
Differential Response of Human Dendritic Cells upon Stimulation with Encapsulated or Non-Encapsulated Isogenic Strains of Porphyromonas gingivalis
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
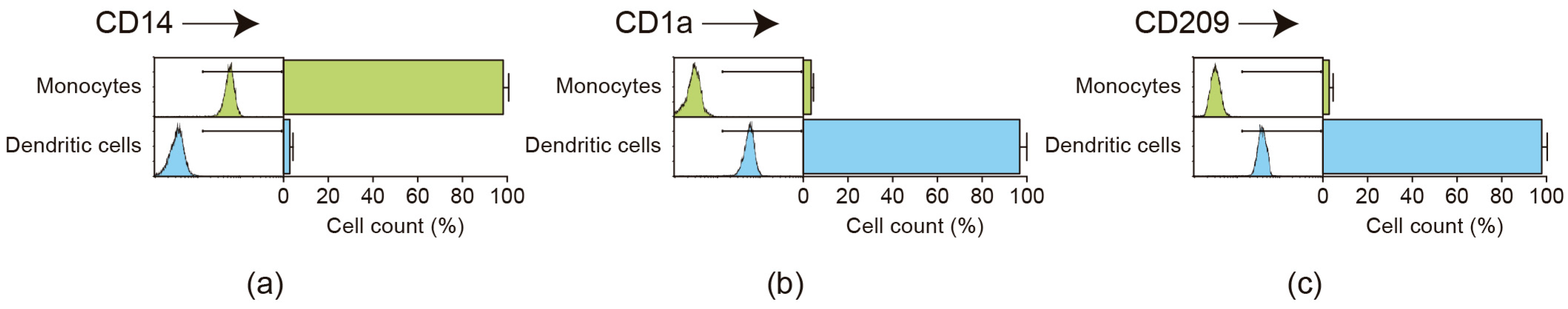
2.1. Flow Cytometric Analysis of Monocyte Purification and Dendritic Cell Differentiation
2.2. Flow Cytometric Analysis of Dendritic Cell Maturation
2.3. Toll-like Receptor Expression in P. gingivalis-Infected Dendritic Cells
2.4. Transcription Factor Expression in P. gingivalis-Infected Dendritic Cells
2.5. Cytokine Expression in P. gingivalis-Infected Dendritic Cells
2.6. Cytokine Secretion in P. gingivalis-Infected Dendritic Cells
3. Discussion
4. Materials and Methods
4.1. Porphyromonas gingivalis Strains
4.2. Individuals
4.3. Monocyte-Derived Dendritic Cell Generation
4.4. Dendritic Cell Infection
4.5. Flow Cytometry Analysis
4.6. Expression of Cytokines, Transcription Factors, and Toll-like Receptors
4.7. Secretion of Cytokines
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Rev. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Tonetti, M.S.; Jepsen, S.; Jin, L.; Otomo-Corgel, J. Impact of the global burden of periodontal diseases on health, nutrition and wellbeing of mankind: A call for global action. J. Clin. Periodontol. 2017, 44, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Peres, M.A.; Macpherson, L.M.D.; Weyant, R.J.; Daly, B.; Venturelli, R.; Mathur, M.R.; Listl, S.; Celeste, R.K.; Guarnizo-Herreno, C.C.; Kearns, C.; et al. Oral diseases: A global public health challenge. Lancet 2019, 394, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Kassebaum, N.J.; Smith, A.G.C.; Bernabe, E.; Fleming, T.D.; Reynolds, A.E.; Vos, T.; Murray, C.J.L.; Marcenes, W.; Collaborators, G.B.D.O.H. Global, regional, and national prevalence, incidence, and disability-adjusted life years for oral conditions for 195 countries, 1990–2015: A systematic analysis for the global burden of diseases, injuries, and risk factors. J. Dent. Res. 2017, 96, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Trindade, D.; Carvalho, R.; Machado, V.; Chambrone, L.; Mendes, J.J.; Botelho, J. Prevalence of periodontitis in dentate people between 2011 and 2020: A systematic review and meta-analysis of epidemiological studies. J. Clin. Periodontol. 2023, 50, 604–626. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.D.; Papapanou, P.N.; Philips, K.H.; Offenbacher, S. Periodontal Medicine: 100 Years of Progress. J. Dent. Res. 2019, 98, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, S.Q.; Zhao, L.; Ren, Z.H.; Hu, C.Y. Global, regional, and national burden of periodontitis from 1990 to 2019: Results from the Global Burden of Disease study 2019. J. Periodontol. 2022, 93, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Chavakis, T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Lamont, R.J. Polymicrobial communities in periodontal disease: Their quasi-organismal nature and dialogue with the host. Periodontology 2000 2021, 86, 210–230. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.; Monasterio, G.; Cavalla, F.; Córdova, L.A.; Hernández, M.; Heymann, D.; Garlet, G.P.; Sorsa, T.; Pärnänen, P.; Lee, H.M.; et al. Osteoimmunology of oral and maxillofacial diseases: Translational applications based on biological mechanisms. Front. Immunol. 