
Salivary Protein Cyclin-Dependent Kinase-like from Grain Aphid Sitobion avenae Suppresses Wheat Defense Response and Enhances Aphid Adaptation
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
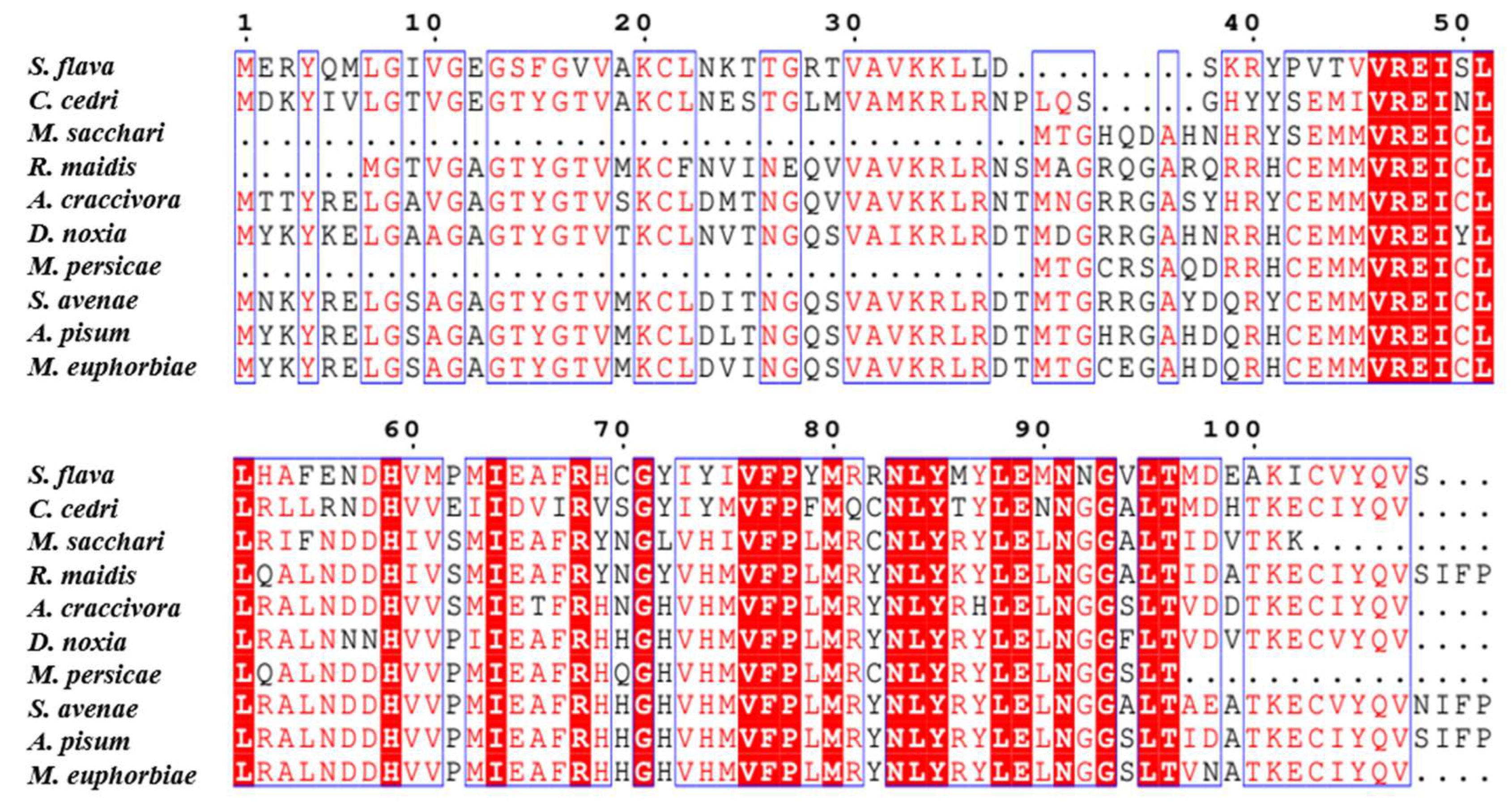
2.1. Sequence Analysis of Candidate Salivary Effector SaCDK
2.2. Spatio-Temporal Expression Profile Analysis of SaCDK
2.3. Transient Expression of SaCDK in N. benthamiana Suppresses Cell Death
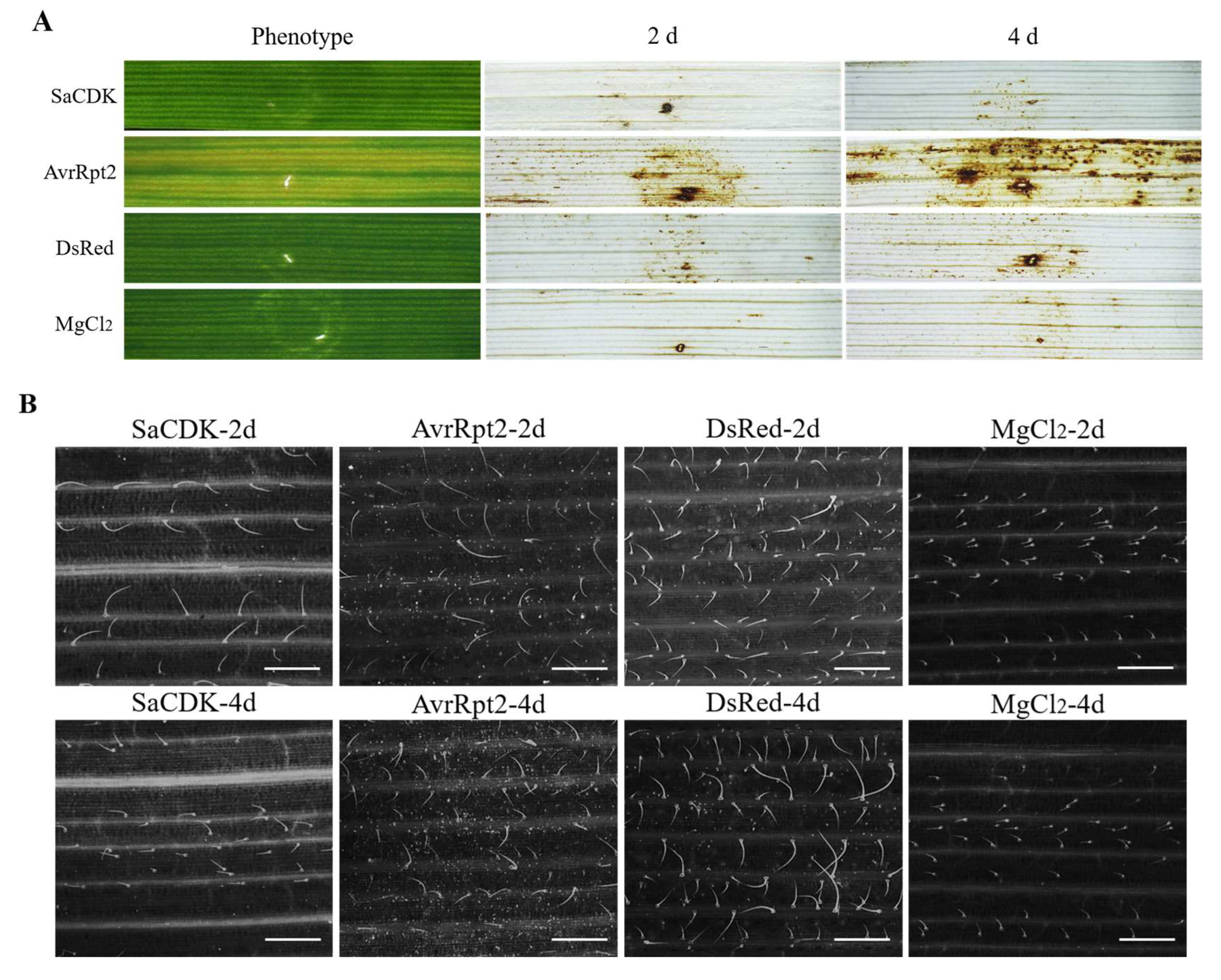
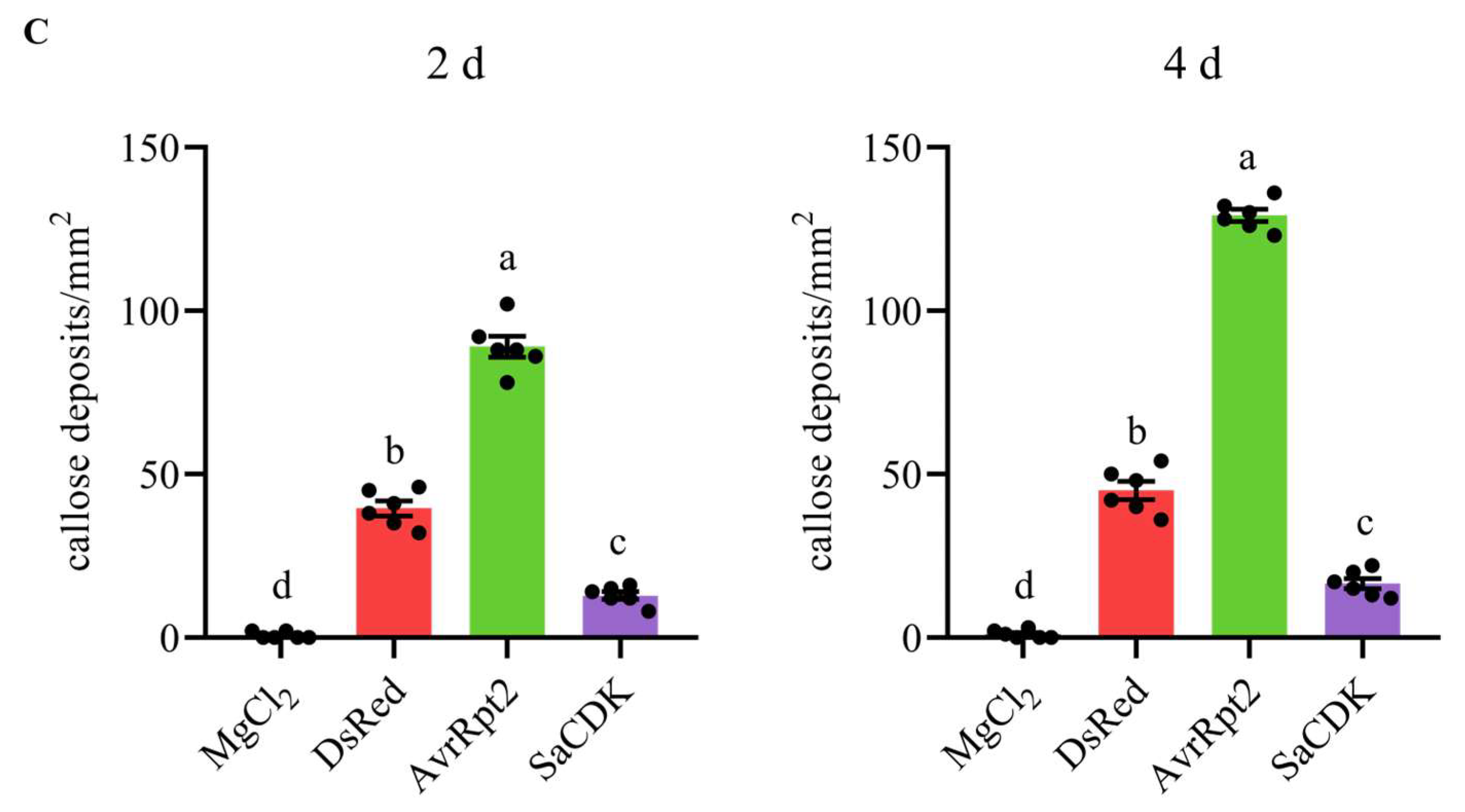
2.4. Delivery of SaCDK Inhibited Callose Deposition in Wheat Leaves
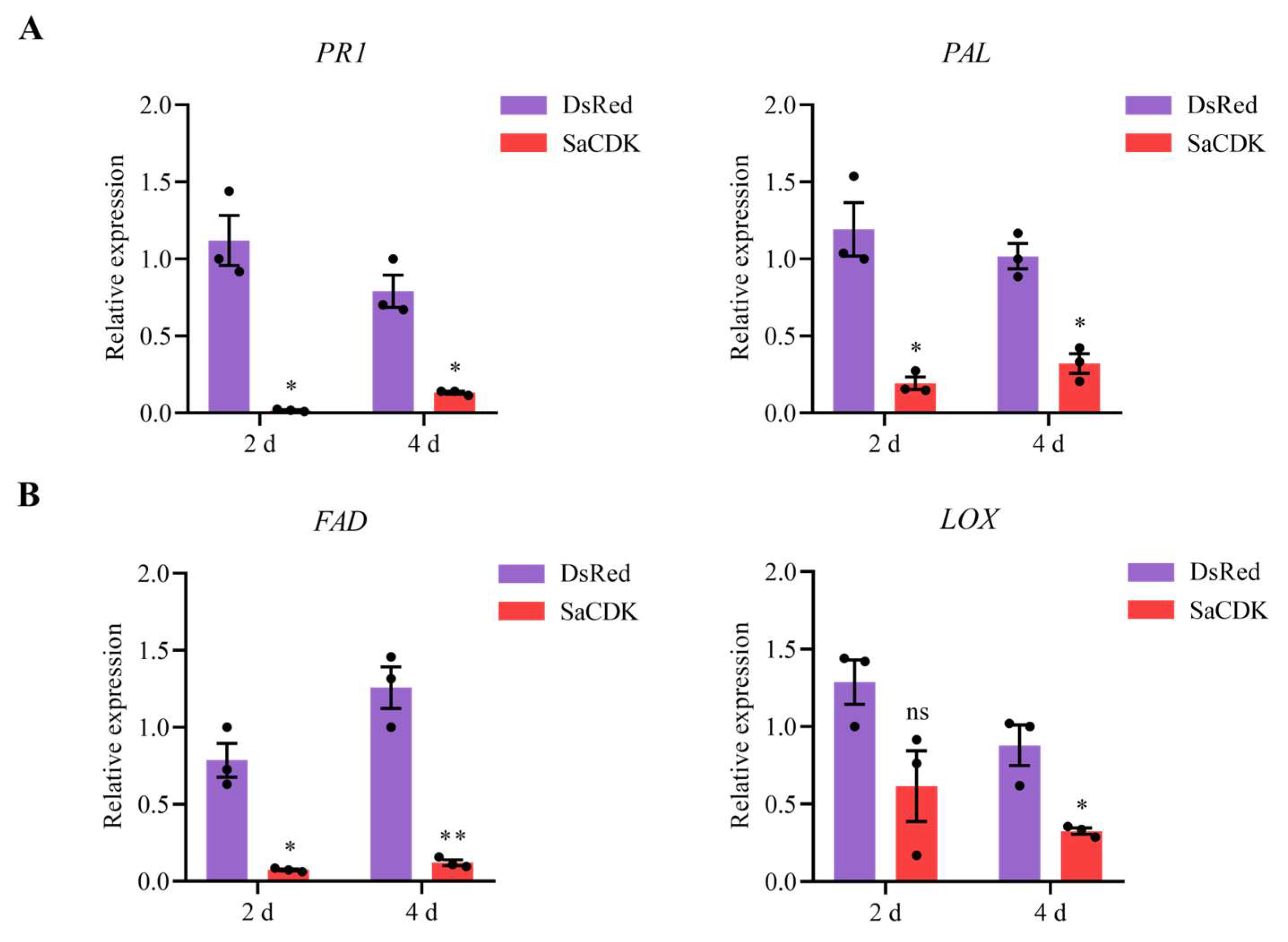
2.5. SaCDK Inhibited the SA and JA Defense Signaling Pathways
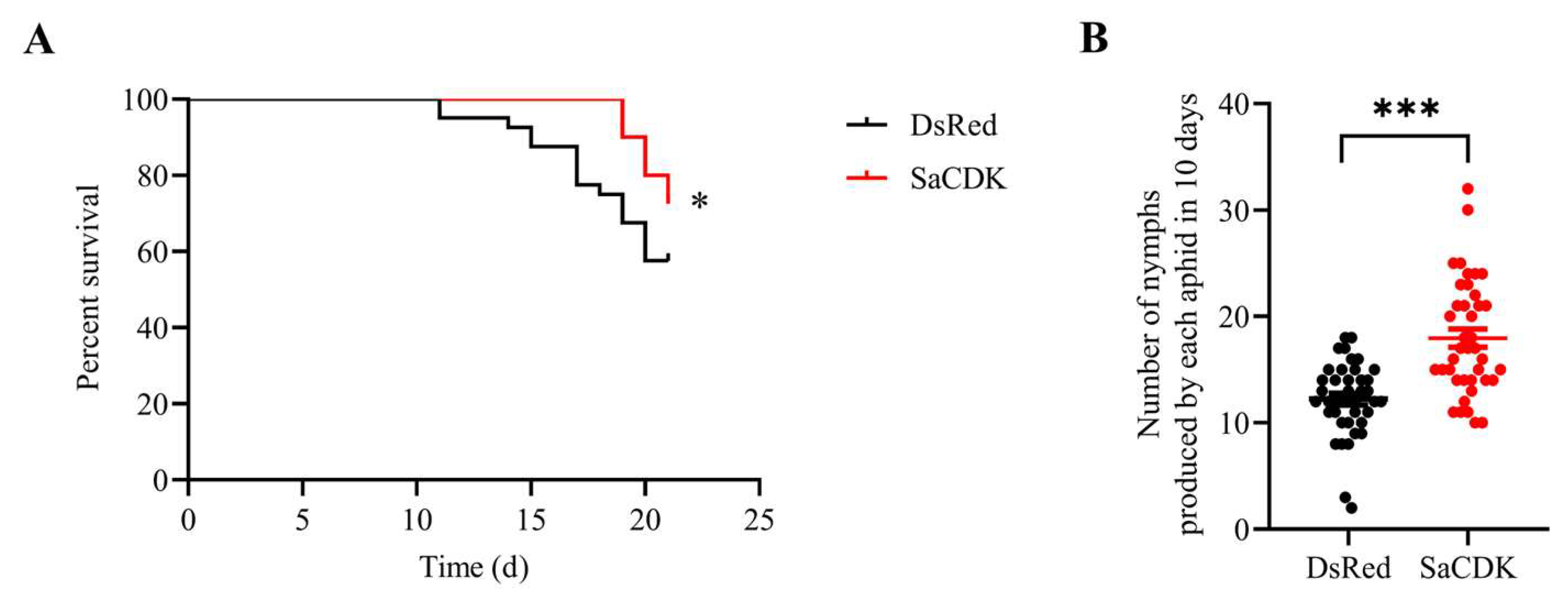
2.6. SaCDK Enhanced the Survival and Fecundity of S. avenae
3. Discussion
4. Materials and Methods
4.1. Aphids and Plants
4.2. Sequence Analysis
4.3. Transient Expression of SaCDK in N. benthamiana
4.4. Delivery of SaCDK into Wheat via the Bacterial Type III Secretion System
4.5. Hydrogen Peroxide Accumulation and Callose Deposition in Wheat Leaves
4.6. RT-qPCR
4.7. Aphid Bioassay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Overmyer, K.; Vuorinen, K.; Brosché, M. Interaction points in plant stress signaling pathways. Physiol. Plant. 2018, 162, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Mignolet-Spruyt, L.; Xu, E.J.; Idänheimo, N.; Hoeberichts, F.A.; Mühlenbock, P.; Brosché, M.; Breusegem, F.V.; Kangasjärvi, J. Spreading the news: Subcellular and organellar reactive oxygen species production and signalling. J. Exp. Bot. 2016, 67, 3831–3844. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, A.M.; Argueso, C.T. No hormone to rule them all: Interactions of plant hormones during the responses of plants to pathogens. Semin. Cell Dev. Biol. 2016, 56, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Böhm, H.; Albert, I.; Fan, L.; Reinhard, A.; Nürnberger, T. Immune receptor complexes at the plant cell surface. Curr. Opin. Plant Biol. 2014, 20, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Pitzschke, A.; Schikora, A.; Hirt, H. MAPK cascade signalling networks in plant defence. Curr. Opin. Plant Biol. 2009, 12, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ji, D.; Chen, T.; Li, B.; Zhang, Z.; Qin, G.; Tian, S. Production, signaling, and scavenging mechanisms of reactive oxygen species in fruit-pathogen interactions. Int. J. Mol. Sci. 2019, 20, 2994. