379–398. [Google Scholar] [CrossRef]
- Kim, Y.H.; Choi, Y.J.; Kang, M.K.; Park, S.H.; Antika, L.D.; Lee, E.J.; Kim, D.Y.; Kang, Y.H. Astragalin inhibits allergic inflammation and airway thickening in ovalbumin-challenged mice. J. Agric. Food Chem. 2017, 65, 836–845. [Google Scholar] [CrossRef]
- Cho, I.H.; Gong, J.H.; Kang, M.K.; Lee, E.J.; Park, J.H.; Park, S.J.; Kang, Y.H. Astragalin inhibits airway eotaxin-1 induction and epithelial apoptosis through modulating oxidative stress-responsive MAPK signaling. BMC Pulm. Med. 2014, 14, e122. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Kang, M.C.; Parveen, A.; Yumnam, S.; Kim, S.Y. Anti-inflammatory effect of chloroform fraction of Pyrus ussuriensis Maxim. leaf extract on 2,4-dinitrochlorobenzene-induced atopic dermatitis in nc/nga mice. Nutrients 2019, 11, 276. [Google Scholar] [CrossRef]
- Nie, L.; Peng, L.; Zhang, H.; Li, Y.F.; Tian, Y. Anti-inflammatory mechanism research of Astragalin on rat intestinaepithelial cell (lEC-6) inflammation model induced by LPS. Mod. Food Sci. Technol. 2020, 36, 17–23. [Google Scholar]
- Nygaard, G.; Firestein, G.S. Restoring synovial homeostasis in rheumatoid arthritis by targeting fibroblast-like synoviocytes. Nat. Rev. Rheumatol. 2020, 16, 316–333. [Google Scholar] [CrossRef]
- Medeiros, N.I.; Gomes, J.A.S.; Correa-Oliveira, R. Synergic and antagonistic relationship between MMP-2 and MMP-9 with fibrosis and inflammation in Chagas’ cardiomyopathy. Parasite Immunol. 2017, 39, e12446. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Matsuda, H.; Tanioka, M.; Kuwabara, K.; Itohara, S.; Suzuki, R. The role of matrix metalloproteinase-2 and matrix metalloproteinase-9 in antibody-induced arthritis. J. Immunol. 2002, 169, 2643–2647. [Google Scholar] [CrossRef]
- Jia, Q.; Wang, T.; Wang, X.; Xu, H.; Liu, Y.; Wang, Y.; Shi, Q.; Liang, Q. Astragalin suppresses inflammatory responses and bone destruction in mice with collagen-induced arthritis and in human fibroblast-like synoviocytes. Front. Pharmacol. 2019, 10, e94. [Google Scholar] [CrossRef]
- Kotani, M.; Matsumoto, M.; Fujita, A.; Higa, S.; Wang, W.; Suemura, M.; Kishimoto, T.; Tanaka, T. Persimmon leaf extract and astragalin inhibit development of dermatitis and IgE elevation in NC/Nga mice. J. Allergy Clin. Immunol. 2000, 106, 159–166. [Google Scholar] [CrossRef]
- Prajapati, V.D.; Maheriya, P.M.; Jani, G.K.; Solanki, H.K. Carrageenan: A natural seaweed polysaccharide and its applications. Carbohydr. Polym. 2014, 105, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Rocha, R.S.; Kassuya, C.A.; Formagio, A.S.; Mauro Mde, O.; Andrade-Silva, M.; Monreal, A.C.; Cunha-Laura, A.L.; Vieira Mdo, C.; Oliveira, R.J. Analysis of the anti-inflammatory and chemopreventive potential and description of the antimutagenic mode of action of the Annona crassiflora methanolic extract. Pharm. Biol. 2016, 54, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Parveen, Z.; Deng, Y.; Saeed, M.K.; Dai, R.; Ahamad, W.; Yu, Y.H. Antiinflammatory and analgesic activities of Thesium chinense Turcz extracts and its major flavonoids, kaempferol and kaempferol-3-O-glucoside. Yakugaku Zasshi 2007, 127, 1275–1279. [Google Scholar] [CrossRef] [PubMed]
