

APOE ε2-Carriers Are Associated with an Increased Risk of Primary Angle-Closure Glaucoma in Patients of Saudi Origin
 , , ,
, , , 
Abstract
:1. Introduction
2. Results
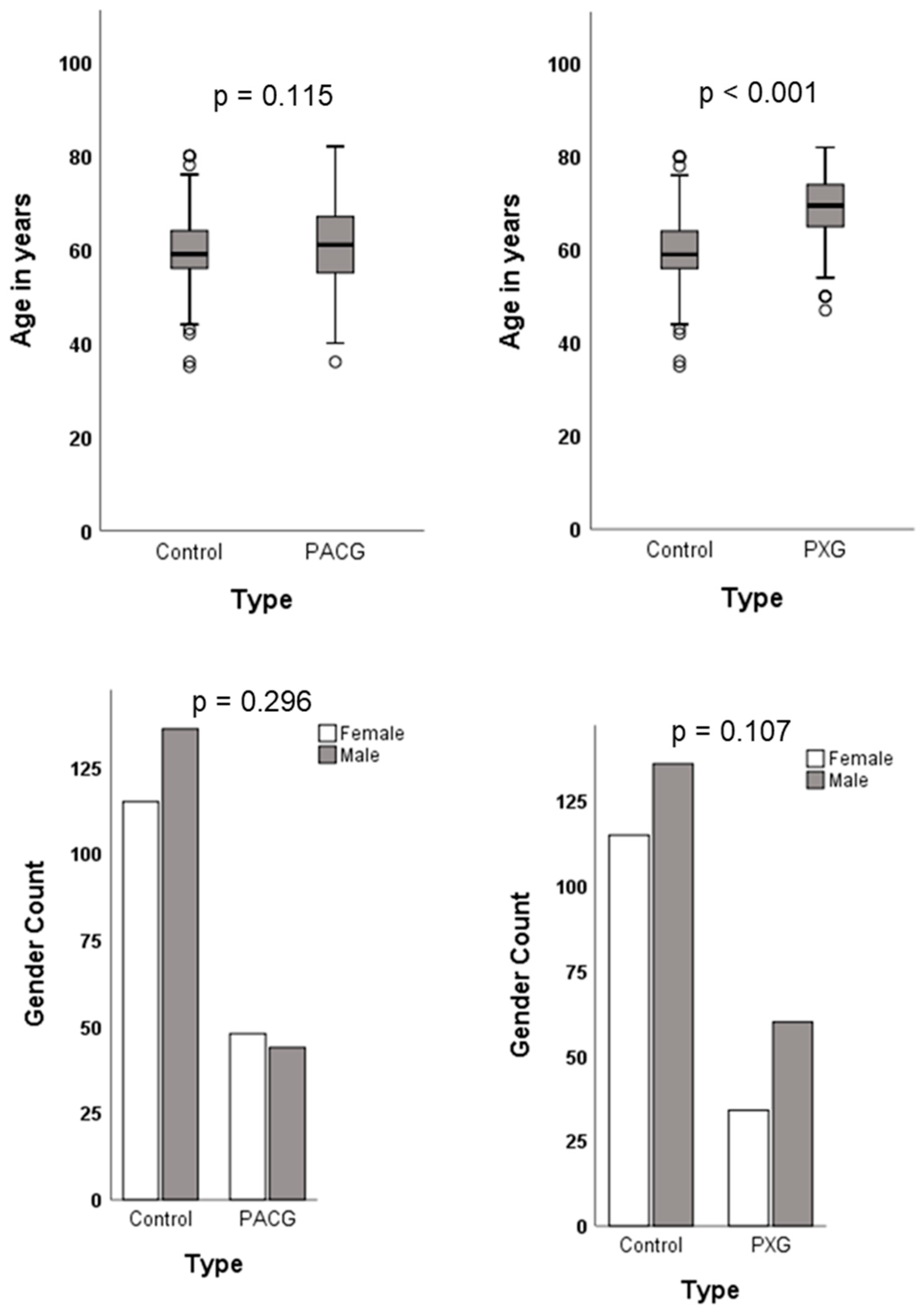
2.1. Demographic Characteristics of Study Cohort
2.2. Association Analysis of rs429358 and rs7412 in the APOE Gene
2.3. APOE Haplotype Association with PACG and PXG
2.4. Logistic Regression Analysis of Risk Factors on Glaucoma Outcome
2.5. Association between APOE Genotypes and Clinical Parameters of Glaucoma
3. Discussion
4. Materials and Methods
4.1. Study Design, Ethics Approval, and Participant Characteristics
4.2. Genotyping rs429358 and rs7412 Polymorphisms in the APOE Gene
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandez-Albarral, J.A.; Ramirez, A.I.; de Hoz, R.; Matamoros, J.A.; Salobrar-Garcia, E.; Elvira-Hurtado, L.; Lopez-Cuenca, I.; Sanchez-Puebla, L.; Salazar, J.J.; Ramirez, J.M. Glaucoma: From pathogenic mechanisms to retinal glial cell response to damage. Front. Cell Neurosci. 2024, 18, 1354569. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Dai, Y.; Chen, Y.; Yu, D.Y.; Cringle, S.J.; Chen, J.; Kong, X.; Wang, X.; Jiang, C. Primary angle closure glaucoma: What we know and what we don’t know. Prog. Retin. Eye Res. 2017, 57, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Miglior, S.; Bertuzzi, F. Exfoliative glaucoma: New evidence in the pathogenesis and treatment. Prog. Brain Res. 2015, 221, 233–241. [Google Scholar] [PubMed]
- Lee, S.S.; Mackey, D.A. Glaucoma—Risk factors and current challenges in the diagnosis of a leading cause of visual impairment. Maturitas 2022, 163, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Kondkar, A.A. Updates on genes and genetic mechanisms implicated in primary angle-closure glaucoma. Appl. Clin. Genet. 2021, 14, 89–112. [Google Scholar] [CrossRef]
- Schlotzer-Schrehardt, U. Genetics and genomics of pseudoexfoliation syndrome/glaucoma. Middle East Afr. J. Ophthalmol. 2011, 18, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, M.; Tiwari, P.; Dada, T.; Dada, R. Beyond the optic nerve: Genetics, diagnosis, and promising therapies for glaucoma. Gene 2024, 894, 147983. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.G.; March, Z.M.; Stephenson, R.A.; Narayan, P.S. Apolipoprotein e in lipid metabolism and neurodegenerative disease. Trends Endocrinol. Metab. 2023, 34, 430–445. [Google Scholar] [CrossRef]
- Anderson, D.H.; Ozaki, S.; Nealon, M.; Neitz, J.; Mullins, R.F.; Hageman, G.S.; Johnson, L.V. Local cellular sources of apolipoprotein e in the human retina and retinal pigmented epithelium: Implications for the process of drusen formation. Am. J. Ophthalmol. 2001, 131, 767–781. [Google Scholar] [CrossRef]