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Resjö, S.; Zahid, M.A.; Burra, D.D.; Lenman, M.; Levander, F.; Andreasson, E. Proteomics of PTI and two ETI immune reactions in potato leaves. Int. J. Mol. Sci. 2019, 20, 4726. [Google Scholar] [CrossRef]
- Steinbrenner, A.D.; Saldivar, E.; Hodges, N.; Guayazán-Palacios, N.; Chaparro, A.F.; Schmelz, E.A. Signatures of plant defense response specificity mediated by herbivore-associated molecular patterns in legumes. Plant J. 2022, 110, 1255–1270. [Google Scholar] [CrossRef]
- Zogli, P.; Pingault, L.; Grover, S.; Louis, J. Ento (o) mics: The intersection of ‘omic’approaches to decipher plant defense against sap-sucking insect pests. Curr. Opin. Plant Biol. 2020, 56, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, X.X.; Zhang, J.; Liu, B.F.; Zhao, Y.; Wang, H.Y.; Wang, Z.Z.; Guo, J.P.; Rao, W.W.; Jing, S.L.; Guan, W.; et al. A mucin-like protein of planthopper is required for feeding and induces immunity response in plants. Plant Physiol. 2018, 176, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Miles, P.W. Aphid saliva. Biol. Rev. 1999, 74, 41–85. [Google Scholar] [CrossRef]
- Cherqui, A.; Tjallingii, W.F. Salivary proteins of aphids, a pilot study on identification, separation and immunolocalisation. J. Insect Physiol. 2000, 46, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Tjallingii, W.F.; Thönnessen, A.; van Bel, A.J.E. Molecular sabotage of plant defense by aphid saliva. Proc. Natl. Acad. Sci. USA 2007, 104, 10536–10541. [Google Scholar] [CrossRef] [PubMed]
- Mutti, N.S.; Louis, J.; Pappan, L.K.; Pappan, K.; Begum, K.; Chen, M.; Park, Y.; Dittmer, N.; Marshall, J.; Reese, J.C.; et al. A protein from the salivary glands of the pea aphid, Acyrthosiphon pisum, is essential in feeding on a host plant. Proc. Natl. Acad. Sci. Belarus-Agrar. Ser. 2008, 105, 9965–9969. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.I.B.; Prince, D.; Pitino, M.; Maffei, M.E.; Win, J.; Hogenhout, S.A. A functional genomics approach identifies candidate effectors from the aphid species Myzus persicae (green peach aphid). PLoS Genet. 2010, 6, e1001216. [Google Scholar] [CrossRef]