2019, 10, 1664. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.T.; Oates, T.; Garlet, G.P. Review of osteoimmunology and the host response in endodontic and periodontal lesions. J. Oral Microbiol. 2011, 3, 5304. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. Interconnection of periodontal disease and comorbidities: Evidence, mechanisms, and implications. Periodontology 2000 2022, 89, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Garlet, G.P. Destructive and protective roles of cytokines in periodontitis: A re-appraisal from host defense and tissue destruction viewpoints. J. Dent. Res. 2010, 89, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.; Michea, M.A.; Vegvari, A.; Arce, M.; Morales, A.; Lanyon, E.; Alcota, M.; Fuentes, C.; Vernal, R.; Budini, M.; et al. Proteomic profile of human gingival crevicular fluid reveals specific biological and molecular processes during clinical progression of periodontitis. J. Periodontal Res. 2023, 58, 1061–1081. [Google Scholar] [CrossRef] [PubMed]
- Buetas, E.; Jordan-Lopez, M.; Lopez-Roldan, A.; D’Auria, G.; Martinez-Priego, L.; De Marco, G.; Carda-Dieguez, M.; Mira, A. Full-length 16S rRNA gene sequencing by PacBio improves taxonomic resolution in human microbiome samples. BMC Genom. 2024, 25, 310. [Google Scholar] [CrossRef] [PubMed]
- Scannapieco, F.A.; Dongari-Bagtzoglou, A. Dysbiosis revisited: Understanding the role of the oral microbiome in the pathogenesis of gingivitis and periodontitis: A critical assessment. J. Periodontol. 2021, 92, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- de Jongh, C.A.; de Vries, T.J.; Bikker, F.J.; Gibbs, S.; Krom, B.P. Mechanisms of Porphyromonas gingivalis to translocate over the oral mucosa and other tissue barriers. J. Oral Microbiol. 2023, 15, 2205291. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.A.; Dou, Y.; Fletcher, H.M.; Boskovic, D.S. Local and systemic effects of Porphyromonas gingivalis infection. Microorganisms 2023, 11, 470. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Belmar, J.; Morales-Olavarría, M.; Vicencio, E.; Vernal, R.; Cárdenas, J.P.; Cortez, C. Contribution of -omics technologies in the study of Porphyromonas gingivalis during periodontitis pathogenesis: A minireview. Int. J. Mol. Sci. 2023, 24, 620. [Google Scholar] [CrossRef] [PubMed]
- Lunar Silva, I.; Cascales, E. Molecular strategies underlying Porphyromonas gingivalis virulence. J. Mol. Biol. 2021, 433, 166836. [Google Scholar] [CrossRef] [PubMed]
- Rocha, F.G.; Berges, A.; Sedra, A.; Ghods, S.; Kapoor, N.; Pill, L.; Davey, M.E.; Fairman, J.; Gibson, F.C., 3rd. A Porphyromonas gingivalis capsule-conjugate vaccine protects from experimental oral bone loss. Front. Oral Health 2021, 2, 686402. [Google Scholar] [CrossRef] [PubMed]
- Bregaint, S.; Boyer, E.; Fong, S.B.; Meuric, V.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Porphyromonas gingivalis outside the oral cavity. Odontology 2022, 110, 1–19. [Google Scholar] [CrossRef]
- Boyer, E.; Leroyer, P.; Malherbe, L.; Fong, S.B.; Loreal, O.; Bonnaure Mallet, M.; Meuric, V. Oral dysbiosis induced by Porphyromonas gingivalis is strain-dependent in mice. J. Oral Microbiol. 2020, 12, 1832837. [Google Scholar] [CrossRef] [PubMed]
