

ZmNAC17 Regulates Mesocotyl Elongation by Mediating Auxin and ROS Biosynthetic Pathways in Maize
 and
and
Abstract
:1. Introduction
2. Results
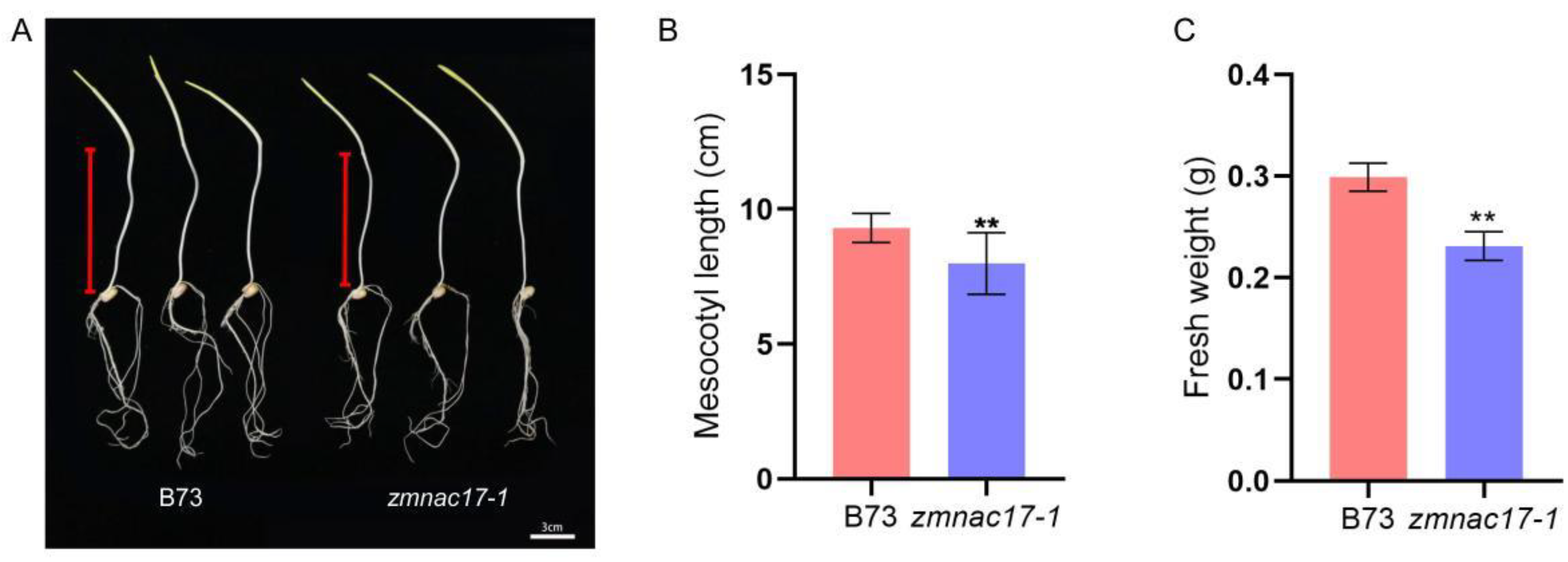
2.1. Phenotype of Maize zmnac17-1 and ZmNAC17-2 Mutants
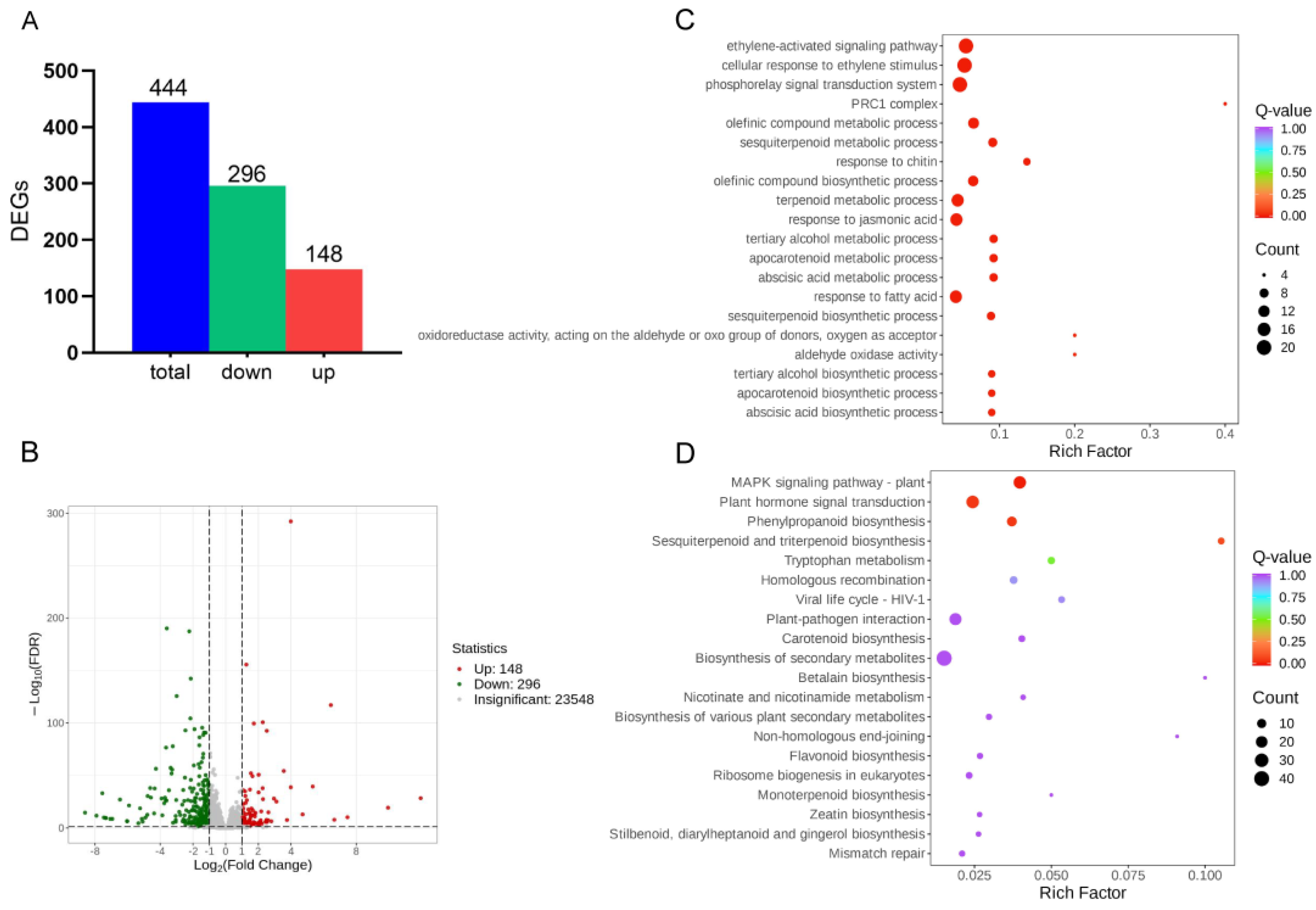
2.2. GO and KEGG Enrichment Analyses
2.3. Auxin Regulates Mesocotyl Length in zmnac17-1 Mutant
2.4. ROS Level Change in zmnac17-1 Mutant during Mesocotyl Elongation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Phenotypic Analysis
4.2. Determination of ROS and Antioxidant Metabolites
4.3. Determination of Endogenous IAA
4.4. RNA Sequencing and Data Analysis
4.5. qRT-PCR Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Ma, P.; Zhao, Z.; Zhao, G.; Tian, B.; Wang, J.; Wang, G. Mapping QTL controlling maize deep-seeding tolerance-related traits and confirmation of a major QTL for mesocotyl length. Theor. Appl. Genet. 2012, 124, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Hochholdinger, F.; Yu, P.; Marcon, C. Genetic control of root system development in maize. Trends Plant Sci. 2018, 23, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Maniou, F.; Chorianopoulou, S.N.; Bouranis, D.L. New insights into trophic aerenchyma formation strategy in maize (Zea mays L.) organs during sulfate deprivation. Front. Plant Sci. 2014, 5, 581. [Google Scholar] [CrossRef] [PubMed]
- Sauter, A.; Hartung, W. The contribution of internode and mesocotyl tissues to root-to-shoot signalling of abscisic acid. J. Exp. Bot. 2002, 53, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Q.; Wang, M.; Xiao, F.; Li, K.; Yang, R.; Sun, M.; Zhang, H.; Guo, J.; Chen, J.; et al. ZmCOP1 regulates maize mesocotyl length and plant height through the phytohormone pathways. Life 2023, 13, 1522. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Jiang, H.; Zhao, G.; Ren, J. Gene cloning of ZmMYB59 transcription factor in maize and its expression during seed germination in response to deep-sowing and exogenous hormones. Plant Breed 2017, 136, 834–844. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, L.; Yu, J. Maize WRKY28 interacts with the DELLA protein D8 to affect skotomorphogenesis and participates in the regulation of shade avoidance and plant architecture. J. Exp. Bot. 2023, 74, 3122–3141. [Google Scholar] [CrossRef] [PubMed]