- Pitino, M.; Hogenhout, S.A. Aphid protein effectors promote aphid colonization in a plant species-specific manner. Mol. Plant-Microbe Interact. 2013, 26, 130–139. [Google Scholar] [CrossRef]
- Elzinga, D.A.; De Vos, M.; Jander, G. Suppression of plant defenses by a Myzus persicae (green peach aphid) salivary effector protein. Mol. Plant-Microbe Interact. 2014, 27, 747–756. [Google Scholar] [CrossRef]
- Atamian, H.S.; Chaudhary, R.; Cin, V.D.; Bao, E.; Girke, T.; Kaloshian, I. In planta expression or delivery of potato aphid Macrosiphum euphorbiae effectors Me10 and Me23 enhances aphid fecundity. Mol. Plant-Microbe Interact. 2013, 26, 67–74. [Google Scholar] [CrossRef]
- Escudero-Martinez, C.; Rodriguez, P.A.; Liu, S.; Santos, P.A.; Stephens, J.; Bos, J.I. An aphid effector promotes barley susceptibility through suppression of defence gene expression. J. Exp. Bot. 2020, 71, 2796–2807. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Cui, J.R.; Xia, X.; Chen, J.; Ye, Y.X.; Zhang, C.X.; Hong, X.Y. Salivary DNase II from Laodelphax striatellus acts as an effector that suppresses plant defence. New Phytol. 2019, 224, 860–874. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Wang, Y.Z.; Li, L.L.; Lu, H.B.; Lu, J.B.; Wang, X.; Ye, Z.X.; Zhang, Z.L.; He, Y.J.; Lu, G.; et al. Planthopper salivary sheath protein LsSP1 contributes to manipulation of rice plant defenses. Nat. Commun. 2023, 14, 737. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Peng, Z.K.; Tong, H.; Xie, W.; Wang, S.L.; Wu, Q.J.; Zhang, J.M.; Li, C.R.; Zhang, Y. A salivary ferritin in the whitefly suppresses plant defenses and facilitates host exploitation. J. Exp. Bot. 2019, 70, 3343–3355. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Ye, Y.X.; Lu, J.B.; Wang, X.; Lu, H.B.; Zhang, Z.L.; Ye, Z.X.; Lu, Y.W.; Sun, Z.T.; Chen, J.P.; et al. Horizontally transferred salivary protein promotes insect feeding by suppressing ferredoxin-mediated plant defenses. Mol. Biol. Evol. 2023, 40, msad221. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Gao, X. Multiple mechanisms responsible for differential susceptibilities of Sitobion avenae (Fabricius) and Rhopalosiphum padi (Linnaeus) to pirimicarb. Bull. Entomol. Res. 2009, 99, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Thackray, D.J.; Diggle, A.J.; Jones, R.A.C. BYDV predictor: A simulation model to predict aphid arrival, epidemics of Barley yellow dwarf virus and yield losses in wheat crops in a Mediterranean-type environment. Plant Pathol. 2009, 58, 186–202. [Google Scholar] [CrossRef]
- Liu, X.F.; Hu, X.S.; Keller, M.A.; Zhao, H.Y.; Wu, Y.F.; Liu, T.X. Tripartite interactions of barley yellow dwarf virus, Sitobion avenae and wheat varieties. PLoS ONE. 2014, 9, e106639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, J.; Francis, F.; Chen, J.L. Watery saliva secreted by the grain aphid Sitobion avenae stimulates aphid resistance in wheat. J. Agric. Food Chem. 2017, 65, 8798–8805. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, J.; Sun, J.R.; Francis, F.; Chen, J.L. Transcriptome analysis of the salivary glands of the grain aphid, Sitobion avenae. Sci. Rep. 2017, 7, 15911. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, Y.; Liu, X.B.; Francis, F.; Fan, J.; Liu, H.; Wang, Q.; Sun, Y.; Zhang, Y.M.; Chen, J.L. SmCSP4 from aphid saliva stimulates salicylic acid-mediated defence responses in wheat by interacting with transcription factor TaWKRY76. Plant Biotechnol. J. 2023, 21, 2389–2407. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, X.B.; Wang, Q.; Liu, H.; Cheng, Y.M.; Li, H.M.; Zhang, Y.; Chen, J.L. Two salivary proteins Sm10 and SmC002 from grain aphid Sitobion miscanthi modulate wheat defense and enhance aphid performance. Front. Plant Sci. 2023, 14, 1104275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.B.; Fu, Y.; Crespo-Herrera, L.; Liu, H.; Wang, Q.; Zhang, Y.M.; Chen, J.L. Salivary effector Sm9723 of grain aphid Sitobion miscanthi suppresses plant defense and is essential for aphid survival on wheat. Int. J. Mol. Sci. 2022, 23, 6909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.B.; Francis, F.; Xie, H.C.; Fan, J.; Wang, Q.; Liu, H.; Sun, Y.; Chen, J.L. The