- Alblihed, M.A. Astragalin attenuates oxidative stress and acute inflammatory responses in carrageenan-induced paw edema in mice. Mol. Biol. Rep. 2020, 47, 6611–6620. [Google Scholar] [CrossRef] [PubMed]
- Sroka-Tomaszewska, J.; Trzeciak, M. Molecular mechanisms of atopic dermatitis pathogenesis. Int. J. Mol. Sci. 2021, 22, 4130. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Kotani, M.; Fujita, A.; Higa, S.; Kishimoto, T.; Suemura, M.; Tanaka, T. Oral administration of persimmon leaf extract ameliorates skin symptoms and transepidermal water loss in atopic dermatitis model mice, NC/Nga. Br. J. Dermatol. 2002, 146, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Casaro, M.; Souza, V.R.; Oliveira, F.A.; Ferreira, C.M. OVA-induced allergic airway inflammation mouse model. Methods Mol. Biol. 2019, 1916, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.H.; Choi, Y.J.; Gong, J.H.; Shin, D.; Kang, M.K.; Kang, Y.H. Astragalin inhibits autophagy-associated airway epithelial fibrosis. Respir. Res. 2015, 16, e51. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, Y.; Zhang, X.; Zhang, X.; Chen, S.; Hu, Z.; Zhou, C.; Zhang, E.; Ma, S. Astragalin attenuates allergic inflammation in a murine asthma model. Inflammation 2015, 38, 2007–2016. [Google Scholar] [CrossRef]
- Ma, X.F.; Ma, L.G.; Liu, H.Y.; Wang, D.D.; Song, X.Y.; Song, Y.L.; Yan, G.H.; Li, L.C.; Sun, L.P. The effect of astragalin on airway inflammation in asthmatic mice. Immunol. J. 2022, 38, 944–949. [Google Scholar]
- Long, M.E.; Mallampalli, R.K.; Horowitz, J.C. Pathogenesis of pneumonia and acute lung injury. Clin. Sci. 2022, 13, 747–769. [Google Scholar] [CrossRef]
- Wu, M.Y.; Yiang, G.T.; Liao, W.T.; Tsai, A.P.; Cheng, Y.L.; Cheng, P.W.; Li, C.Y.; Li, C.J. Current mechanistic concepts in ischemia and reperfusion injury. Cell Physiol. Biochem. 2018, 46, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Algoet, M.; Janssens, S.; Himmelreich, U.; Gsell, W.; Pusovnik, M.; Van den Eynde, J.; Oosterlinck, W. Myocardial ischemia-reperfusion injury and the influence of inflammation. Trends Cardiovasc. Med. 2023, 33, 357–366. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, H.; Shi, J.; Huang, T.; Wang, Y. Astragalin protects against spinal cord ischemia reperfusion injury through attenuating oxidative stress-induced necroptosis. Biomed. Res. Int. 2021, 2021, e7254708. [Google Scholar] [CrossRef]
- Qu, D.; Han, J.; Ren, H.; Yang, W.; Zhang, X.; Zheng, Q.; Wang, D. Cardioprotective effects of astragalin against myocardial ischemia/reperfusion injury in isolated rat heart. Oxid. Med. Cell Longev. 2016, 2016, e8194690. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, X.; Qu, D.; Han, J.; Meng, F.; Xu, M.; Zheng, Q. Astragalin and dihydromyricetin as adjuncts to histidine-tryptophan-ketoglutarate cardioplegia enhances protection during cardioplegic arrest. Mol. Med. Rep. 2018, 18, 2929–2936. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, C.; Wang, L.F.; Kuang, X.; Liu, K.; Zhang, H.; Du, J.R. Neuroprotective effect of kaempferol glycosides against brain injury and neuroinflammation by inhibiting the activation of NF-κB and STAT3 in transient focal stroke. PLoS ONE 2013, 8, e55839. [Google Scholar] [CrossRef]