- Sharma, S.; Chataway, T.; Burdon, K.P.; Jonavicius, L.; Klebe, S.; Hewitt, A.W.; Mills, R.A.; Craig, J.E. Identification of loxl1 protein and apolipoprotein e as components of surgically isolated pseudoexfoliation material by direct mass spectrometry. Exp. Eye Res. 2009, 89, 479–485. [Google Scholar] [CrossRef]
- Weisgraber, K.H.; Rall, S.C., Jr.; Mahley, R.W. Human e apoprotein heterogeneity. Cysteine-arginine interchanges in the amino acid sequence of the apo-e isoforms. J. Biol. Chem. 1981, 256, 9077–9083. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.; Gregg, R.E.; Sing, C.F. Apolipoprotein e polymorphism and atherosclerosis. Arteriosclerosis 1988, 8, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Raulin, A.C.; Doss, S.V.; Trottier, Z.A.; Ikezu, T.C.; Bu, G.; Liu, C.C. Apoe in Alzheimer’s disease: Pathophysiology and therapeutic strategies. Mol. Neurodegener. 2022, 17, 72. [Google Scholar] [CrossRef]
- Fernandez-Calle, R.; Konings, S.C.; Frontinan-Rubio, J.; Garcia-Revilla, J.; Camprubi-Ferrer, L.; Svensson, M.; Martinson, I.; Boza-Serrano, A.; Venero, J.L.; Nielsen, H.M.; et al. Apoe in the bullseye of neurodegenerative diseases: Impact of the apoe genotype in Alzheimer’s disease pathology and brain diseases. Mol. Neurodegener. 2022, 17, 62. [Google Scholar] [PubMed]
- Rall, S.C., Jr.; Weisgraber, K.H.; Innerarity, T.L.; Mahley, R.W. Structural basis for receptor binding heterogeneity of apolipoprotein e from type iii hyperlipoproteinemic subjects. Proc. Natl. Acad. Sci. USA 1982, 79, 4696–4700. [Google Scholar] [CrossRef]
- Hu, M.L.; Quinn, J.; Xue, K. Interactions between apolipoprotein e metabolism and retinal inflammation in age-related macular degeneration. Life 2021, 11, 635. [Google Scholar] [CrossRef]
- Shen, L.; Hoffmann, T.J.; Melles, R.B.; Sakoda, L.C.; Kvale, M.N.; Banda, Y.; Schaefer, C.; Risch, N.; Jorgenson, E. Differences in the genetic susceptibility to age-related macular degeneration clinical subtypes. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4290–4299. [Google Scholar] [CrossRef]
- Sepulveda-Falla, D.; Sanchez, J.S.; Almeida, M.C.; Boassa, D.; Acosta-Uribe, J.; Vila-Castelar, C.; Ramirez-Gomez, L.; Baena, A.; Aguillon, D.; Villalba-Moreno, N.D.; et al. Distinct tau neuropathology and cellular profiles of an apoe3 christchurch homozygote protected against autosomal dominant Alzheimer’s dementia. Acta Neuropathol. 2022, 144, 589–601. [Google Scholar] [CrossRef]