salivary effector protein Sg2204 in the greenbug Schizaphis graminum suppresses wheat defence and is essential for enabling aphid feeding on host plants. Plant Biotechnol. J. 2022, 20, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Elzinga, D.A.; Jander, G. The role of protein effectors in plant-aphid interactions. Curr. Opin. Plant Biol. 2013, 16, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.D.; Michel, A. Mechanisms of aphid adaptation to host plant resistance. Curr. Opin. Insect Sci. 2018, 26, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Shimotohno, A.; Umeda-Hara, C.; Bisova, K.; Uchimiya, H.; Umeda, M. The plant-specific kinase CDKF; 1 is involved in activating phosphorylation of cyclin-dependent kinase-activating kinases in Arabidopsis. Plant Cell. 2004, 16, 2954–2966. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.F.; Fan, B.F.; Scholz, J.; Chen, Z.X. Roles of Arabidopsis cyclin-dependent kinase C complexes in cauliflower mosaic virus infection, plant growth, and development. Plant Cell. 2007, 19, 1388–1402. [Google Scholar] [CrossRef]
- Zhu, Y.F.; Schluttenhoffer, C.M.; Wang, P.C.; Fu, F.Y.; Thimmapuram, J.; Zhu, J.K.; Yeol Lee, S.; Yun, D.J.; Mengiste, T. CYCLIN-DEPENDENT KINASE8 differentially regulates plant immunity to fungal pathogens through kinase-dependent and-independent functions in Arabidopsis. Plant Cell. 2014, 26, 4149–4170. [Google Scholar] [CrossRef]
- Li, Q.; Fu, Y.; Liu, X.B.; Sun, J.X.; Hou, M.L.; Zhang, Y.; Chen, J.L. Activation of wheat defense response by Buchnera aphidicola-derived small chaperone protein GroES in wheat aphid saliva. J. Agric. Food Chem. 2022, 70, 1058–1067. [Google Scholar] [CrossRef]
- Chaudhary, R.; Atamian, H.S.; Shen, Z.X.; Briggs, S.P.; Kaloshian, I. GroEL from the endosymbiont Buchnera aphidicola betrays the aphid by triggering plant defense. Proc. Natl. Acad. Sci. USA 2014, 111, 8919–8924. [Google Scholar] [CrossRef]
- Boumela, I.; Guillemin, Y.; Guérin, J.F.; Aouacheria, A. The Bcl-2 family pathway in gametes and preimplantation embryos. Gynécologie Obs. Fertil. 2009, 37, 720–732. [Google Scholar] [CrossRef]