- Friml, J. Subcellular trafficking of PIN auxin efflux carriers in auxin transport. Eur. J. Cell Biol. 2010, 89, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Jiang, C.; Chen, X.; Zhang, T.; Ding, L.; Song, W.; Luo, H.; Lai, J.; Chen, H.; Liu, R.; et al. Maize LAZY1 mediates shoot gravitropism and inflorescence development through regulating auxin transport, auxin signaling, and light response. Plant Physiol. 2013, 163, 1306–1322. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, Y.; Huang, G.; Zhu, N.; Li, Z.; Zhang, M.; Duan, L. Coronatine inhibits mesocotyl elongation by promoting ethylene production in etiolated maize seedlings. Plant Growth Regul. 2020, 90, 51–61. [Google Scholar] [CrossRef]
- Borucka, J.; Fellner, M. Auxin binding proteins ABP1 and ABP4 are involved in the light- and auxin-induced down-regulation of phytochrome gene PHYB in maize (Zea mays L.) mesocotyl. Plant Growth Regul. 2012, 68, 503–509. [Google Scholar] [CrossRef]
- Van Breusegem, F.; Bailey-Serres, J.; Mittler, R. Unraveling the tapestry of networks involving reactive oxygen species in plants. Plant Physiol. 2008, 147, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Tarkowski, Ł.P.; Signorelli, S.; Considine, M.J.; Montrichard, F. Integration of reactive oxygen species and nutrient signalling to shape root system architecture. Plant Cell Environ. 2023, 46, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Niu, Y.; Bai, X.; Mao, T. Transcriptomic and metabolic profiling reveals a lignin metabolism network involved in mesocotyl elongation during maize seed germination. Plants 2022, 11, 1034. [Google Scholar] [CrossRef] [PubMed]
- Fehér, A.; Otvös, K.; Pasternak, T.P.; Szandtner, A.P. The involvement of reactive oxygen species (ROS) in the cell cycle activation (G(0)-to-G(1) transition) of plant cells. Plant Signal. Behav. 2008, 3, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Mangano, S.; Denita-Juarez, S.P.; Choi, H.S.; Marzol, E.; Hwang, Y.; Ranocha, P.; Velasquez, S.M.; Borassi, C.; Barberini, M.L.; Aptekmann, A.A.; et al. Molecular link between auxin and ROS-mediated polar growth. Proc. Natl. Acad. Sci. USA 2017, 114, 5289–5294. [Google Scholar] [CrossRef] [PubMed]
- Tivendale, N.D.; Millar, A.H. How is auxin linked with cellular energy pathways to promote growth? New Phytol. 2022, 233, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Tognetti, V.B.; Mühlenbock, P.; Van Breusegem, F. Stress homeostasis—The redox and auxin perspective. Plant Cell Environ. 2012, 35, 321–333. [Google Scholar] [CrossRef]