- Liu, C.C.; Murray, M.E.; Li, X.; Zhao, N.; Wang, N.; Heckman, M.G.; Shue, F.; Martens, Y.; Li, Y.; Raulin, A.C.; et al. Apoe3-jacksonville (v236e) variant reduces self-aggregation and risk of dementia. Sci. Transl. Med. 2021, 13, eabc9375. [Google Scholar] [CrossRef]
- Le Guen, Y.; Belloy, M.E.; Grenier-Boley, B.; de Rojas, I.; Castillo-Morales, A.; Jansen, I.; Nicolas, A.; Bellenguez, C.; Dalmasso, C.; Kucukali, F.; et al. Association of rare apoe missense variants v236e and r251g with risk of alzheimer disease. JAMA Neurol. 2022, 79, 652–663. [Google Scholar] [CrossRef]
- Yi, W.; Lv, D.; Sun, Y.; Mu, J.; Lu, X. Role of apoe in glaucoma. Biochem. Biophys. Res. Commun. 2024, 694, 149414. [Google Scholar] [CrossRef]
- Margeta, M.A.; Yin, Z.; Madore, C.; Pitts, K.M.; Letcher, S.M.; Tang, J.; Jiang, S.; Gauthier, C.D.; Silveira, S.R.; Schroeder, C.M.; et al. Apolipoprotein e4 impairs the response of neurodegenerative retinal microglia and prevents neuronal loss in glaucoma. Immunity 2022, 55, 1627–1644.e7. [Google Scholar] [CrossRef]
- Omodaka, K.; Nishiguchi, K.M.; Yasuda, M.; Tanaka, Y.; Sato, K.; Nakamura, O.; Maruyama, K.; Nakazawa, T. Neuroprotective effect against axonal damage-induced retinal ganglion cell death in apolipoprotein e-deficient mice through the suppression of kainate receptor signaling. Brain Res. 2014, 1586, 203–212. [Google Scholar] [CrossRef]
- Margeta, M.A.; Letcher, S.M.; Igo, R.P., Jr.; Cooke Bailey, J.N.; Pasquale, L.R.; Haines, J.L.; Butovsky, O.; Wiggs, J.L. Association of apoe with primary open-angle glaucoma suggests a protective effect for apoe epsilon4. Investig. Ophthalmol. Vis. Sci. 2020, 61, 3. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, M.; Huang, W.; Chen, S.; Zhang, X. Lack of association of apolipoprotein e (apo e) epsilon2/epsilon3/epsilon4 polymorphisms with primary open-angle glaucoma: A meta-analysis from 1916 cases and 1756 controls. PLoS ONE 2013, 8, e72644. [Google Scholar]
- Wang, Y.; Zhou, Y.F.; Zhao, B.Y.; Gu, Z.Y.; Li, S.L. Apolipoprotein e gene epsilon4epsilon4 is associated with elevated risk of primary open angle glaucoma in asians: A meta-analysis. BMC Med. Genet. 2014, 15, 60. [Google Scholar] [CrossRef] [PubMed]
- Al-Dabbagh, N.M.; Al-Dohayan, N.; Arfin, M.; Tariq, M. Apolipoprotein e polymorphisms and primary glaucoma in saudis. Mol. Vis. 2009, 15, 912–919. [Google Scholar] [PubMed]