- Monasterio, G.; Fernández, B.; Castillo, F.; Rojas, C.; Cafferata, E.A.; Rojas, L.; Alvarez, C.; Fernández, A.; Hernández, M.; Bravo, D.; et al. Capsular-defective Porphyromonas gingivalis mutant strains induce less alveolar bone resorption than W50 wild-type strain due to a decreased Th1/Th17 immune response and less osteoclast activity. J. Periodontol. 2019, 90, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Polak, D.; Ferdman, O.; Houri-Haddad, Y. Porphyromonas gingivalis capsule-mediated coaggregation as a virulence factor in mixed infection with Fusobacterium nucleatum. J. Periodontol. 2017, 88, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Vernal, R.; Diaz-Guerra, E.; Silva, A.; Sanz, M.; Garcia-Sanz, J.A. Distinct human T-lymphocyte responses triggered by Porphyromonas gingivalis capsular serotypes. J. Clin. Periodontol. 2014, 41, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Vernal, R.; Díaz-Zúñiga, J.; Melgar-Rodríguez, S.; Pujol, M.; Díaz-Guerra, E.; Silva, A.; Sanz, M.; Garcia-Sanz, J.A. Activation of RANKL-induced osteoclasts and memory T lymphocytes by Porphyromonas gingivalis is serotype dependant. J. Clin. Periodontol. 2014, 41, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, C.M.F.; Maltos, K.L.M.; Shehabeldin, M.S.; Thomas, L.L.; Zhuang, Z.; Yoshizawa, S.; Verdelis, K.; Gaffen, S.L.; Garlet, G.P.; Little, S.R.; et al. Local sustained delivery of anti-IL-17A antibodies limits inflammatory bone loss in murine experimental periodontitis. J. Immunol. 2021, 206, 2386–2392. [Google Scholar] [CrossRef] [PubMed]
- Sosa Cuevas, E.; Saas, P.; Aspord, C. Dendritic cell subsets in melanoma: Pathophysiology, clinical prognosis and therapeutic exploitation. Cancers 2023, 15, 2206. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, A.; Salvi, V.; Soriani, A.; Laffranchi, M.; Sozio, F.; Bosisio, D.; Sozzani, S. Dendritic cell subsets in cancer immunity and tumor antigen sensing. Cell Mol. Immunol. 2023, 20, 432–447. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Dong, G.; Guo, L.; Graves, D.T. The function of dendritic cells in modulating the host response. Mol. Oral Microbiol. 2018, 33, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, N.; Kumamoto, Y. Role of mouse dendritic cell subsets in priming naive CD4 T cells. Curr. Opin. Immunol. 2023, 83, 102352. [Google Scholar] [CrossRef] [PubMed]
- Schlitzer, A.; McGovern, N.; Teo, P.; Zelante, T.; Atarashi, K.; Low, D.; Ho, A.W.; See, P.; Shin, A.; Wasan, P.S.; et al. IRF4 transcription factor-dependent CD11b+ dendritic cells in human and mouse control mucosal IL-17 cytokine responses. Immunity 2013, 38, 970–983. [Google Scholar] [CrossRef] [PubMed]
- Mildner, A.; Jung, S. Development and function of dendritic cell subsets. Immunity 2014, 40, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, A.T.; Wu, X.; Albring, J.C.; Murphy, K.M. Re(de)fining the dendritic cell lineage. Nat. Immunol. 2012, 13, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Monasterio, G.; Guevara, J.; Ibarra, J.P.; Castillo, F.; Díaz-Zúñiga, J.; Alvarez, C.; Cafferata, E.A.; Vernal, R. Immunostimulatory activity of low-molecular-weight hyaluronan on dendritic cells stimulated with Aggregatibacter actinomycetemcomitans or Porphyromonas gingivalis. Clin. Oral Investig. 2019, 23, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Bedoui, S.; Heath, W.R. Krüppel-ling of IRF4-dependent DCs into two functionally distinct DC subsets. Immunity 2015, 42, 785–787. [Google Scholar] [CrossRef]