- Kamoun, S.; Klucher, K.M.; Coffey, M.D.; Tyler, B.M. A gene encoding a host-specific elicitor protein of Phytophthora parasitica. Mol. Plant-Microbe Interact. 1993, 6, 573. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, Q.; Zhang, Y.; Chen, J.L. Cloning and functional analysis of salivary protein gene Sm13498 of the grain aphid, Sitobion miscanthi (Hemiptera: Aphididae). Acta Ent. Sin. 2021, 64, 1009–1019. [Google Scholar]
- Duan, C.X.; Wang, X.M.; Zhu, Z.D.; Zhang, Z.W.; Jin, D.S. Review of studies on wheat resistance to Sitobion avenae (Fabricius) in China. J. Plant. Genet. Resour. 2003, 4, 175–178. [Google Scholar]
- Segonzac, C.; Feike, D.; Gimenez-Ibanez, S.; Hann, D.R.; Zipfel, C.; Rathjen, J.P. Hierarchy and roles of pathogen-associated molecular pattern-induced responses in Nicotiana benthamiana. Plant Physiol. 2011, 156, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.L.; Wu, K.; Yao, J.N.; Li, S.M.; Wang, X.J.; Huang, L.L.; Kang, Z.S. PSTha5a23, a candidate effector from the obligate biotrophic pathogen Puccinia striiformis f. sp. tritici, is involved in plant defense suppression and rust pathogenicity. Environ. Microbiol. 2017, 19, 1717–1729. [Google Scholar] [PubMed]
- Zhao, M.X.; Wang, J.F.; Ji, S.; Chen, Z.J.; Xu, J.H.; Tang, C.L.; Chen, S.T.; Kang, Z.S.; Wang, X.J. Candidate effector Pst_8713 impairs the plant immunity and contributes to virulence of Puccinia striiformis f. sp. tritici. Front. Plant Sci. 2018, 9, 1294. [Google Scholar] [CrossRef]
- Bai, X.X.; Zhan, G.M.; Tian, S.X.; Peng, H.; Cui, X.Y.; Islam, M.A.; Goher, F.; Ma, Y.Z.; Kang, Z.S.; Xu, Z.S.; et al. Transcription factor BZR2 activates chitinase Cht20.2 transcription to confer resistance to wheat stripe rust. Plant Physiol. 2021, 187, 2749–2762. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.T.; Hulbert, S. Prospects for functional analysis of effectors from cereal rust fungi. Euphytica 2011, 179, 57–67. [Google Scholar] [CrossRef]