- Kondkar, A.A.; Sultan, T.; Azad, T.A.; Khatlani, T.; Alshehri, A.A.; Osman, E.A.; Lobo, G.P.; Almobarak, F.A.; Al-Obeidan, S.A. Common variants rs429358 and rs7412 in apoe gene are not associated with poag in a saudi cohort. Biology 2024, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Alfares, A.; Alfadhel, M.; Wani, T.; Alsahli, S.; Alluhaydan, I.; Al Mutairi, F.; Alothaim, A.; Albalwi, M.; Al Subaie, L.; Alturki, S.; et al. A multicenter clinical exome study in unselected cohorts from a consanguineous population of saudi arabia demonstrated a high diagnostic yield. Mol. Genet. Metab. 2017, 121, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Namipashaki, A.; Razaghi-Moghadam, Z.; Ansari-Pour, N. The essentiality of reporting hardy-weinberg equilibrium calculations in population-based genetic association studies. Cell J. 2015, 17, 187–192. [Google Scholar] [PubMed]
- Alagarsamy, J.; Jaeschke, A.; Hui, D.Y. Apolipoprotein e in cardiometabolic and neurological health and diseases. Int. J. Mol. Sci. 2022, 23, 9892. [Google Scholar] [CrossRef]
- Liuska, P.J.; Ramo, J.T.; Lemmela, S.; Kaarniranta, K.; Uusitalo, H.; Lahtela, E.; Daly, M.J.; Harju, M.; Palotie, A.; Turunen, J.A. Association of apoe haplotypes with common age-related ocular diseases in 412,171 individuals. Investig. Ophthalmol. Vis. Sci. 2023, 64, 33. [Google Scholar] [CrossRef] [PubMed]
- Mahley, R.W.; Rall, S.C., Jr. Apolipoprotein e: Far more than a lipid transport protein. Annu. Rev. Genom. Hum. Genet. 2000, 1, 507–537. [Google Scholar] [CrossRef]
- Corbo, R.M.; Scacchi, R. Apolipoprotein e (apoe) allele distribution in the world. Is apoe*4 a t’hrifty’ allele? Ann. Hum. Genet. 1999, 63, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.P.; Singh, M.; Mastana, S.S. Apoe distribution in world populations with new data from india and the uk. Ann. Hum. Biol. 2006, 33, 279–308. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, S.M.; Clark, A.G.; Weiss, K.M.; Nickerson, D.A.; Taylor, S.L.; Stengard, J.H.; Salomaa, V.; Vartiainen, E.; Perola, M.; Boerwinkle, E.; et al. Apolipoprotein e variation at the sequence haplotype level: Implications for the origin and maintenance of a major human polymorphism. Am. J. Hum. Genet. 2000, 67, 881–900. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Rimbach, G.; Huebbe, P. Apoe genotype: From geographic distribution to function and responsiveness to dietary factors. Proc. Nutr. Soc. 2012, 71, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Huebbe, P.; Rimbach, G. Evolution of human apolipoprotein e (apoe) isoforms: Gene structure, protein function and interaction with dietary factors. Ageing Res. Rev. 2017, 37, 146–161. [Google Scholar] [CrossRef] [PubMed]
- Mabuchi, F.; Tang, S.; Ando, D.; Yamakita, M.; Wang, J.; Kashiwagi, K.; Yamagata, Z.; Iijima, H.; Tsukahara, S. The apolipoprotein e gene polymorphism is associated with open angle glaucoma in the japanese population. Mol. Vis. 2005, 11, 609–612. [Google Scholar] [PubMed]