- Meng, Q.; Zhang, C.; Gai, J.; Yu, D. Molecular cloning, sequence characterization and tissue-specific expression of six NAC-like genes in soybean (Glycine max (L.) Merr.). J. Plant Physiol. 2007, 164, 1002–1012. [Google Scholar] [CrossRef]
- Fan, K.; Wang, M.; Miao, Y.; Ni, M.; Bibi, N.; Yuan, S.; Li, F.; Wang, X. Molecular evolution and expansion analysis of the NAC transcription factor in Zea mays. PLoS ONE 2014, 9, e111837. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Maruyama, K.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Tran, L.S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J. Cell Mol. Biol. 2004, 39, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, Y.S.; Redillas, M.C.; Jang, G.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.H.; Reuzeau, C.; Kim, J.K. OsNAC5 overexpression enlarges root diameter in rice plants leading to enhanced drought tolerance and increased grain yield in the field. Plant Biotechnol. J. 2013, 11, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.H. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2000, 14, 3024–3036. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, S.; Wang, Y.; Zhang, X.; Lv, B.; Luo, L.; Xi, D.; Shen, J.; Ma, H.; Ming, F. OsNAC2 encoding a NAC transcription factor that affects plant height through mediating the gibberellic acid pathway in rice. Plant J. Cell Mol. Biol. 2015, 82, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.K.; Lindemose, S.; de Masi, F.; Reimer, J.J.; Nielsen, M.; Perera, V.; Workman, C.T.; Turck, F.; Grant, M.R.; Mundy, J.; et al. ATAF1 transcription factor directly regulates abscisic acid biosynthetic gene NCED3 in Arabidopsis thaliana. FEBS Open Bio 2013, 3, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yan, J.; Liu, W.; Liu, L.; Sheng, Y.; Sun, Y.; Li, Y.; Scheller, H.V.; Jiang, M.; Hou, X.; et al. Phosphorylation of a NAC Transcription Factor by a Calcium/Calmodulin-Dependent Protein Kinase Regulates Abscisic Acid-Induced Antioxidant Defense in Maize. Plant Physiol. 2016, 171, 1651–1664. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Seo, P.J.; Lee, H.J.; Park, C.M. A NAC transcription factor NTL4 promotes reactive oxygen species production during drought-induced leaf senescence in Arabidopsis. Plant J. Cell Mol. Biol. 2012, 70, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Liao, K.; Du, H.; Xu, Y.; Song, H.; Li, X.; Xiong, L. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2015, 66, 6803–6817. [Google Scholar] [CrossRef]