- Liu, D.; Gu, Y.; Wang, W.; Chen, W. Astragalin alleviates ischemia/reperfusion-induced brain injury via suppression of endoplasmic reticulum stress. Mol. Med. Rep. 2020, 22, 4070–4078. [Google Scholar] [CrossRef] [PubMed]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and depression: A review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef]
- Tong, Y.; Fu, H.; Xia, C.; Song, W.; Li, Y.; Zhao, J.; Zhang, X.; Gao, X.; Yong, J.; Liu, Q.; et al. Astragalin exerted antidepressant-like action through SIRT1 signaling modulated NLRP3 inflammasome deactivation. ACS Chem. Neurosci. 2020, 11, 1495–1503. [Google Scholar] [CrossRef]
- Yao, G.; Bai, Z.; Niu, J.; Zhang, R.; Lu, Y.; Gao, T.; Wang, H. Astragalin attenuates depression-like behaviors and memory deficits and promotes M2 microglia polarization by regulating IL-4R/JAK1/STAT6 signaling pathway in a murine model of perimenopausal depression. Psychopharmacology 2022, 239, 2421–2443. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target Ther. 2020, 5, e209. [Google Scholar] [CrossRef]
- Lee, S.U.; Lee, S.; Ro, H.; Choi, J.H.; Ryu, H.W.; Kim, M.O.; Yuk, H.J.; Lee, J.; Hong, S.T.; Oh, S.R. Piscroside C inhibits TNF-α/NF-κB pathway by the suppression of PKCδ activity for TNF-RSC formation in human airway epithelial cells. Phytomedicine 2018, 40, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.M.; Koh, J.; Kim, J.H.; Lee, J.; Im, J.P.; Kim, J.S. Astragalin inhibits nuclear factor-κB signaling in human colonic epithelial cells and attenuates experimental colitis in mice. Gut Liver 2021, 15, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, W.Y.; Xie, J.; Wang, Z.L.; Wen, Y.L.; Zhao, C.C.; Tao, L.; Li, L.F.; Tian, Y.; Sheng, J. Astragalin inhibits the proliferation and migration of human colon cancer HCT116 cells by regulating the NF-κB signaling pathway. Front. Pharmacol. 2021, 12, e639256. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liang, D.; Yang, Z.; Wang, T.; Wang, W.; Song, X.; Guo, M.; Zhou, E.; Li, D.; Cao, Y.; et al. Astragalin suppresses inflammatory responses via down-regulation of NF-κB signaling pathway in lipopolysaccharide-induced mastitis in a murine model. Int. Immunopharmacol. 2013, 17, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, W.; Cao, Y.; Liang, D.; Zhang, W.; Zhang, Z.; Jiang, H.; Guo, M.; Zhang, N. Inhibitory effects of astragalin on lipopolysaccharide-induced inflammatory response in mouse mammary epithelial cells. J. Surg. Res. 2014, 192, 573–581. [Google Scholar] [CrossRef]
- Zusso, M.; Lunardi, V.; Franceschini, D.; Pagetta, A.; Lo, R.; Stifani, S.; Frigo, A.C.; Giusti, P.; Moro, S. Ciprofloxacin and levofloxacin attenuate microglia inflammatory response via TLR4/NF-κB pathway. J. Neuroinflamm. 2019, 16, e148. [Google Scholar] [CrossRef]