- Freeman, E.E.; Bastasic, J.; Grant, A.; Leung, G.; Li, G.; Buhrmann, R.; Roy-Gagnon, M.H. Inverse association of apoe epsilon4 and glaucoma modified by systemic hypertension: The canadian longitudinal study on aging. Investig. Ophthalmol. Vis. Sci. 2022, 63, 9. [Google Scholar] [CrossRef]
- Occhiutto, M.L.; de Melo, M.B.; Cabral de Vasconcellos, J.P.; Rodrigues, T.A.R.; Bajano, F.F.; Costa, F.F.; Costa, V.P. “Association of apoe gene polymorphisms with primary open angle glaucoma in brazilian patients”. Ophthalmic Genet. 2021, 42, 53–61. [Google Scholar] [CrossRef]
- Mullany, S.; Marshall, H.; Diaz-Torres, S.; Berry, E.C.; Schmidt, J.M.; Thomson, D.; Qassim, A.; To, M.S.; Dimasi, D.; Kuot, A.; et al. The apoe e4 allele is associated with faster rates of neuroretinal thinning in a prospective cohort study of suspect and early glaucoma. Ophthalmol. Sci. 2022, 2, 100159. [Google Scholar] [CrossRef] [PubMed]
- Mullany, S.; Diaz-Torres, S.; Schmidt, J.M.; Thomson, D.; Qassim, A.; Marshall, H.N.; Knight, L.S.W.; Berry, E.C.; Kolovos, A.; Dimasi, D.; et al. No strong association between the apolipoprotein e e4 allele and glaucoma: A multicohort study. Ophthalmol. Sci. 2023, 3, 100287. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.Y.; Tam, P.O.; Chiang, S.W.; Ding, N.; Chen, L.J.; Yam, G.H.; Pang, C.P.; Wang, N.L. Multiple gene polymorphisms analysis revealed a different profile of genetic polymorphisms of primary open-angle glaucoma in northern chinese. Mol. Vis. 2009, 15, 89–98. [Google Scholar] [PubMed]
- Tamura, H.; Kawakami, H.; Kanamoto, T.; Kato, T.; Yokoyama, T.; Sasaki, K.; Izumi, Y.; Matsumoto, M.; Mishima, H.K. High frequency of open-angle glaucoma in japanese patients with Alzheimer’s disease. J. Neurol. Sci. 2006, 246, 79–83. [Google Scholar] [CrossRef]
- Saglar, E.; Yucel, D.; Bozkurt, B.; Ozgul, R.K.; Irkec, M.; Ogus, A. Association of polymorphisms in apoe, p53, and p21 with primary open-angle glaucoma in turkish patients. Mol. Vis. 2009, 15, 1270–1276. [Google Scholar]
- Song, Q.; Chen, P.; Liu, Q. Role of the apoe epsilon2/epsilon3/epsilon4 polymorphism in the development of primary open-angle glaucoma: Evidence from a comprehensive meta-analysis. PLoS ONE 2013, 8, e82347. [Google Scholar] [CrossRef]
- Krumbiegel, M.; Pasutto, F.; Mardin, C.Y.; Weisschuh, N.; Paoli, D.; Gramer, E.; Weber, B.H.; Kruse, F.E.; Schlotzer-Schrehardt, U.; Reis, A. Apolipoprotein e genotypes in pseudoexfoliation syndrome and pseudoexfoliation glaucoma. J. Glaucoma 2010, 19, 561–565. [Google Scholar] [CrossRef]