- Chandra, J.; Kuo, P.T.; Hahn, A.M.; Belz, G.T.; Frazer, I.H. Batf3 selectively determines acquisition of CD8+ dendritic cell phenotype and function. Immunol. Cell Biol. 2017, 95, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Nizzoli, G.; Krietsch, J.; Weick, A.; Steinfelder, S.; Facciotti, F.; Gruarin, P.; Bianco, A.; Steckel, B.; Moro, M.; Crosti, M.; et al. Human CD1c+ dendritic cells secrete high levels of IL-12 and potently prime cytotoxic T-cell responses. Blood 2013, 122, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Persson, E.K.; Uronen-Hansson, H.; Semmrich, M.; Rivollier, A.; Hägerbrand, K.; Marsal, J.; Gudjonsson, S.; Håkansson, U.; Reizis, B.; Kotarsky, K.; et al. IRF4 transcription-factor-dependent CD103+CD11b+ dendritic cells drive mucosal T helper 17 cell differentiation. Immunity 2013, 38, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Tussiwand, R.; Gautier, E.L. Transcriptional regulation of mononuclear phagocyte development. Front. Immunol. 2015, 6, 533. [Google Scholar] [CrossRef] [PubMed]
- Geginat, J.; Nizzoli, G.; Paroni, M.; Maglie, S.; Larghi, P.; Pascolo, S.; Abrignani, S. Immunity to pathogens taught by specialized human dendritic cell subsets. Front. Immunol. 2015, 6, 527. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Zúñiga, J.; Monasterio, G.; Alvarez, C.; Melgar-Rodríguez, S.; Benítez, A.; Ciuchi, P.; García, M.; Arias, J.; Sanz, M.; Vernal, R. Variability of the dendritic cell response triggered by different serotypes of Aggregatibacter actinomycetemcomitans or Porphyromonas gingivalis is toll-like receptor 2 (TLR2) or TLR4 dependent. J. Periodontol. 2015, 86, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Vernal, R.; León, R.; Silva, A.; van Winkelhoff, A.J.; García-Sanz, J.A.; Sanz, M. Differential cytokine expression by human dendritic cells in response to different Porphyromonas gingivalis capsular serotypes. J. Clin. Periodontol. 2009, 36, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, S.R.; D’Epiro, T.T.; Pinheiro, E.T.; Simionato, M.R.; Taniwaki, N.N.; Kisielius, J.J.; Mayer, M.P. Lineage variability in surface components expression within Porphyromonas gingivalis. Microb. Pathog. 2014, 77, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Ranjit, D.K.; Walker, A.R.; Getachew, H.; Progulske-Fox, A.; Davey, M.E. A novel regulation of K-antigen capsule synthesis in Porphyromonas gingivalis is driven by the response regulator PG0720-directed antisense RNA. Front. Oral Health 2021, 2, 701659. [Google Scholar] [CrossRef] [PubMed]
- Aduse-Opoku, J.; Slaney, J.M.; Hashim, A.; Gallagher, A.; Gallagher, R.P.; Rangarajan, M.; Boutaga, K.; Laine, M.L.; Van Winkelhoff, A.J.; Curtis, M.A. Identification and characterization of the capsular polysaccharide (K-antigen) locus of Porphyromonas gingivalis. Infect. Immun. 2006, 74, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Laine, M.L.; van Winkelhoff, A.J.; Dahlen, G. Genotype variation and capsular serotypes of Porphyromonas gingivalis from chronic periodontitis and periodontal abscesses. FEMS Microbiol. Lett. 2007, 270, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Lamont, R.J. Breaking bad: Manipulation of the host response by Porphyromonas gingivalis. Eur. J. Immunol. 