- Peng, L.; Gao, X.; Nie, L.; Xie, J.; Dai, T.; Shi, C.; Tao, L.; Wang, Y.; Tian, Y.; Sheng, J. Astragalin attenuates dextran sulfate sodium (DSS)-induced acute experimental colitis by alleviating gut microbiota dysbiosis and inhibiting NF-κB activation in mice. Front. Immunol. 2020, 11, e2058. [Google Scholar] [CrossRef]
- Hu, X.; Wang, M.; Pan, Y.; Xie, Y.; Han, J.; Zhang, X.; Niayale, R.; He, H.; Li, Q.; Zhao, T.; et al. Anti-inflammatory effect of astragalin and chlorogenic acid on escherichia coli-induced inflammation of sheep endometrial epithelium cells. Front. Vet. Sci. 2020, 7, e201. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.W.; Zhang, M.; Wang, G.; Zou, J.; Gao, J.H.; Zhou, L.; Wan, X.J.; Zhang, D.W.; Yu, X.H.; Tang, C.K. Astragalin retards atherosclerosis by promoting cholesterol efflux and inhibiting the inflammatory response via upregulating ABCA1 and ABCG1 expression in macrophages. J. Cardiovasc. Pharmacol. 2021, 77, 217–227. [Google Scholar] [CrossRef]
- Osipo, C.; Golde, T.E.; Osborne, B.A.; Miele, L.A. Off the beaten pathway: The complex cross talk between Notch and NF-kappaB. Lab. Investig. 2008, 88, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Fang, X.; Wang, J.; Ren, T.T.; Zhao, Y.Y.; Dai, J.F.; Qin, X.Y.; Lan, R. Astragalin attenuates AlCl3/D-galactose-induced aging-like disorders by inhibiting oxidative stress and neuroinflammation. Neurotoxicology 2022, 91, 60–68. [Google Scholar] [CrossRef]
- Park, J.H.; Seo, Y.H.; Jang, J.H.; Jeong, C.H.; Lee, S.; Park, B. Asiatic acid attenuates methamphetamine-induced neuroinflammation and neurotoxicity through blocking of NF-κB/STAT3/ERK and mitochondria-mediated apoptosis pathway. J. Neuroinflammation 2017, 14, e240. [Google Scholar] [CrossRef] [PubMed]
- Innamorato, N.G.; Rojo, A.I.; García-Yagüe, A.J.; Yamamoto, M.; de Ceballos, M.L.; Cuadrado, A. The transcription factor Nrf2 is a therapeutic target against brain inflammation. J. Immunol. 2008, 181, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, K.; Gao, X.; Zhao, K.; Chen, H.; Xu, M. Anti-inflammatory effects of isoalantolactone on LPS-stimulated BV2 microglia cells through activating GSK-3β-Nrf2 signaling pathway. Int. mmunopharmacol. 2018, 65, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cheng, C.; Zuo, X.; Huang, W. Astragalin alleviates cerebral ischemia-reperfusion injury by improving anti-oxidant and anti-inflammatory activities and inhibiting apoptosis pathway in rats. BMC Complement. Med. Ther. 2020, 20, e120. [Google Scholar] [CrossRef] [PubMed]
- Sul, O.J.; Ra, S.W. Quercetin prevents LPS-Induced oxidative stress and inflammation by modulating NOX2/ROS/NF-κB in lung epithelial cells. Molecules 2021, 26, 6949. [Google Scholar] [CrossRef]
- Hu, C.; Wu, Z.; Huang, Z.; Hao, X.; Wang, S.; Deng, J.; Yin, Y.; Tan, C. Nox2 impairs VEGF-A-induced angiogenesis in placenta via mitochondrial ROS-STAT3 pathway. Redox Biol. 2021, 45, e102051. [Google Scholar] [CrossRef]
- Ohayon, D.; De Chiara, A.; Dang, P.M.; Thieblemont, N.; Chatfield, S.; Marzaioli, V.; Burgener, S.S.; Mocek, J.; Candalh, C.; Pintard, C.; et al. Cytosolic PCNA interacts with p47phox and controls NADPH oxidase NOX2 activation in neutrophils. J. Exp. Med. 2019, 216, 2669–2687. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.F.; Liu, H.Y.; Wang, D.D.; Song, Y.L.; Yan, G.H.; Li, L.C. Astragalin inhibits airway inflammation in asthma through NOX2/ROS/NF-KB signaling pathway. Chin. Pharmacol. Bull. 2021, 37, 797–802. [Google Scholar]