- Chiras, D.; Tzika, K.; Kokotas, H.; Oliveira, S.C.; Grigoriadou, M.; Kastania, A.; Dima, K.; Stefaniotou, M.; Aspiotis, M.; Petersen, M.B.; et al. Development of novel loxl1 genotyping method and evaluation of loxl1, apoe and mthfr polymorphisms in exfoliation syndrome/glaucoma in a greek population. Mol. Vis. 2013, 19, 1006–1016. [Google Scholar]
- Yilmaz, A.; Tamer, L.; Ates, N.A.; Camdeviren, H.; Degirmenci, U. Effects of apolipoprotein e genotypes on the development of exfoliation syndrome. Exp. Eye Res. 2005, 80, 871–875. [Google Scholar] [CrossRef]
- Yaylacioglu Tuncay, F.; Aktas, Z.; Ergun, M.A.; Ergun, S.G.; Hasanreisoglu, M.; Hasanreisoglu, B. Association of polymorphisms in apoe and loxl1 with pseudoexfoliation syndrome and pseudoexfoliation glaucoma in a turkish population. Ophthalmic Genet. 2017, 38, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Ritland, J.S.; Utheim, T.P.; Utheim, O.A.; Espeseth, T.; Lydersen, S.; Semb, S.O.; Rootwelt, H.; Elsas, T. Effects of apoe and chrna4 genotypes on retinal nerve fibre layer thickness at the optic disc and on risk for developing exfoliation syndrome. Acta Ophthalmol. Scand. 2007, 85, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.Y.; Fan, B.J.; Wang, D.Y.; Tam, P.O.; Yung Tham, C.C.; Leung, D.Y.; Ping Fan, D.S.; Chiu Lam, D.S.; Pang, C.P. Association of apolipoprotein e polymorphisms with normal tension glaucoma in a chinese population. J. Glaucoma 2006, 15, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Amaratunga, A.; Abraham, C.R.; Edwards, R.B.; Sandell, J.H.; Schreiber, B.M.; Fine, R.E. Apolipoprotein e is synthesized in the retina by muller glial cells, secreted into the vitreous, and rapidly transported into the optic nerve by retinal ganglion cells. J. Biol. Chem. 1996, 271, 5628–5632. [Google Scholar] [CrossRef]
- Guillaume, D.; Bertrand, P.; Dea, D.; Davignon, J.; Poirier, J. Apolipoprotein e and low-density lipoprotein binding and internalization in primary cultures of rat astrocytes: Isoform-specific alterations. J. Neurochem. 1996, 66, 2410–2418. [Google Scholar] [CrossRef] [PubMed]
- Riddell, D.R.; Zhou, H.; Atchison, K.; Warwick, H.K.; Atkinson, P.J.; Jefferson, J.; Xu, L.; Aschmies, S.; Kirksey, Y.; Hu, Y.; et al. Impact of apolipoprotein e (apoe) polymorphism on brain apoe levels. J. Neurosci. 2008, 28, 11445–11453. [Google Scholar] [CrossRef] [PubMed]
- Bouchareychas, L.; Raffai, R.L. Apolipoprotein e and atherosclerosis: From lipoprotein metabolism to microrna control of inflammation. J. Cardiovasc. Dev. Dis. 2018, 5, 30. [Google Scholar] [CrossRef]