- Geng, X.Q.; Shen, M.Z.; Kim, J.H.; Mackey, D. The Pseudomonas syringae type III effectors AvrRpm1 and AvrRpt2 promote virulence dependent on the F-box protein COI1. Plant Cell Reports 2016, 35, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.T.S.; Kunkel, B.N. The Pseudomonas syringae avrRpt2 Gene Contributes to Virulence on Tomato. Mol. Plant-Microbe Interact. 2005, 18, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Agnew, J.L.; Cohen, J.D.; He, P.; Shan, L.B.; Sheen, J.; Kunkel, B.N. Pseudomonas syringae type III effector AvrRpt2 alters Arabidopsis thaliana auxin physiology. Proc. Natl. Acad. Sci. USA 2007, 104, 20131–20136. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; van Bel, A.J.E. Physical and chemical interactions between aphids and plants. J. Exp. Bot. 2006, 57, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.P.; Atta, U.R.; Wang, R.F.; Liu, K.X.; Ma, X.; Weng, Q.B. Defense-related hormone signaling coordinately controls the role of melatonin during Arabidopsis thaliana-Pseudomonas syringae interaction. Eur. J. Plant Pathol. 2021, 160, 707–716. [Google Scholar] [CrossRef]
- Li, Z.; Ahammed, G.J. Salicylic acid and jasmonic acid in elevated CO2-induced plant defense response to pathogens. J. Plant Physiol. 2023, 286, 154019. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Z.; Chen, H.H.; Wang, Y.W.; Zhu, J.C.; Zhang, X.L.; Sun, J.; Liu, F.; Zhao, Y.Y. Function analysis of GhWRKY53 regulating cotton resistance to verticillium wilt by JA and SA signaling pathways. Front. Plant Sci. 2023, 14, 1203695. [Google Scholar] [CrossRef] [PubMed]
- Züst, T.; Agrawal, A.A. Mechanisms and evolution of plant resistance to aphids. Nat. Plants 2016, 2, 15206. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Shrestha, K.; Huang, Y.H. Revealing Differential Expression of Phytohormones in Sorghum in Response to Aphid Attack Using the Metabolomics Approach. Int. J. Mol. Sci. 2022, 23, 13782. [Google Scholar] [CrossRef]
- Feng, J.L.; Zhang, J.; Yang, J.; Zou, L.P.; Fang, T.T.; Xu, H.L.; Cai, Q.N. Exogenous salicylic acid improves resistance of aphid-susceptible wheat to the grain aphid, Sitobion avenae (F.) (Hemiptera: Aphididae). Bull. Entomol. Res. 2021, 111, 544–552. [Google Scholar] [CrossRef]
- Cooper, W.C.; Jia, L.; Goggin, F.L. Acquired and R-gene-mediated resistance against the potato aphid in tomato. J. Chem. Ecol. 2004, 30, 2527–2542. [Google Scholar] [CrossRef] [PubMed]