- Shen, Z.; Ajmo, J.M.; Rogers, C.Q.; Liang, X.; Le, L.; Murr, M.M.; Peng, Y.; You, M. Role of SIRT1 in regulation of LPS- or two ethanol metabolites-induced TNF-alpha production in cultured macrophage cell lines. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1047–G1053. [Google Scholar] [CrossRef] [PubMed]
- de Lamirande, E.; O’Flaherty, C. Sperm activation: Role of reactive oxygen species and kinases. Biochim. Biophys. Acta 2008, 1784, 106–115. [Google Scholar] [CrossRef]
- Marshall, C.J. MAP kinase kinase kinase, MAP kinase kinase and MAP kinase. Curr. Opin. Genet. Dev. 1994, 4, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Plastira, I.; Bernhart, E.; Joshi, L.; Koyani, C.N.; Strohmaier, H.; Reicher, H.; Malle, E.; Sattler, W. MAPK signaling determines lysophosphatidic acid (LPA)-induced inflammation in microglia. J. Neuroinflamm. 2020, 17, e127. [Google Scholar] [CrossRef] [PubMed]
- Bogoyevitch, M.A.; Boehm, I.; Oakley, A.; Ketterman, A.J.; Barr, R.K. Targeting the JNK MAPK cascade for inhibition: Basic science and therapeutic potential. Biochim. Biophys. Acta 2004, 1697, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef]
- Yang, B.C.; Fu, X.J.; Zhu, L. Effect of Astragalus Glycosides on JNK/p38 MAPK signaling pathway in rats with knee osteoarthritis. Chin. J. Osteoporos. 2022, 28, 1491–1495+1500. [Google Scholar]
- Zhang, W.; Lu, X.; Wang, W.; Ding, Z.; Fu, Y.; Zhou, X.; Zhang, N.; Cao, Y. Inhibitory effects of emodin, thymol, and astragalin on leptospira interrogans-induced inflammatory response in the uterine and endometrium epithelial cells of mice. Inflammation 2017, 40, 666–675. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kang, M.K.; Lee, E.J.; Kim, D.Y.; Oh, H.; Kim, S.I.; Oh, S.Y.; Na, W.; Shim, J.H.; Kang, I.J.; et al. Astragalin inhibits cigarette smoke-induced pulmonary thrombosis and alveolar inflammation and disrupts PAR activation and oxidative stress-responsive MAPK-signaling. Int. J. Mol. Sci. 2021, 22, 3692. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Gan, C.; Zhu, J.; Ma, N.; Wu, L.; Wang, L.; Wang, X. Anti-atherosclerotic activities of flavonoids from the flowers of Helichrysum arenarium L. MOENCH through the pathway of anti-inflammation. Bioorg. Med. Chem. Lett. 2017, 27, 2812–2817. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.T.; Lin, C.S.; Liao, Y.C.; Ho, L.J.; Yang, S.P.; Lai, J.H. JNK/AP-1 activation contributes to tetrandrine resistance in T-cell acute lymphoblastic leukaemia. Acta Pharmacol. Sin. 2017, 38, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Chung, Y.H.; Ahn, H.; Kim, H.; Rho, J.; Jeong, D. Selective regulation of MAPK signaling mediates RANKL-dependent osteoclast differentiation. Int. J. Biol. Sci. 2016, 12, 235–245. [Google Scholar] [CrossRef]