- Kondkar, A.A.; Sultan, T.; Azad, T.A.; Osman, E.A.; Almobarak, F.A.; Lobo, G.P.; Al-Obeidan, S.A. Evaluation of abca1 and fndc3b gene polymorphisms associated with pseudoexfoliation glaucoma and primary angle-closure glaucoma in a saudi cohort. Front. Genet. 2022, 13, 877174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| SNP ID | rs429358 | rs7412 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Type | Minor Allele | MAF | OR (95% CI) | p | HWE p | Minor Allele | MAF | OR (95% CI) | p | HWE p |
| Controls | C | 0.10 | Reference | - | 0.710 | T | 0.03 | Reference | - | 0.170 |
| PACG | C | 0.11 | 1.21 (0.70–2.1) | 0.475 | 0.086 | T | 0.05 | 1.79 (0.76–4.21) | 0.175 | 0.190 |
| PXG | C | 0.06 | 0.65 (0.33–1.24) | 0.187 | 1.000 | T | 0.04 | 1.34 (0.53–3.40) | 0.527 | 0.110 |
| APOE | Controls n (%) | PACG n (%) | OR (95% CI) | p |
|---|---|---|---|---|
| Alleles | ||||
| ε3 | 440 (87.6) | 154 (83.7) | 1.00 | - |
| ε2 | 14 (2.8) | 9 (4.9) | 1.83 (0.78–4.32) | 0.225 |
| ε4 | 48 (9.6) | 21 (11.4) | 1.25 (0.72–2.15) | 0.470 |
| Genotypes a | ||||
| ε3/ε3 | 199 (79.3) | 66 (71.7) | 1.00 | - |
| ε2/ε2 | 1 (0.4) | 1 (1.1) | 3.01 (0.18–48.89) | 0.999 |
| ε2/ε3 | 4 (1.6) | 7 (7.6) | 5.27 (1.49–18.60) | 0.009 |
| ε2/ε4 | 8 (3.2) | 0 (0) | - | 0.205 |
| ε3/ε4 | 38 (15.1) | 15 (16.3) | 1.19 (0.61–2.30) | 0.730 |
| ε4/ε4 | 1 (0.4) | 3 (3.2) | 9.04 (0.92–88.45) | 0.053 |
| ε3/ε3 vs. All | 52 (20.7) | 26 (28.2) | 1.50 (0.87–2.60) | 0.147 |
| Carrier b | ||||
| ε3/ε3 | 199 (81.9) | 66 (71.7) | 1.00 | - |
| ε*2 c | 5 (2.0) | 8 (8.7) | 4.82 (1.52–15.26) | 0.007 |
| ε*4 d | 39 (16.0) | 18 (19.5) | 1.39 (0.74–2.60) | 0.320 |
| APOE | Controls n (%) | PXG n (%) | OR (95% CI) | p |
|---|---|---|---|---|
| Alleles | ||||
| ε3 | 440 (87.6) | 169 (89.9) | 1.00 | - |
| ε2 | 14 (2.8) | 7 (3.7) | 1.30 (0.51–3.28) | 0.557 |
| ε4 | 48 (9.6) | 12 (6.4) | 0.65 (0.33–1.25) | 0.197 |
| Genotypes a | ||||
| ε3/ε3 | 199 (79.3) | 77 (81.9) | 1.00 | - |
| ε2/ε2 | 1 (0.4) | 1 (1.1) | 2.58 (0.16–41.8) | 0.999 |
| ε2/ε3 | 4 (1.6) | 4 (4.2) | 2.80 (0.63–10.60) | 0.230 |
| ε2/ε4 | 8 (3.2) | 1 (1.1) | 0.32 (0.04–2.62) | 0.452 |
| ε3/ε4 | 38 (15.1) | 11 (11.7) | 0.75 (0.36–1.53) | 0.488 |
| ε4/ε4 | 1 (0.4) | 0 (0) | - | 0.999 |
| ε3/ε3 vs. All | 52 (20.7) | 17 (18.0) | 0.85 (0.46–1.55) | 0.583 |
| Carrier b | ||||
| ε3/ε3 | 199 (81.9) | 77 (82.8) | 1.00 | - |
| ε*2 c | 5 (2.0) | 5 (5.4) | 2.58 (0.72–9.17) | 0.156 |
| ε*4 d | 39 (16.0) | 11 (11.8) | 0.72 (0.35–1.49) | 0.489 |
| Group Variables | B a | SE | Wald | OR (95% CI) | p |
|---|---|---|---|---|---|
| PACG | |||||
| Age | 0.023 | 0.017 | 1.812 | 1.02 (0.99–1.05) | 0.178 |