- Javed, K.; Wang, Y.; Javed, H.; Wang, C.; Liu, C.; Huang, Y.Q. Tomato Aphid (Aphis gossypii) Secreted Saliva Can Enhance Aphid Resistance by Upregulating Signaling Molecules in Tomato (Solanum lycopersicum). Int. J. Mol. Sci. 2023, 24, 12768. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Tang, C.L.; Wang, X.D.; Sun, S.T.; Zhao, J.R.; Kang, Z.S.; Wang, X.J. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef] [PubMed]
- Hood, M.E.; Shew, H.D. Applications of KOH-aniline blue fluorescence in the study of plant-fungal interactions. Phytopathology 1996, 86, 704–708. [Google Scholar] [CrossRef]
- Wang, C.F.; Huang, L.L.; Buchenauer, H.; Han, Q.M.; Zhang, H.C.; Kang, Z.S. Histochemical studies on the accumulation of reactive oxygen species (O2− and H2O2) in the incompatible and compatible interaction of wheat—Puccinia striiformis f. sp. tritici. Physiol. Mol. Plant Pathol. 2007, 71, 230–239. [Google Scholar] [CrossRef]
- Yang, C.X.; Pan, H.P.; Liu, Y.; Zhou, X.G. Selection of reference genes for expression analysis using quantitative real-time PCR in the pea aphid, Acyrthosiphon pisum (Harris) (Hemiptera, Aphidiae). PLoS ONE 2014, 9, e110454. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, X.; Sun, Y.; Liu, Y.; Zhang, Y.; Ding, T.; Chen, J. Salivary Protein Cyclin-Dependent Kinase-like from Grain Aphid Sitobion avenae Suppresses Wheat Defense Response and Enhances Aphid Adaptation. Int. J. Mol. Sci. 2024, 25, 4579. https://doi.org/10.3390/ijms25094579
Zhang Y, Liu X, Sun Y, Liu Y, Zhang Y, Ding T, Chen J. Salivary Protein Cyclin-Dependent Kinase-like from Grain Aphid Sitobion avenae Suppresses Wheat Defense Response and Enhances Aphid Adaptation. International Journal of Molecular Sciences. 2024; 25(9):4579. https://doi.org/10.3390/ijms25094579
Chicago/Turabian StyleZhang, Yumeng, Xiaobei Liu, Yu Sun, Yong Liu, Yong Zhang, Tianbo Ding, and Julian Chen. 2024. "Salivary Protein Cyclin-Dependent Kinase-like from Grain Aphid Sitobion avenae Suppresses Wheat Defense Response and Enhances Aphid Adaptation" International Journal of Molecular Sciences 25, no. 9: 4579. https://doi.org/10.3390/ijms25094579