- Xing, F.; Geng, L.; Guan, H.; Liu, D.; Li, Y.; Zeng, L.; Chen, Y.; Tian, R.; Li, Z.; Cao, R.; et al. Astragalin mitigates inflammatory osteolysis by negatively modulating osteoclastogenesis via ROS and MAPK signaling pathway. Int. Immunopharmacol. 2022, 112, e109278. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK-STAT signaling as a target for inflammatory and autoimmune diseases: Current and future prospects. Drugs 2017, 77, 521–546. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Hanh Nguyen, T.T.; Hur, J.; Kim, N.M.; Kim, S.B.; Hwang, K.H.; Moon, Y.H.; Kang, C.; Chung, B.; Kim, Y.M.; et al. Synthesis and characterization of novel astragalin galactosides using β-galactosidase from Bacillus circulans. Enzyme Microb. Technol. 2017, 103, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Choung, W.J.; Hwang, S.H.; Ko, D.S.; Kim, S.B.; Kim, S.H.; Jeon, S.H.; Choi, H.D.; Lim, S.S.; Shim, J.H. Enzymatic synthesis of a novel kaempferol-3-O-β-D-glucopyranosyl-(1→4)-O-α-D-glucopyranoside using cyclodextrin glucanotransferase and its inhibitory effects on aldose reductase, inflammation, and oxidative stress. J. Agric. Food Chem. 2017, 65, 2760–2767. [Google Scholar] [CrossRef]
- Pei, J.; Dong, P.; Wu, T.; Zhao, L.; Fang, X.; Cao, F.; Tang, F.; Yue, Y. Metabolic engineering of Escherichia coli for astragalin biosynthesis. J. Agric. Food Chem. 2016, 64, 7966–7972. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Cell | Induction Method | Testing Index | Refs. |
|---|---|---|---|---|
| Macrophages | RAW264.7 | LPS + IFN-γ | NO ↓, TNF-α ↓, PGE2 ↓, IL-1β ↓, IL-6 ↓, IL-12 ↓, IL-13 ↓, IL-18 ↓, LTB4 ↓; IL-10 ↑ | [31,32,33,34] |
| J774A.1 | LPS | iNOS ↓, COX ↓, TNF-α ↓, IL-1β ↓, IL-6 ↓, MIP-1α ↓, MCP-1 ↓ | [35] | |
| Microglia cells | Mice microglia cell | LPS | iNOS ↓, NO ↓ | [38,39] |
| BV2 microglial cell | LPS | iNOS ↓, COX-2 ↓, PTGES2 ↓, TNF-α ↓, IL-1β ↓, IL-6 ↓, NO ↓ | [38] | |
| Epithelial cells | BEAS-2B cell | H2O2 | ICAM-1 ↓, MCP-1 ↓, MCP-1/ICAM-1/αv ↓ | [42] |
| BEAS-2B cell | LPS/H2O2 | TLR4 ↓, Eotaxin-1 ↓, PLCγ1 ↓, PKCβ2 ↓, p22phox ↓, p47phox | [43] | |
| HaCaT | TNF-α/IFN-γ | IL-6 ↓, IL-1β ↓ | [44] | |
| IEC-6 | LPS | TNF-α ↓, IL-6 ↓ | [45] | |
| Other types of cells | RA-FLS cell | TNF-α | MMP-1 ↓, MMP-3 ↓, MMP-13 ↓ | [49] |
| KU812 | FcεRI + CRA-1 | Histamine ↓ | [50] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, J.; Shi, Z.; Cao, X.; Dang, Z.; Zhang, Q.; Zhang, W.; Wu, L.; Zhang, Y.; Wang, T. Research Progress on Anti-Inflammatory Effects and Related Mechanisms of Astragalin. Int. J. Mol. Sci. 2024, 25, 4476. https://doi.org/10.3390/ijms25084476
Ruan J, Shi Z, Cao X, Dang Z, Zhang Q, Zhang W, Wu L, Zhang Y, Wang T. Research Progress on Anti-Inflammatory Effects and Related Mechanisms of Astragalin. International Journal of Molecular Sciences. 2024; 25(8):4476. https://doi.org/10.3390/ijms25084476
Chicago/Turabian StyleRuan, Jingya, Zhongwei Shi, Xiaoyan Cao, Zhunan Dang, Qianqian Zhang, Wei Zhang, Lijie Wu, Yi Zhang, and Tao Wang. 2024. "Research Progress on Anti-Inflammatory Effects and Related Mechanisms of Astragalin" International Journal of Molecular Sciences 25, no. 8: 4476. https://doi.org/10.3390/ijms25084476