| Sex | −0.285 | 0.250 | 1.292 | 0.75 (0.46–1.23) | 0.256 |
| APOE genotypes | 7.487 | 0.024 | |||
| ε2-carriers b | 1.556 | 0.590 | 6.953 | 4.74 (1.49–15.06) | 0.008 |
| ε4-carriers c | 0.324 | 0.320 | 1.023 | 1.38 (0.74–2.59) | 0.312 |
| PXG | |||||
| Age | 0.163 | 0.021 | 61.389 | 1.18 (1.130–1.22) | 0.000 |
| Sex | 0.202 | 0.294 | 0.475 | 1.22 (0.69–2.17) | 0.490 |
| APOE genotypes | 1.342 | 0.511 | |||
| ε2-carriers b | 0.388 | 0.778 | 0.249 | 1.47 (0.32–6.77) | 0.618 |
| ε4-carriers c | −0.438 | 0.436 | 1.010 | 0.64 (0.27–1.51) | 0.315 |
| APOE Alleles | ε3 | ε2 | ε4 |
|---|---|---|---|
| Haplotype | rs429358-T rs7412-C | rs429358-T rs7412-T | rs429358-C rs7412-C |
| Residue combination | 112-Cys 158-Arg | 112-Cys 158-Cys | 112-Arg 158-Arg |
| Ethnicity | Allele frequency a | ||
| Europeans | 0.640–0.900 | 0.044–0.120 | 0.052–0.310 |
| Asians | 0.620–0.870 | 0.020–0.140 | 0.071–0.240 |
| Africans | 0.536–0.850 | 0.031–0.116 | 0.085–0.407 |
| Native Americans | 0.720–0.911 | 0.0–0.014 | 0.089–0.280 |
| Oceanians | 0.486–0.740 | 0.0–0.145 | 0.260–0.368 |
| Our study (Saudi Arabians) b | 0.876 | 0.028 | 0.096 |
| Primer Type | APOE Primer Sequences (5′–3′) | Thermal Cycling Conditions |
|---|---|---|
| Forward | a GACCATGAAGGAGTTGAAGGCCTAC | Initial denaturation—95 °C for 15 min Cycling—95 °C—1 min, 59 °C—30 s, 72 °C—1 min for 35 cycles Final extension—72 °C—10 min |
| Reverse | b GATGGCGCTGAGGCCGCGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondkar, A.A.; Azad, T.A.; Sultan, T.; Khatlani, T.; Alshehri, A.A.; Radhakrishnan, R.; Lobo, G.P.; Alsirhy, E.; Almobarak, F.A.; Osman, E.A.; et al. APOE ε2-Carriers Are Associated with an Increased Risk of Primary Angle-Closure Glaucoma in Patients of Saudi Origin. Int. J. Mol. Sci. 2024, 25, 4571. https://doi.org/10.3390/ijms25084571
Kondkar AA, Azad TA, Sultan T, Khatlani T, Alshehri AA, Radhakrishnan R, Lobo GP, Alsirhy E, Almobarak FA, Osman EA, et al. APOE ε2-Carriers Are Associated with an Increased Risk of Primary Angle-Closure Glaucoma in Patients of Saudi Origin. International Journal of Molecular Sciences. 2024; 25(8):4571. https://doi.org/10.3390/ijms25084571
Chicago/Turabian StyleKondkar, Altaf A., Taif A. Azad, Tahira Sultan, Tanvir Khatlani, Abdulaziz A. Alshehri, Rakesh Radhakrishnan, Glenn P. Lobo, Ehab Alsirhy, Faisal A. Almobarak, Essam A. Osman, and et al. 2024. "APOE ε2-Carriers Are Associated with an Increased Risk of Primary Angle-Closure Glaucoma in Patients of Saudi Origin" International Journal of Molecular Sciences 25, no. 8: 4571. https://doi.org/10.3390/ijms25084571