- Niu, L.; Hao, R.; Wu, X.; Wang, W. Maize mesocotyl: Role in response to stress and deep-sowing tolerance. Plant Breed 2020, 139, 466–473. [Google Scholar] [CrossRef]
- Mano, Y.; Nemoto, K. The pathway of auxin biosynthesis in plants. J. Exp. Bot. 2012, 63, 2853–2872. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Craig, J.C.; Petzold, H.E.; Dickerman, A.W.; Beers, E.P. The xylem and phloem transcriptomes from secondary tissues of the Arabidopsis root-hypocotyl. Plant Physiol. 2005, 138, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.-W.; Wang, J.-H. Effect of auxin on mesocotyl elongation of dark-grown maize under different seeding depths. Russ. J. Plant Physiol. 2010, 57, 79–86. [Google Scholar] [CrossRef]
- van Mourik, H.; van Dijk, A.D.J.; Stortenbeker, N.; Angenent, G.C.; Bemer, M. Divergent regulation of Arabidopsis SAUR genes: A focus on the SAUR10-clade. BMC Plant Biol. 2017, 17, 245. [Google Scholar] [CrossRef]
- Chae, K.; Isaacs, C.G.; Reeves, P.H.; Maloney, G.S.; Muday, G.K.; Nagpal, P.; Reed, J.W. Arabidopsis SMALL AUXIN UP RNA63 promotes hypocotyl and stamen filament elongation. Plant J. Cell Mol. Biol. 2012, 71, 684–697. [Google Scholar] [CrossRef]
- Xi, D.; Chen, X.; Wang, Y.; Zhong, R.; He, J.; Shen, J.; Ming, F. Arabidopsis ANAC092 regulates auxin-mediated root development by binding to the ARF8 and PIN4 promoters. J. Integr. Plant Biol. 2019, 61, 1015–1031. [Google Scholar] [CrossRef]
- Park, J.; Kim, Y.S.; Kim, S.G.; Jung, J.H.; Woo, J.C.; Park, C.M. Integration of auxin and salt signals by the NAC transcription factor NTM2 during seed germination in Arabidopsis. Plant Physiol. 2011, 156, 537–549. [Google Scholar] [CrossRef]
- Mao, C.; He, J.; Liu, L.; Deng, Q.; Yao, X.; Liu, C.; Qiao, Y.; Li, P.; Ming, F. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol. J. 2020, 18, 429–442. [Google Scholar] [CrossRef]
- Kan, C.; Zhang, Y.; Wang, H.L.; Shen, Y.; Xia, X.; Guo, H.; Li, Z. Corrigendum: Transcription factor NAC075 delays leaf senescence by deterring reactive oxygen species accumulation in Arabidopsis. Front. Plant Sci. 2021, 12, 691607. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, T.; Potters, G.; Caubergs, R.; Jansen, M.A. Complementary interactions between oxidative stress and auxins control plant growth responses at plant, organ, and cellular level. J. Exp. Bot. 2005, 56, 1991–2001. [Google Scholar] [CrossRef]