2014, 44, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, F.; Murakami, Y.; Nishikawa, K.; Hasegawa, Y.; Kawaminami, S. Surface components of Porphyromonas gingivalis. J. Periodontal Res. 2009, 44, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dierickx, K.; Pauwels, M.; Laine, M.L.; Van Eldere, J.; Cassiman, J.J.; van Winkelhoff, A.J.; van Steenberghe, D.; Quirynen, M. Adhesion of Porphyromonas gingivalis serotypes to pocket epithelium. J. Periodontol. 2003, 74, 844–848. [Google Scholar] [CrossRef] [PubMed]
- Rosen, G.; Sela, M.N. Coaggregation of Porphyromonas gingivalis and Fusobacterium nucleatum PK 1594 is mediated by capsular polysaccharide and lipopolysaccharide. FEMS Microbiol. Lett. 2006, 256, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, V.; Utreja, S.; Hovey, K.M.; Sun, Y.; LaMonte, M.J.; Wactawski-Wende, J.; Diaz, P.I.; Buck, M.J. Defining Porphyromonas gingivalis strains associated with periodontal disease. Sci. Rep. 2024, 14, 6222. [Google Scholar] [CrossRef] [PubMed]
- Laine, M.L.; Appelmelk, B.J.; van Winkelhoff, A.J. Novel polysaccharide capsular serotypes in Porphyromonas gingivalis. J. Periodontal Res. 1996, 31, 278–284. [Google Scholar] [CrossRef] [PubMed]
- van Winkelhoff, A.J.; Appelmelk, B.J.; Kippuw, N.; de Graaff, J. K-antigens in Porphyromonas gingivalis are associated with virulence. Oral Microbiol. Immunol. 1993, 8, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Igboin, C.O.; Griffen, A.L.; Leys, E.J. Porphyromonas gingivalis strain diversity. J. Clin. Microbiol. 2009, 47, 3073–3081. [Google Scholar] [CrossRef] [PubMed]
- Naito, M.; Hirakawa, H.; Yamashita, A.; Ohara, N.; Shoji, M.; Yukitake, H.; Nakayama, K.; Toh, H.; Yoshimura, F.; Kuhara, S.; et al. Determination of the genome sequence of Porphyromonas gingivalis strain ATCC 33277 and genomic comparison with strain W83 revealed extensive genome rearrangements in P. gingivalis. DNA Res. 2008, 15, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Cortegana, C.; Palomares, F.; Alba, G.; Santa-María, C.; de la Cruz-Merino, L.; Sánchez-Margalet, V.; López-Enriquez, S. Dendritic cells: The yin and yang in disease progression. Front. Immunol. 2023, 14, 1321051. [Google Scholar] [CrossRef] [PubMed]
- Acuto, O. T-cell virtuosity in “knowing thyself”. Front. Immunol. 2024, 15, 1343575. [Google Scholar] [CrossRef]
- Behzadi, P.; Garcia-Perdomo, H.A.; Karpinski, T.M. Toll-Like receptors: General molecular and structural biology. J. Immunol. Res. 2021, 2021, 9914854. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Zúñiga, J.; Yánez, J.P.; Alvarez, C.; Melgar-Rodríguez, S.; Hernández, M.; Sanz, M.; Vernal, R. Serotype-dependent response of human dendritic cells stimulated with Aggregatibacter actinomycetemcomitans. J. Clin. Periodontol. 2014, 41, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Hahne, F.; Gopalakrishnan, N.; Khodabakhshi, A.; Wong, C.; Lee, K. flowStats: Statistical Methods for the Analysis of Flow Cytometry Data. 2023. Available online: http://www.github.com/RGLab/flowStatsBioinformatics (accessed on 20 April 2023).