- Lu, X.; Liu, J.; Ren, W.; Yang, Q.; Chai, Z.; Chen, R.; Wang, L.; Zhao, J.; Lang, Z.; Wang, H.; et al. Gene-Indexed mutations in maize. Mol. Plant 2018, 11, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.Z.; Wang, B.G.; Gao, Q.H.; Zhang, Q.; Yan, G.Q.; Duan, K.; Huang, J.H. Molecular diversity and differential expression of starch-synthesis genes in developing kernels of three maize inbreds. Plant Cell Rep. 2009, 28, 1487–1495. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Annotation | B73_fpkm | zmnac17-1_fpkm | Log2FC |
|---|---|---|---|---|
| Zm00001eb396770 | Aldehyde oxidase AA01 (possible homologue gene in Arabidopsis) | 11.89 | 0.80 | −3.920 |
| Zm00001eb396780 | E3 ubiquitin-protein ligase UPL6 | 7.95 | 0.60 | −3.76 |
| Zm00001eb396760 | Alpha hydrolases-like domain-containing protein (possible homologue gene in Arabidopsis) | 3.43 | 0 | −7.34 |
| Zm00001eb416690 | Tryptophan decarboxylase 1-like | 0.11 | 0.64 | 2.45 |
| Zm00001eb396790 | Indole-3-acetaldehyde oxidase | 2.85 | 0.31 | −3.22 |
| Gene ID | Annotation | B73_fpkm | zmnac17-1_fpkm | Log2FC |
|---|---|---|---|---|
| Zm00001eb330530 | peroxidase 70 isoform X1 | 25.90 | 12.72 | −1.06 |
| Zm00001eb333290 | peroxidase 72 precursor | 18.69 | 9.29 | −1.04 |
| Zm00001eb111420 | peroxidase 66 precursor | 5.79 | 2.61 | −1.18 |
| Zm00001eb226370 | Peroxidase 45 | 2.22 | 4.78 | 1.07 |
| Zm00001eb109910 | peroxidase 2-like | 0.08 | 1.09 | 3.76 |
| Zm00001eb348950 | peroxidase 2 | 0.68 | 0.15 | −2.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Li, K.; Wang, M.; Sun, M.; Li, Q.; Chen, L.; Xiao, F.; Zhang, Z.; Zhang, H.; Jiao, F.; et al. ZmNAC17 Regulates Mesocotyl Elongation by Mediating Auxin and ROS Biosynthetic Pathways in Maize. Int. J. Mol. Sci. 2024, 25, 4585. https://doi.org/10.3390/ijms25094585
Yang R, Li K, Wang M, Sun M, Li Q, Chen L, Xiao F, Zhang Z, Zhang H, Jiao F, et al. ZmNAC17 Regulates Mesocotyl Elongation by Mediating Auxin and ROS Biosynthetic Pathways in Maize. International Journal of Molecular Sciences. 2024; 25(9):4585. https://doi.org/10.3390/ijms25094585
Chicago/Turabian StyleYang, Ran, Kangshi Li, Ming Wang, Meng Sun, Qiuhua Li, Liping Chen, Feng Xiao, Zhenlong Zhang, Haiyan Zhang, Fuchao Jiao, and et al. 2024. "ZmNAC17 Regulates Mesocotyl Elongation by Mediating Auxin and ROS Biosynthetic Pathways in Maize" International Journal of Molecular Sciences 25, no. 9: 4585. https://doi.org/10.3390/ijms25094585