- Van, P.; Jiang, W.; Gottardo, R.; Finak, G. ggcyto: Next-generation open-source visualization software for cytometry. Bioinformatics 2018, 34, 3951–3953. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Clone | Fluorochrome | Dilution | Supplier | Code |
|---|---|---|---|---|---|
| anti-CD1a | HI149 | APC-Cy7 2 | 1:400 | Biolegend 7 | 300126 |
| anti-CD3 | OKT3 | eFluor 660 | 1:800 | eBioscience 8 | 50003741 |
| anti-CD4 | RPA-T4 | BUV395 3 | 1:200 | BD Biosciences 9 | 564724 |
| anti-CD8 | SK1 | BV510 4 | 1:200 | Biolegend | 344732 |
| anti-CD14 | M5E2 | BV650 | 1:200 | Biolegend | 301835 |
| anti-CD45 | 2D1 | Alexa Fluor 700 | 1:800 | eBioscience | 56945941 |
| anti-CD80 | W17149D | PerCP-Cy5.5 5 | 1:400 | Biolegend | 375412 |
| anti-CD83 | HB15e | BV711 | 1:400 | Biolegend | 305333 |
| anti-CD86 | IT2.2 | PE-Cy7 6 | 1:400 | eBioscience | 25086941 |
| anti-CD209 | 9E9A8 | BV421 | 1:400 | Biolegend | 330118 |
| Cell viability kit 1 | -- | BUV496 | 1:1000 | Biolegend | 423108 |
| mRNA | Forward Primer | Reverse Primer |
|---|---|---|
| IL-1β | ctgtcctgcgtgttgaaaga | ttgggtaatttttgggatctaca |
| IL-6 | gcccagctatgaactccttct | gaaggcagcaggcaacac |
| IL-10 | tgggggagaacctgaagac | ccttgctcttgttttcacagg |
| IL-12p35 | cactcccaaaacctgctgag | tctcttcagaagtgcaagggta |
| IL-23 | agcttcatgcctccctactg | ctgctgagtctcccagtggt |
| IFN-γ | ggcattttgaagaattggaaag | tttggatgctctggtcatctt |
| TNF-α | cagcctcttctccttcctgat | gccagagggctgattagaga |
| TGF-β1 | cacgtggagctgtaccagaa | cagccggttgctgaggta |
| IRF4 1 | gacaacgccttacccttcg | aggggtggcatcatgtagtt |
| IRF8 2 | tggggatgatcaaaaggagcc | aactggctggtgtcgaagac |
| NOTCH2 3 | cagttacccacccacaggtc | ccatacaggcagtcaatggaa |
| BATF3 4 | agacccagaaggctgacaag | ctccgcagcatggtgttt |
| TLR2 | ctctcggtgtcggaatgtc | aggatcagcaggaacagagc |
| TLR4 | ccctcccctgtaccttct | tccctgccttgaataccttc |
| 18S rRNA | ctcaacacgggaaacctcac | cgctccaccaactaagaacg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melgar-Rodríguez, S.; Polanco, A.; Ríos-Muñoz, J.; García, M.; Sierra-Cristancho, A.; González-Osuna, L.; Díaz-Zúñiga, J.; Carvajal, P.; Vernal, R.; Bravo, D. Differential Response of Human Dendritic Cells upon Stimulation with Encapsulated or Non-Encapsulated Isogenic Strains of Porphyromonas gingivalis. Int. J. Mol. Sci. 2024, 25, 4510. https://doi.org/10.3390/ijms25084510
Melgar-Rodríguez S, Polanco A, Ríos-Muñoz J, García M, Sierra-Cristancho A, González-Osuna L, Díaz-Zúñiga J, Carvajal P, Vernal R, Bravo D. Differential Response of Human Dendritic Cells upon Stimulation with Encapsulated or Non-Encapsulated Isogenic Strains of Porphyromonas gingivalis. International Journal of Molecular Sciences. 2024; 25(8):4510. https://doi.org/10.3390/ijms25084510
Chicago/Turabian StyleMelgar-Rodríguez, Samanta, Alan Polanco, Jearitza Ríos-Muñoz, Michelle García, Alfredo Sierra-Cristancho, Luis González-Osuna, Jaime Díaz-Zúñiga, Paola Carvajal, Rolando Vernal, and Denisse Bravo. 2024. "Differential Response of Human Dendritic Cells upon Stimulation with Encapsulated or Non-Encapsulated Isogenic Strains of Porphyromonas gingivalis" International Journal of Molecular Sciences 25, no. 8: 4510. https://doi.org/10.3390/ijms25084510